
Abstract
Limitations of conventional regeneration modalities underscore the necessity of recapitulating development for periodontal tissue engineering. In this study, we proposed a novel three-dimensional pellet cultivation system for periodontal ligament stem cells (PDLSCs) to recreate the biological microenvironment similar to those of a regenerative milieu. Monodispersed human PDLSCs were cultured in medium with ascorbic acid and conditioned medium from developing apical tooth germ cells and were subsequently harvested from culture plate as a contiguous cell sheet with abundant extracellular matrix. The detached cell–matrix membrane spontaneously contracted to produce a single-cell pellet. The PDLSCs embedded within this cell–matrix complex exhibited several phenotypic characteristics of cementoblast lineages, as indicated by upregulated alkaline phosphatase activity, accelerated mineralization, and the expression of bone sialoprotein and osteocalcin genes. When this PDLSC pellets were transplanted into immunocompromised mice, a regular aligned cementum/PDL-like complex was formed. These results suggest that the combination of apical tooth germ cell–conditioned medium and endogenous extracellular matrix could maximally mimic the microenvironment of root/periodontal tissue development and enhance the reconstruction of physiological architecture of a cementum/PDL-like complex in a tissue-mimicking way; on the other hand, such PDLSC pellet may also be a promising alternative to promote periodontal defect repair for future clinical applications.
Introduction
As known, exogenous 3D biomaterial scaffolds, which appear to mimic the in vivo functions of extracellular matrix (ECM), play a crucial role in tissue engineering. Yet, because in available scaffolds there exist some shortages, including unforeseeable cell–biomaterial interactions, uneven degradation of the biomaterial, inflammatory reaction, and limited cell seeding efficiency, no scaffold has been widely used in periodontal tissue engineering so far. In recent years, the use of naturally occurring ECM materials as scaffolds for the repair and regeneration of tissues is receiving increased attention,9,10 and strategies using natural reaggregation potential to assemble monodispersed cells in a tissue-mimicking way represent a valuable extension of current scaffold-based tissue engineering initiatives. 11 Cell sheet engineering is an emerging scaffold-free method in the field of tissue engineering.12,13 Via either direct transplantation of cell sheets to host tissues or the homotypic layering of individual cell sheets, an entirely natural periodontal tissue assembled by cells, with mature ECM, therefore can be engineered, which avoids shortcomings of traditional scaffold-based design.14–16 Yet, there are some drawbacks in cell sheets. A principal disadvantage is their poor mechanical properties, which leads to the demand for additional materials such as fibrin gel during the stratification, and makes it hard to fabricate grafts with special thickness and controllable stiffness. 16
For periodontal tissue engineering, fabricating periodontal grafts is of great significance for the management of periodontal defects, and appropriate form and sufficient strength of periodontal grafts must be taken into consideration to allow placement of bioengineered material into the defect that prevents subsequent collapse of the overlying tissues into the defect site. 17 In addition, with increased attention for the potential therapeutic applications of periodontal ligament stem cells (PDLSCs) in periodontal regeneration, the optimal 3D environment that provides instructive and stimulatory signals necessary to regulate the stem cell self-renewal and differentiation also is an aspect deserved to be considered.18,19 It has been proposed that intercellular communications through growth factors and ECM are the basis for advancing differentiation of adult mesenchymal stem cells into periodontal cells (including cementoblast, fibroblast, and osteoblast).19,20 As a preliminary attempt to recapitulate events involved in early cementogenesis, our group has demonstrated that conditioned medium (CM) from developing apical tooth germ cells (APTGs) is able to provide the cementogenic microenvironment and induce differentiation of PDLSCs along the cementoblasts lineage. 21 Thus, the creation of 3D environment of ECM for PDLSCs and the intimate dynamic relationship between them will be the issue of key importance in present study.
Given the above, we proposed a novel 3D cell pellet cultivation system based on the cell sheet engineering, which aimed to fabricate periodontal grafts composed of aggregated PDLSCs and self-produced ECM in a tissue-mimicking way, and presented a feasible strategy to construct 3D structures of periodontal tissues by using these cell pellets. We hypothesized that the novel PDLSC pellets could provide grafts with better handling properties than cell sheets, which may facilitate the operation of repairing the periodontal defects caused by periodontitis in the future; on the other hand, the combination of APTG-CM and endogenous ECM could maximally mimic the microenvironment of root/periodontal tissue development and enhance the reconstruction of the physiological architecture of a PDL–cementum complex.
