
Abstract
While infected skin wounds are on the increase because of ageing populations, rising incidence of diabetes, and antibiotic resistance, we lack relevant in vivo or in vitro models to study many aspects of bacterial interaction with skin. The aim of this study was to develop three-dimensional models of normal human skin to study bacterial infection. The common dermatological pathogens Staphylococcus aureus and Pseudomonas aeruginosa were used to infect tissue-engineered skin, and the course of infection in the skin was examined over several days. Two forms of model were developed—one in which bacteria were introduced directly to 10 mm wounds in the epidermis, and another in which wounds were created by burning a 4 mm hole in the center of the tissue before inoculation. The bacteria flourished within the engineered skin, and colonized the upper epidermal layers before invasion into the dermis. Infection with P. aeruginosa caused a loss of epidermis and de-keratinization of the skin constructs, as well as partial loss of basement membrane. These novel complex human skin infection models could be used to investigate microbial invasion of normal skin epithelium, basement membrane, and connective tissue, and as a model to study approaches to reduce bacterial burden in skin wounds.
Introduction
The development of reconstituted human tissue models consisting of multiple cell types and naturally formed extracellular matrix for studies aimed at mimicking human in vivo disease processes has many advantages over 2D cell cultures and even on occasional animal in vivo models. Advances in tissue engineering over the last 20 years have led to the development of tissue-engineered skin models that closely resemble normal human skin.18–21 These models have been used to study the mechanisms of various cutaneous conditions, including skin graft contraction,22,23 psoriasis, 24 melanoma,25,26 chemical irritation, 27 and skin pigmentation, 28 but have not so far been used to study bacterial infection. A previously published model, using a skin equivalent of dermal matrix composed of fibrin populated with fibroblasts rather than human decellularized dermal scaffold (DED), has been successfully used to investigate skin colonization by cutaneous flora. 29 However, the skin equivalent constructs used in the study possessed intact epidermides and therefore do not represent skin injury and the resulting invasion and infection by opportunistic pathogens or commensal flora.
Excised pieces of human skin have been used to study fungal skin and HIV infections with some success.30–32 However, these models are difficult to standardize because the skin is directly excised from patients, the skin must be used within hours of excision, and reproducibility is not guaranteed. In contrast, the reconstituted skin model employed in our laboratory is robust and extremely reproducible. The tissue-engineered skin shares many of the properties of normal skin such as a well-differentiated keratinocyte layer, a convoluted epidermal–dermal junction retaining a basement membrane, and a fibroblast populated dermis, and is histologically similar to human skin. It has been used clinically to treat skin contractures, 33 and an oral mucosa version (based on the same methodology) has been used to treat scarring of the urethra;34,35 these studies attest to the physiological relevance of the tissue-engineered skin and oral mucosa.
The purpose of the current study was to develop a physiologically relevant model of skin infection that could be used to study any cutaneous invasive bacterial or fungal infection. We used two species of bacteria well known for their ability to invade skin: Pseudomonas aeruginosa, a Gram-negative opportunistic pathogen that particularly infects wounded skin such as that of burns victims, 36 and Staphylococcus aureus, a Gram-positive organism responsible for skin infections ranging from abscesses to impetigo. 37 These two bacteria, although not unique in their capacities to cause dermatological infections, 38 account for millions of clinical cases each year worldwide and contribute to thousands of fatalities, both community-acquired and hospital-acquired.39–43
Reconstructed human skin models were produced by seeding normal human skin, laboratory–expanded keratinocytes and fibroblasts onto a DED that was treated to retain its native basement membrane. After 10–14 days culture at an air–liquid interface, the reconstituted tissue develops characteristics very similar to normal skin. In this study, to create a higher-throughput system, we also made skin constructs grown within 10-mm-diameter tissue culture inserts rather than the previously used 2.5-cm-diameter models raised on grids. The use of inserts allowed us to create smaller skin constructs, thus maximizing the use of the tissue available.
We also explored two methods of introducing the two bacterial species into the skin tissue: (1) seeding bacteria into an area of DED lacking epidermis and (2) by burning the skin constructs before introducing bacteria. Both methods of introducing bacteria were effective; however, the smaller skin constructs are potentially more useful as they require less skin to establish. The burnt skin is also particularly suited to the study of P. aeruginosa and S. aureus infection; both species commonly infect burn wounds, and the rise of antibiotic resistant strains of S. aureus along with the inherent and acquired antimicrobial resistance factors of P. aeruginosa make treatment of infected burn wounds difficult. The model described in the current study could provide useful data for generating novel antimicrobial/decontamination strategies for burns victims.