Materials and Methods
PDLSC isolation and establishment of co-culture system
Human PDLSCs were isolated as previously described21,22 and cultured with α-minimum essential medium (α-MEM; Gibco BRL, Gaithersburg, MD) supplemented with 10% fetal bovine serum (FBS). The stem cell nature of isolated putative PDLSCs was confirmed by fluorescence-activated cell sorting analysis for STRO-1 and CD146 (R&D Systems, Minneapolis, MN) according to the manufacturer's protocol. Conditions for the induction of calcium accumulation were as reported previously. 23 Calcium accumulation was detected by 2% Alizarin Red S (pH 4.2) staining. The induction of adipogenesis was as previously reported. 24
Eight-day postnatal Sprague-Dawley rats were killed by decapitation under anesthesia. Developing mandibular first molar germs were dissected from jaws. The apical portions of tooth germs were separated as we have previously reported. 21 Single-cell suspensions were then generated by filtration through a 70 μm strainer, placed into 75 cm2 culture flasks (Costar, Cambridge, MA) at 1 × 105 cells/mL, and cultured in α-MEM containing 10% FBS. The culture medium of primary APTGs was changed every 48 h until full confluence for collecting the supernatants, which would be filtered through a 0.22 μm Millipore strainer (Carrigtwohill, Cork, Ireland). The supernatants were mixed with equal volume of fresh α-MEM supplemented with 10% FBS and 50 μg/mL ascorbic acid (Sigma, St. Louis, MO) and used as apical tooth germ cell–conditioned medium (APTG-CM) for PDLSCs co-culture. The α-MEM supplemented with 10% FBS and 50 μg/mL ascorbic acid was used as a control medium.
Culture strategy for generation of PDLSC pellets
Multiple colony-derived human PDLSCs from third passage were seeded at initial density of about 2.5 × 104/cm2 into culture plates and incubated for 48 h in normal α-MEM culture medium containing 10% FBS. Then, cell culture medium was changed to APTG-CM supplemented with 50 μg/mL ascorbic acid and replaced every 3 to 4 days for further 14 days. During the monolayer cultivation, the PDLSCs secrete white substances on the bottom of the dish, and a PDLSC–matrix membrane formed and can be easily detached from the bottom of the culture plate with cell scraper in 0.25% trypsin/EDTA solution (Fig. 1, Step 1). Finally, the wavy cell sheet was transferred to a 10-mL conical polypropylene tube (Asahi Techno Glass, Tokyo, Japan) and incubated en masse in the presence of 2 mL APTG-CM plus 50 μg/mL ascorbic acid. Within 12–24 h of incubation, the cell sheet began to shrink and formed a cell mass that did not adhere to the walls of the tube (Fig. 1, Step 2). Changes of the medium were carried out at 24 h intervals, and PDLSC pellets were harvested at time points up to 7 days. For the examination of the microscopic structure, the pellets were fixed in 10% neutral buffered formalin, dehydrated in a series of ethanol, embedded in paraffin, and cut into 5 μm sections. After deparaffinization, the sections were evaluated via either hematoxylin and eosin (H&E) or Masson trichrome staining.
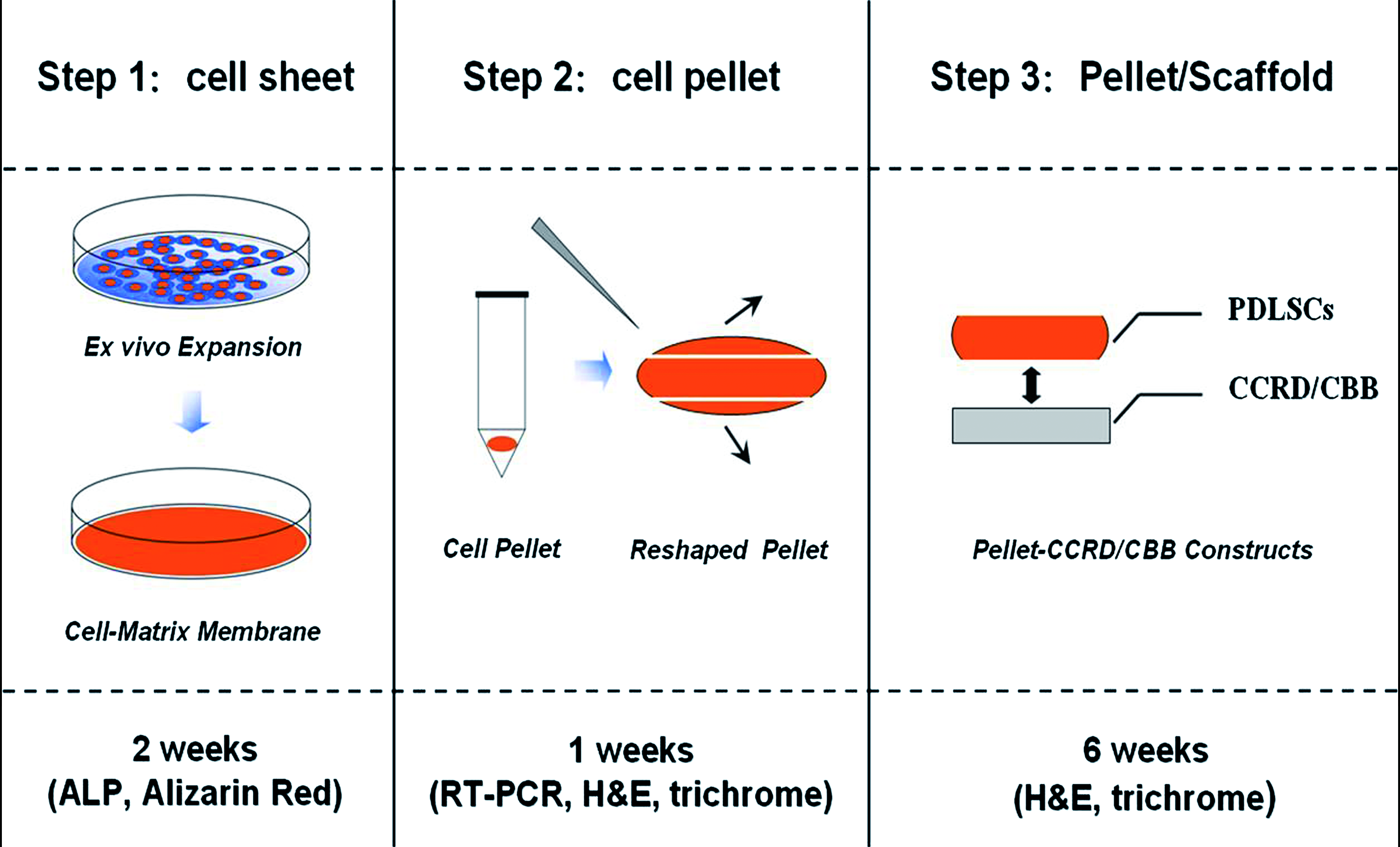
Schemes of construction strategy for generation of a cementum/PDL complex using PDLSC pellets. Step 1: Isolated PDLSCs were seeded on the culture dish and expanded in monolayer culture to form a cell–matrix membrane. Step 2: PDLSC sheet in a conical polypropylene tube condensed into an essentially spherical pellet that did not adhere to the walls of the tube; at time points up to 7 days, PDLSC pellet was harvested and reshaped. Step 3: Reshaped PDLSC pellets were placed onto the surface of CBB/CCRD and further fabricated into PDLSC pellet–CBB/CCRD grafts. Color images available online at
Alkaline phosphatase activity and Alizarin red staining
ALP activity assay was performed to determine the influence of APTG-CM on ALP expression. After 3, 7, and 14 days of co-culture with APTG-CM, ALP activity of PDLSC sheets was detected using an ALP assay kit (Zhongsheng, Beijing, China). The results were measured at 405 nm in a spectrophotometer using a microplate reader. Meanwhile, cytochemical ALP staining was also carried out. At time points up to 14 days, confluent monolayer cell sheets of PDLSCs in six-well culture dishes were thoroughly washed with 1 × phosphate-buffered saline (PBS), fixed with 2% paraformaldehyde dissolved in PBS, and stained cytochemically using the azo-dye coupling technique. 25
In addition, to investigate nodule formation and mineralization, the samples of PDLSC sheets initially cultured in APTG-CM, containing 10% FBS and 50 μg/mL of ascorbic acid, for 14 days were fixed with 4% polyoxymethylene for 0.5 h. Alizarin red staining was performed to reveal calcium deposition as previously reported. 23 The mineralized nodules were imaged and analyzed with Leica Q-Win image analysis software.