Materials and Methods
Skin constructs
Skin constructs were produced as previously described. 19 Briefly, human dermal fibroblasts and keratinocytes were isolated from split-thickness skin biopsies received from abdominoplasties and breast reductions performed at the Northern General Hospital, Sheffield. Sterile decellularized dermis (DED), also sourced from human skin biopsies, was used as a base scaffold. DED varied in thickness due to harvesting methods but was generally ∼0.5 mm deep. In the case of the larger skin constructs, DED was placed in tissue culture wells with the reticular surface uppermost. Sterile 27-mm-diameter steel rings were placed on each piece of dermis, and a seal was obtained by applying gentle pressure. A second, smaller sterile steel ring (0.8 cm2) was then placed in the middle of the larger ring. Dulbecco's modified Eagle's medium (10% fetal calf serum, penicillin and streptomycin/fungizone was used to flood the dermis surrounding the larger ring, and the dermis within the center of the smaller ring. The ring of bare DED between the two rings was then seeded with 1 × 105 fibroblasts and 5 × 105 keratinocytes, each in 250 μL of Greens' medium supplemented with 10% fetal calf serum. After 24 h incubation at 37°C, seeding medium was removed and replaced with fresh Greens' medium. After a further 24 h, both rings were removed and the skin constructs raised onto stainless steel grids in new tissue culture wells. Fresh 10% Greens' was added to the level of the base of the skin constructs, forming an air–liquid interface. Medium was changed every 2–3 days, and skin constructs were used for experimentation after 10 days at the air–liquid interface.
For the smaller skin constructs, rings of DED 15 mm in diameter were cut using a sterile cork borer and placed within 12 mm tissue culture inserts with 4 μm pores in the base (Greiner). DED was cut slightly larger than the inserts to allow for skin contraction on burning. Inserts were suspended from the edges of 12-well plates into the wells. The 10% Greens' medium was added to the bottom of the wells so that it lapped the under surface of the DED. The DED was then seeded with 1 × 105 fibroblasts and 5 × 105 keratinocytes, each in 250 μL of 10% Greens' medium. After 24 h at 37°C, the seeding medium was replaced with fresh Greens' medium, which was removed from inside the inserts 24 h later to allow the skin constructs to be at the air–liquid interface. Subsequently Greens' medium was replaced every 24 h and skin constructs were used for experimentation after 10–14 days at the air–liquid interface.
Bacteria
Clinical isolates of P. aeruginosa (SOM1) and S. aureus (S-235) and the laboratory strain S. aureus NCTC 6571 were cultured in 20 mL Brain Heart Infusion (BHI; Oxoid, Basingstoke, UK) broth at 37°C for 24 h from stock plates before use. Broths were centrifuged, and the resulting pellet was washed and resuspended in PBS to a concentration of 1 × 1010 cfu/mL.
Skin construct infection
Skin constructs were washed in antibiotic-free Greens' medium for 72 h before infection. Larger constructs were infected by pipetting 1 × 107 bacteria in 20 μL BHI broth per composite directly onto the exposed dermis in the center of the skin constructs. Smaller constructs in inserts were burnt by the application of a heated metal rod (4 mm diameter), for 6 s immediately before infection to provide the bacteria a mode of entry into the skin. 1 × 107 bacteria in 100 μL BHI broth per construct were pipetted into the inserts covering the epidermal surface. Infected skin constructs and noninfected controls were incubated in antibiotic-free Greens' medium at 37°C/5% CO2 for 24 or 72 h postinfection, and then sacrificed for analysis. Skin constructs were bisected with sterile scalpels. Half of the construct was fixed in 10% formalin for >24 h, and then processed and paraffin embedded for histological analysis. The other half was weighed, and then the tissue was homogenized in 1 mL BHI. The resulting homogenates were diluted serially and used to perform viable counts of bacteria in the samples.
Histology
Formalin-fixed, paraffin-embedded samples were sectioned to 6 μm thick on a microtome. Sections were stained with hematoxylin and eosin using standard techniques, and also Gram stained to visualize bacteria, again using standard techniques. Numbers of P. aeruginosa visible in 150 ×100 μm sections were quantified and expressed as number of bacteria/field of view at a magnification of 60 ×.