Reverse transcriptase polymerase chain reaction
After 7 days of incubation in APTG-CM, PDLSC pellets were harvested. Total cellular RNA was isolated by Trizol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's recommendations. Isolated RNA was then used as a template for cDNA synthesis, prepared using a Superscript II first-strand cDNA synthesis kit (Invitrogen Life Technologies, Carlsbad, CA). The expression of various transcripts was assessed by polymerase chain reaction (PCR) amplification using a standard protocol. Primer sets used in this study were as listed below:
Osteocalcin (OCN)—sense, 5′-atgagagccctcagactcctc-3′; antisense, 5′-cgggccgtagaagcgccgata-3′ Bone sialoprotein (BSP)—sense, 5′-gaaccacttccccacctttt-3′; antisense, 5′-tctgaccatcatagccatcg-3′ Collagen type I (COLI)—sense, 5′-ctgaccttcctgcgcctgatgtcc-3′; antisense, 5′-gtctggggcaccaacgtccaaggg-3′ Glyceraldehyde-3-phosphate (GAPDH)—sense, 5′-tccaccaccctgttgctgta-3′; antisense, 5′-accacagtccatgccatcac-3′ for the internal quantitative control.
The amplifications process was performed for 30 cycles after an initial 30 s denaturation at 94°C, annealed for 30 s at 56°C, and extended for 1 min at 72°C. The PCR products were separated, and observed on 1.5% agarose gel containing 5 g/L ethidium bromide.
Preparation of ceramic bovine bone and chemical conditioned root dentin
The blocks of ceramic bovine bone (CBB) were produced from fresh bovine rib bones. 26 The resected ribs were boiled, cleaned from the periosteum and adjacent connective tissues, and cut into blocks. Then, the blocks were washed in normal saline and soaked in H2O2 to remove proteins. After washing in running water, the deproteinized blocks were heated at 900°C for 1 h, cut into round, thin slices (thickness 1.0 mm, diameter 3.0 mm), and thereafter cleaned and sterilized with ethylene oxide for spare use.
Chemical conditioned root dentin (CCRD) specimens were prepared from collected human teeth after the periodontal tissues were removed for obtaining the PDLSCs. The CCRD was also cut into discs (1.0 mm thick) with a diameter approximating 3.0 mm. A series of chemical procedures were performed according to reported protocols with minor modifications. 27 In brief, dentin discs were treated with 17% EDTA for 10 min and 0.6 M HCl for 1–10 days to achieve complete demineralization, and with betadine and 5.25% NaOCl for 30 min each for sterilization, and then rinsed and soaked with 1 × PBS for 2 days and culture medium for 5 days to remove the residual sterilization agents.
In vivo differentiation of PDLSCs pellets
For the ectopic transplantation, PDLSC pellets co-cultured with APTG-CM (as experimental group) for 3 weeks were reshaped (approximately 3 × 3 mm), placed onto the surface of CBB/CCRD in culture dishes, and incubated in a small volume of culture medium (1 mL of α-MEM) at 37°C for 2 h to allow stable adhesion (Fig. 1, Step 3). All animal procedures complied with the guidelines provided by the Animal Care Committee of Fourth Military Medical University. Six 6-week-old male mice (BALB/c-nu; FMMU Medical Laboratory Animal Center, Xi’an, China) with severe combined immunodeficiency were used as hosts for the allogenic transplantation. General anesthesia was administered by intraperitoneal injection of 2% sodium pentobarbital sodium (40 mg/kg) for all surgical procedures. The host mice underwent aseptic preparation on their back, then midsagittal incisions were made on the dorsa, and the constructs of PDLSC pellets with CBB/CCRD were implanted into subcutaneous pockets on the left side. As a control, the constructs of untreated PDLSC pellets with CBB/CCRD were implanted into the other side of the same host. A total of 36 grafts were implanted. Each immunodeficient mouse received a set of six different grafts: a CBB/CCRD without cells and two types of PDLSC pellet–CBB/CCRD complexes (i.e., one that had, and one that had not, been cultured in the APTG-CM). The implants were recovered at 6 weeks posttransplantation, fixed with 4% paraformaldehyde for 2 days, and then decalcified for a further 14 days in 10% EDTA (pH 8.0) before embedding in paraffin. For histological analysis, 5 μm sections of implants were prepared and stained with either H&E or Masson trichrome staining.
Statistical analyses
Student's t-test was used to analyze the significance between the two groups with SPSS software (version 12.0; SPSS, Chicago, IL). p-Values of less than 0.05 were judged to be statistically significant.
Results
Isolation and characterization of PDLSCs
In the present study, we used the limiting dilution technique to purify human PDLSCs based on their ability to form adherent clonogenic cell clusters of fibroblast-like cells, which was similar to those observed for different mesenchymal stem cell populations. Single-cell suspensions were prepared after trypsin/EDTA digestion of subconfluent primary cultures of PDL cells and then subjected to clone selection. The isolated PDLSCs were capable of forming adherent colonies, characteristic of other stromal stem cell populations, and majority of the cells retained their fibroblastic spindle shape (Fig. 2A). In addition, fluorescence-activated cell sorting analysis was performed to characterize the phenotype of ex vivo–expanded PDLSCs, and approximately 12.8% of the PDLSCs stained positive for STRO-1 and 78.5% for CD146 (Fig. 2D, E).

Characterization of human PDLSCs. (
To investigate the differentiation potential of PDLSCs, multiple colony-derived PDLSCs at passage six were grown in the presence of 50 μg/mL L-ascorbate-2-phosphate, 10 nM dexamethasone, and 10 mM inorganic phosphate to induce mineralization in vitro as described previously. After 4 weeks of induction, cultured PDLSCs formed extensive amounts of alizarin red-positive mineral deposits throughout the adherent layers (Fig. 2B). Moreover, PDLSCs were capable of undergoing an adipogenic differentiation with an adipogenic-inductive cocktail (Fig. 2C).
In vitro examination of PDLSC pellets
When PDLSCs were seeded into culture plates in α-MEM plus 50 μg/mL ascorbic acid, they showed significant accumulation of abundant ECM and rapidly proliferated to a confluent state within 24–48 h (Fig. 3B). After 14 days 2D monolayer culture, a semitransparent membrane of cell–matrix complex formed, which was stiff enough to be intactly detached from the edge of culture dish by scraping technique after trypsin/EDTA treatment (Fig. 3E, F). By contrast, cells grown in culture medium without ascorbic acid seldom formed cell–matrix structures and separated into many small cell masses immediately after treatment with trypsin/EDTA solution (Fig. 3C). Therefore, α-MEM containing 50 μg/mL ascorbic acid was ideal for collagen synthesis and construction of a cell–matrix membrane. When transferred to a 10-mL conical polypropylene tube, the cell sheet began to shrink and condensed into an aggregate immediately within 12–24 h and finally formed an essentially spherical pellet (Fig. 3G). Usually, PDLSC pellets steadily maintained their sizes after 7 days in culture. Pellet diameters mainly depended on original cell sheet areas, which were about 1.5 and 3.5 mm for different original cell sheet areas of 9.6 (areas of each well of six-well culture plate) and 49 cm2 (areas of 90 mm culture plate), respectively, after 7 days of incubation in the tube.

PDLSCs were cultured for 14 days in culture medium with (
As expected, when PDLSCs were cultured in α-MEM supplemented with APTG-CM for 2–3 weeks, they exhibited several characteristics of osteo/cementoblast phenotype, as indicated by markedly increased ALP activity and calcium deposition as well as high expression of BSP and OCN transcripts (Fig. 4), which has been demonstrated to be selectively expressed by root-lining cells. 28 Histological assessment of PDLSC pellets revealed a uniform tissue structure with a connective tissue–like morphology, which contained cells entrapped within an ECM (Fig. 5). Although the ECM was abundant, collagen fibrils were arranged randomly into the matrices, and the cells embedded in the matrices were also not oriented in any particular manner. There is no obvious difference in histological appearance between PDLSC pellets co-cultured with or without APTG-CM (data not show). Nevertheless, with extended cultivation time of 10 days in the tube, significant histological changes (a lot of fractures appeared within PDLSC pellets) could be observed, which is generally believed to be the signs of inadequate supply of nutrients (Fig. 5I–L). It seemed that these changes appeared in a time- and diameter-dependent manner.