Immunohistochemistry
Sections were fluorescently stained for keratinocyte and basement membrane antigens using antibodies against pancytokeratin (mouse monoclonal anti-pancytokeratin PCK-26; Abcam, Cambridge, UK) and type IV collagen (rabbit polyclonal anti-collagen IV; Abcam), respectively. High-temperature antigen retrieval was performed on dewaxed sections in sodium citrate buffer, pH 6.0, before addition of primary antibodies at 1:100 in PBS, which were incubated at 4°C overnight. Secondary antibodies (FITC-labeled polyclonal sheep anti-mouse IgG [Abcam] and Cy3-labeled polyclonal goat anti-rabbit IgG [Abcam]) at 1:200 were incubated for 30 min at room temperature, and following washing steps in PBS sections were mounted using a DAPI-containing mountant (Vectashield; Vector Laboratories, Burlingame, CA). Images were captured and processed on a Zeiss Axiovert 200 M inverted fluorescence microscope, using Axiovision Rel 4.6 software (Zeiss, Welwyn Garden City, UK).
Statistics
Statistical analyses were performed using the paired t-test.
Results
Morphology of skin constructs
Histological examination of the epidermal layers of the larger dermis-exposed skin constructs reveals them to be structurally similar to human skin (Fig. 1b). The epidermal layers display well-differentiated keratinocytes, convoluted epidermal–dermal junctions retaining basement membranes, and fibroblast-populated dermis (Fig. 1b). In the larger skin constructs, the exposed dermal area is clearly demarked from the full thickness skin (Fig. 1c). Infection of the larger skin constructs with either 1 × 107 cfu S. aureus (Fig. 1e) or P. aeruginosa (Fig. 1f) resulted in colonization of the epidermal layers and, in the case of P. aeruginosa, invasion into the dermis after 24 h infection.

Creation of skin constructs with breaks in the epidermal barrier. (
Skin constructs grown within tissue culture inserts are also histologically similar to normal human skin (Fig. 2b). Burn injury to the smaller skin constructs results in a dysfunctional epidermal layer surrounded by unaffected, healthy epidermis (Fig. 2c).
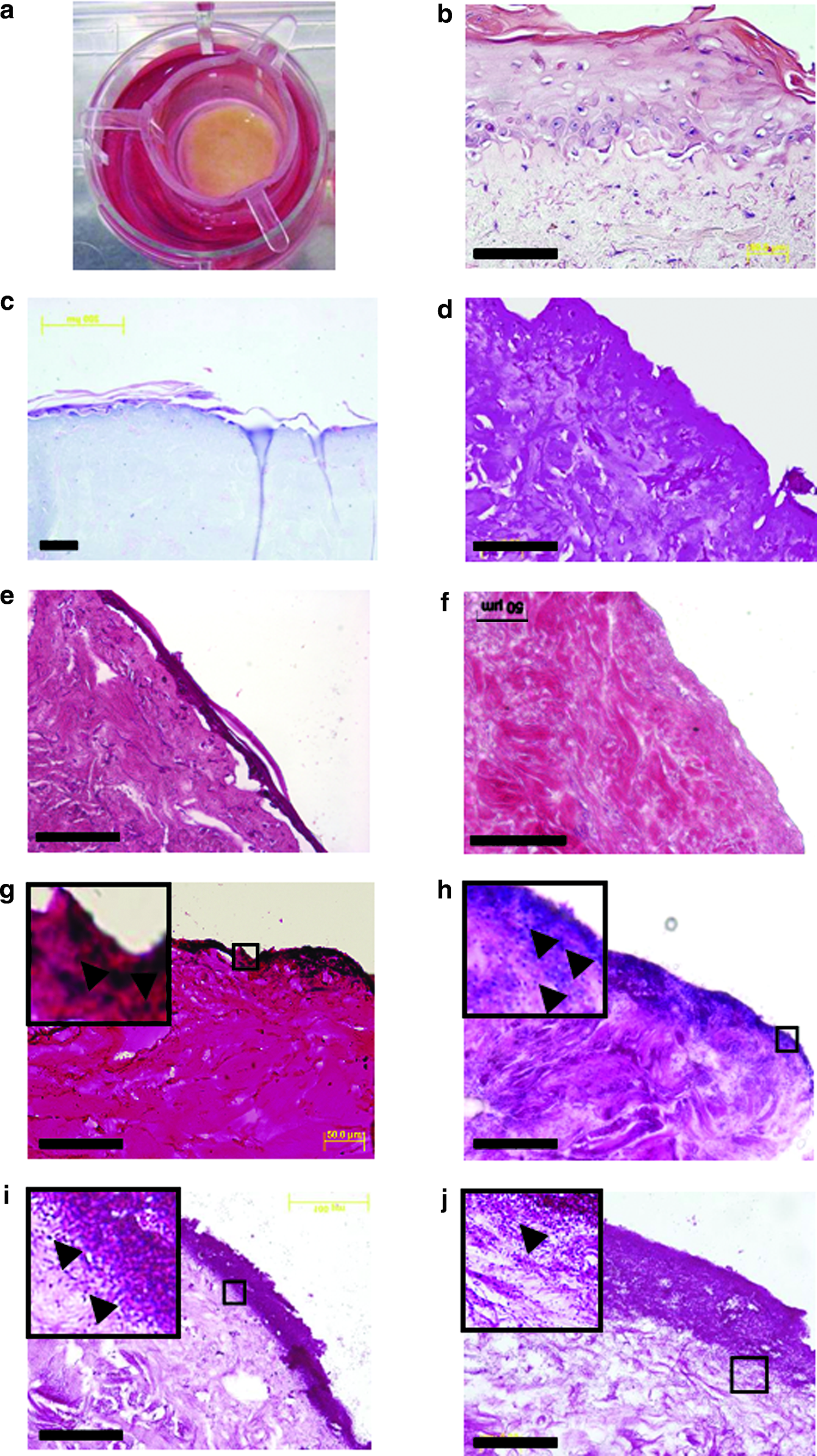
Creation and infection of small burn-wound skin constructs. Construct skin was cultured in tissue culture inserts suspended from the rim of wells in 12-well plates (
P. aeruginosa invades into the lower dermis over 72 h, whereas S. aureus remains in the superficial epidermis
In preliminary studies we employed the laboratory strain of S. aureus NCTC 6571 (Oxford) to infect the skin constructs; however, this strain infected only poorly, so we employed a clinical strain for all other studies. We presume that strain NCTC 6571 had lost full expression of virulence factors important for invasion of the skin model.