(

Histological assessment of PDLSC pellets stained with either H&E or Masson trichrome staining. (
In vivo evaluation of PDLSC pellet–CBB/CCRD constructs
A total of 36 specimens, including surrounding soft tissues, in current study were harvested and evaluated by light microscopy (DX51; Olympus, Tokyo, Japan) after HE and Masson trichrome staining. In experimental group, a typical regularly arranged cementum/periodontal-ligament structure was regenerated on the CBB/CCRD surfaces in all (six of six, 100%) specimens of PDLSC pellet–CBB constructs and four of six (66.7%) specimens of PDLSC pellet–CCRD constructs. Histological observations revealed cementum-like mineralized tissues lining the CBB/CCRD surfaces and PDL-like fibrous tissues interfaced with these newly deposited cementum-like tissues (Fig. 6A, E). Significantly, higher magnification revealed that large collagen bundles were inserted perpendicularly into cementum-like tissues, and this orientation closely resembles the physiological attachment of natural Sharpey's fibers in the PDL (Fig. 6B, F). Masson trichrome staining further confirmed the collagen content and the orientation of PDL-like tissues (Fig. 6I, K). By contrast, in control group transplants of CBB/CCRD with PDLSC pellets cultured without APTG-CM seldom formed cementum/PDL-like tissues throughout the observation periods. Only two of six (33.3%) specimens of PDLSC pellet–CBB constructs and one of six (16.7%) specimens of PDLSC pellet–CCRD constructs formed small amounts of cementum-like deposits. Regularly aligned connective tissues oriented perpendicularly to the CBB surfaces were found in samples of PDLSC pellet–CBB constructs (Fig. 6C). As to the constructs of untreated PDLSC pellets integrated with CCRD, cells and/or surrounding soft tissues were detached from the dentin surfaces during histological sectioning in most specimens, implying weak integration between dentin surfaces and PDLSC pellets (Fig. 6G). In the negative control where individual CBB/CCRD without cells was transplanted, no tissue regeneration was observed (Fig. 6D, H).
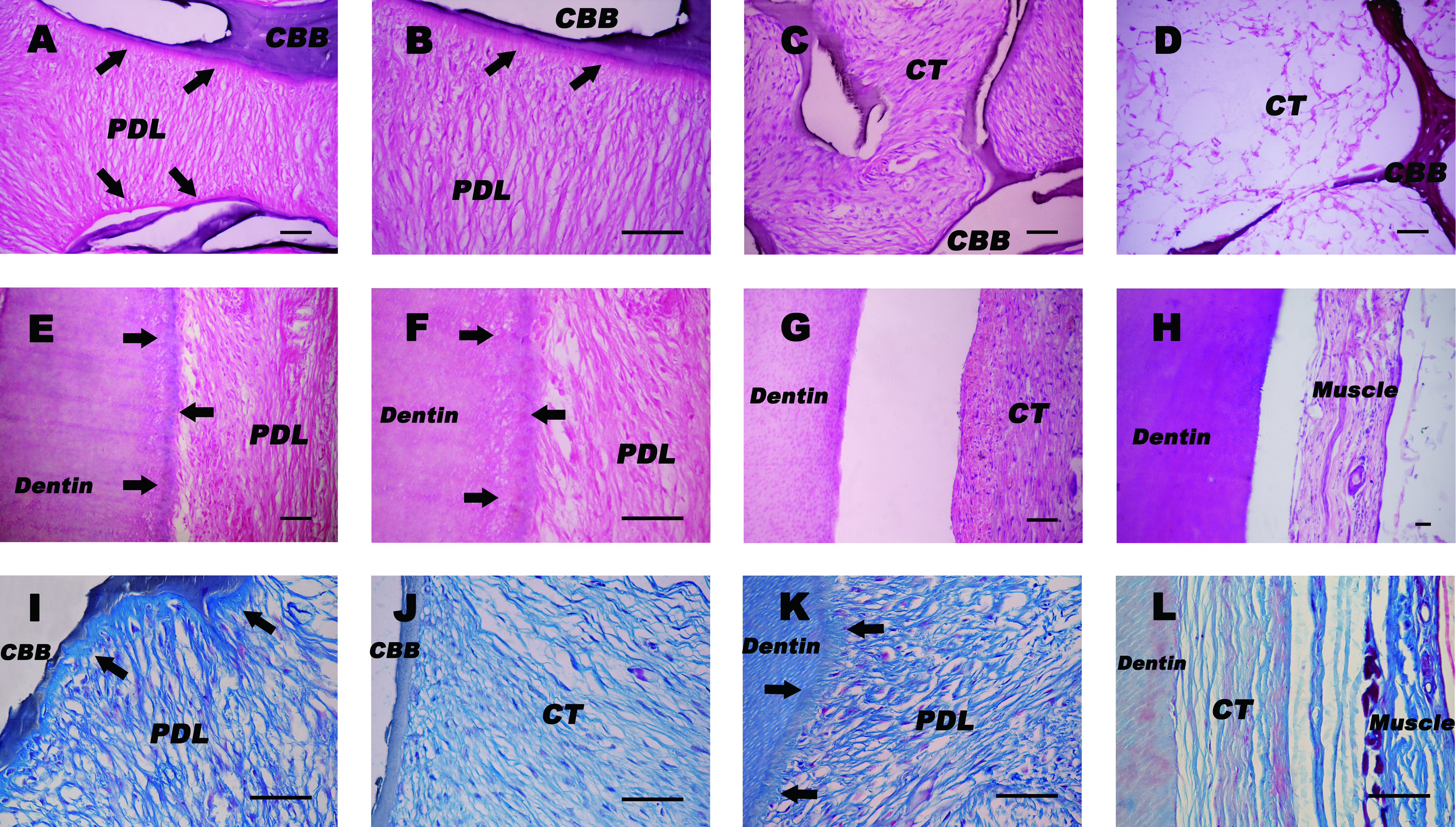
Histological examination 6 weeks after subcutaneous transplantation of PDLSC pellet–CBB/CCRD constructs. (
Discussion
PDLC-based therapies have been extensively studied since the successful report that cultured PDL cells applied in various scaffolds possess the capability to induce a new periodontal tissue apparatus on root dentin surfaces and dental implants.3–6 However, these procedures may have various limitations and thus underscores the requirement that a scaffold-free cell delivery system be introduced into the reconstruction of periodontal tissues. The present study demonstrated the feasibility of using a novel 3D cell pellet cultivation system for human PDLSCs to obtain the complete reconstruction of the physiological architecture of a cementum/PDL complex, and the following important points have been yielded: (1) a novel scaffold-free PDLSC pellet can be generated using an easy and efficient culturing protocol; (2) such PDLSC pellet generated in the presence of ascorbic acid presents some important biological characteristics as compared with conventional tissue engineering approaches; (3) this novel pellet cultivation strategy is a good in vitro experimental model of recapitulating the events involved in the original formation of the periodontal tissues; (4) the physiological architecture of a cementum/PDL complex was reconstructed in an immunocompromised mouse model with a tissue-mimicking way.
Development of appropriate scaffold-free periodontal grafts is one of the main targets of current study. Compared with cell sheet engineering and traditional pellet culture system generally used for chondrogenesis,29,30 there are some distinct features of present pellet culture strategy. First, this method does not require any specific substrate or temperature-responsive dishes and thus could be an easier and more practical method to develop a construct without an artificial scaffold. Cell pellets can be generated from most types of cell species that respond to ascorbic acid for collagen synthesis. Second, sufficient ECM has been formed before the creation of the pellet in present method, while the initial amount of ECM is essentially zero in traditional chondrogenic pellet culture system, which means much cost of time for pellet culture (over 4 weeks) for production of ECM and close cell–cell adhesion. The PDLSC pellets we described here exhibited some important biological characteristics, including high cell utility efficiency, even cell distribution, easy and time-saving fabricating procedure, and generation of periodontal grafts with satisfactory size and good handling properties. Although the present experiments used a xenogeneic model utilizing cultured human PDLSCs in immunodeficient mice, it was not difficult to envisage that PDLSC pellets that composed of autogenic cells, and self-secreted ECM will be less immunogenic and much more biocompatible and provide a promising alternative for current approaches to repair periodontal defects in the future.
There is another unique biological characteristic of the PDLSC pellet. Over the decade, a number of in vitro studies have shown that 3D environment of ECM and growth factors regulate the differentiation of stem cells. 31 It has been found that a feeder layer of fibroblasts is not required if a basement membrane ECM, Matrigel™, and fibroblast conditioned medium are used to culture human embryonic stem cells. 32 When bone marrow–derived adult stem cells are cultured on basement membrane Matrigel with fibroblast growth factor 4 (FGF-4) and hepatocyte growth factor (HGF), they have morphological, functional, and phenotypical characteristics of hepatocytes. 33 Classical recombination experiments indicate that tooth development actually derives from the reciprocal and sequential inductions of different cell pellet termed “tooth buds”—that is, the interactions of epithelial and mesenchymal cells.34,35 As an integrated part of tooth structure, there is no exception in root/periodontal tissue development. Because cellular self-organization is an intrinsic property of the tissues undergoing development, 36 using the ability of monodispersed cells to self-assemble into a microtissue such as a 3D spheroid,37,38 we proposed this 3D scaffold-free cell pellet cultivation system. Within this system the cells contact only with one another and with the ECM produced by them, which partially imitates the natural environment and complies with the notion that the pellet (or spheroid) is a good in vitro experimental model reflecting in vivo status of cells in living tissues or organs.39,40 In addition, this self-assembly approach for tissue reconstruction is more akin to the physiological phenomenon that occurs in the womb during organogenesis.41,42 By use of endogenous scaffolds formed by natural ECM and developing periodontal microenvironment provided by APTG-CM, this in vitro pellet cellular system undergoes a process that resembles many aspects of tissue development, including cell proliferation, aggregation, and condensation, the expression of tissue-specific genes, and most importantly, morphogenesis and pattern formation of periodontal tissues, so it is actually a simplified model of recreating early periodontal tissue development.
What is the most worth mentioning in this study is that the physiological architecture of cementum/PDL complex was successfully reconstructed in a xenogeneic subcutaneous model. As known, during the process of embryonic development and tissue formation, epithelial–mesenchymal interactions through growth factors and ECM are necessary for cell differentiation, tissue regeneration, and morphogenesis. 43 Previous investigations44–46 on odontogenesis have revealed that although monodispersed dental papilla mesenchymal cells lost original intercellular ECM during cell preparation process, reaggregated mesenchymal cells in pellet culture could secrete new ECM and growth factors necessary for tissue formation and regeneration. The presence of ECM, with its endogenous growth factors, might mimic the odontogenic microenvironment created by embryonic basement membrane and inductive signals, which was responsible for the recurrence of dentinogenesis and morphogenesis. MacNeil and Thomas,47,48 in a series of tissue recombination experiments, showed clearly that cells from dental follicle tissue could produce an adherent cementum-like mineralized tissue on the surface of dentin fragments, where root-associated basement membrane (RBM) was preserved. Further, when epithelial root sheath (ERS) was included in recombinations between root dentin specimens and dental sac, cementum-like tissue also formed in the absence of RBM; meanwhile, part of recombinations in which RBM was preserved formed PDL-like tissues. This suggests that ERS and basement membrane play important roles in cementogenesis and PDL formation. As far as current study is concerned, the periodontal microenvironment from developing APTGs served the similar role of ERS, and PDLSCs co-cultured with APTG-CM present some features of cementoblast lineages, as indicated by upregulated ALP activity, accelerated mineralization, and the expression of BSP and OCN genes along with the in vivo cementum-like tissue formation. On the other hand, natural ECM produced by PDLSC pellet acted as role of basement membrane and induced the recurrence of regular aligned cementum/PDL-like complex, which further confirmed the biologically active role of ECM in tissue morphogenesis. 49
The ultimate goal of periodontal therapy is the regeneration of attachment apparatus lost due to periodontitis with the formation of new bone, new cementum, and a functionally oriented PDL. To create a microenvironment favorable for selective cell repopulation and subsequent matrix synthesis, dentin root surfaces have been conditioned with various agents. 50 Prior study has demonstrated that fibronectin is a major ECM protein that is synthesized and secreted by cultured PDL cells.51,52 So, fibronectin matrix incorporated into the ECM could function as a natural adhesive to attach PDLSC pellets to root surfaces as it did in HPDL cell sheets previously reported.14,16,53 Further, simple and rapid implantation procedures based PDLSC pellets for the repair of periodontal defects in the future could be expected to proceed without additional augmentation of the initial fixation. Besides, the periodontium is such unique environment that it consists of two mineralized tissues in proximity: bone and cementum, as well as a nonmineralized interface—the PDL. Although it has been reported that matrix elasticity could direct mesenchymal stem cell lineage specification, 54 no definitive data in current study are yet available to show that natural dentin matrix and bone matrix may be effective for PDLSCs differentiation into cementoblasts and osteoblasts, respectively. Therefore, it is not too difficult to envisage the engineering of multilayered cell pellets comprising calcified bone-forming cell pellets (i.e., bone marrow mesenchymal stem cells) and cementum/PDL-forming cell pellets (i.e., PDLSCs) to fully reconstruct various compartments of the periodontium in vitro and then implant such constructs into critical-sized periodontal defects.
To sum up, in the present work we proposed a novel 3D cell pellet cultivation system for human PDLSCs to obtain the complete reconstruction of the physiological architecture of a cementum/PDL complex. Further orthotopic transplantation experiment of surgically created acute periodontal defects in animals may lead to the application of this tissue-engineered construct for periodontal regeneration in clinical settings.
Footnotes
Acknowledgments
This work was supported by grants from the Nature Science Foundation of China (Project Nos. 30572046 and 30725042).
Disclosure Statement
No competing financial interests exist.