Burn-wound skin constructs were infected with 1 × 107 cfu P. aeruginosa, the clinical S. aureus strain, or PBS as control, and the tissue was then sacrificed for histology either 24 or 72 h postinfection (n ≥ 3 for each). Whereas S. aureus largely remained located within the epidermis and upper dermal layers (Fig. 2g, h), P. aeruginosa invaded the underlying dermis, even by 24 h (Fig. 2i, j). P. aeruginosa infection had a marked effect on the histological appearance of the dermis, rendering it less compact with large areas of apparently degraded tissue matrix, presumably due to the action of proteases.
Inoculation of the skin constructs with either strain of bacteria without prior burning of the epidermal surface did not result in bacterial invasion of the tissue. Histological examination of Gram-stained sections from intact skin infected with 1 × 107 cfu S. aureus (Fig. 2e) or P. aeruginosa (Fig. 2f) for 72 h revealed a lack of bacteria within the dermis and a relative paucity of bacteria in the epidermal layers. Viable counts of bacteria recovered from infected, unburnt tissue samples were extremely low compared to burnt samples infected with the same numbers of bacteria for the same time period; after 72 h infection, ∼1000 cfu/mg tissue of S. aureus and ∼200 cfu/mg tissue of P. aeruginosa were recovered from unburnt samples as compared to ∼40,000 cfu/mg and ∼70,000 cfu/mg, respectively, from the burnt tissue (Fig. 6).
Bacterial infection causes loss of epidermal keratinocytes
On histological examination of Gram-stained sections, skin constructs infected with P. aeruginosa appear to lack the epidermal layer, with a thick (around 30–40 μm) band of bacteria colonizing the upper layers of the dermis (Fig. 2g, h). In occasional Gram-stained infected skin sections, we observed a bacterial infiltrate beneath the epidermis, which appeared to be in the process of detaching from the dermis (Fig. 3a). To confirm loss of epidermis, we immunohistochemically stained sections of infected and noninfected skin constructs using antibodies against pancytokeratin and collagen IV. In uninfected sections, as expected, a well defined layer of keratinocytes was situated above the basement membrane (Fig. 3b, c). In infected sections, however, no keratin staining was visible (Fig. 3d), and the basement membrane also appeared to have been destroyed as evident by lack of collagen IV staining (Fig. 3e). Basement membrane was not lost due to the burn injury, because in uninfected, burnt samples, there was a clear line of demarcation of the keratinized epidermal layer but with a continuation of basement membrane (Fig. 3f).

Infection with P. aeruginosa results in loss of epidermis. (
P. aeruginosa migrates through the dermis
As it appeared that P. aeruginosa migrated efficiently through the infected tissue into the lower dermal layers in this model, we quantified the numbers of bacteria found deeper in the tissue samples at two time points postinfection. Histological examination of Gram-stained sections of whole skin constructs and microscopical counting of bacteria per field showed greater numbers of bacteria in deeper layers, further from the epidermis, after 72 h than after 24 h (Fig. 4a, b, and Fig. 5).

P. aeruginosa migrate into tissue over 72 h. Gram-stained sections of skin infected with P. aeruginosa for 24 h (
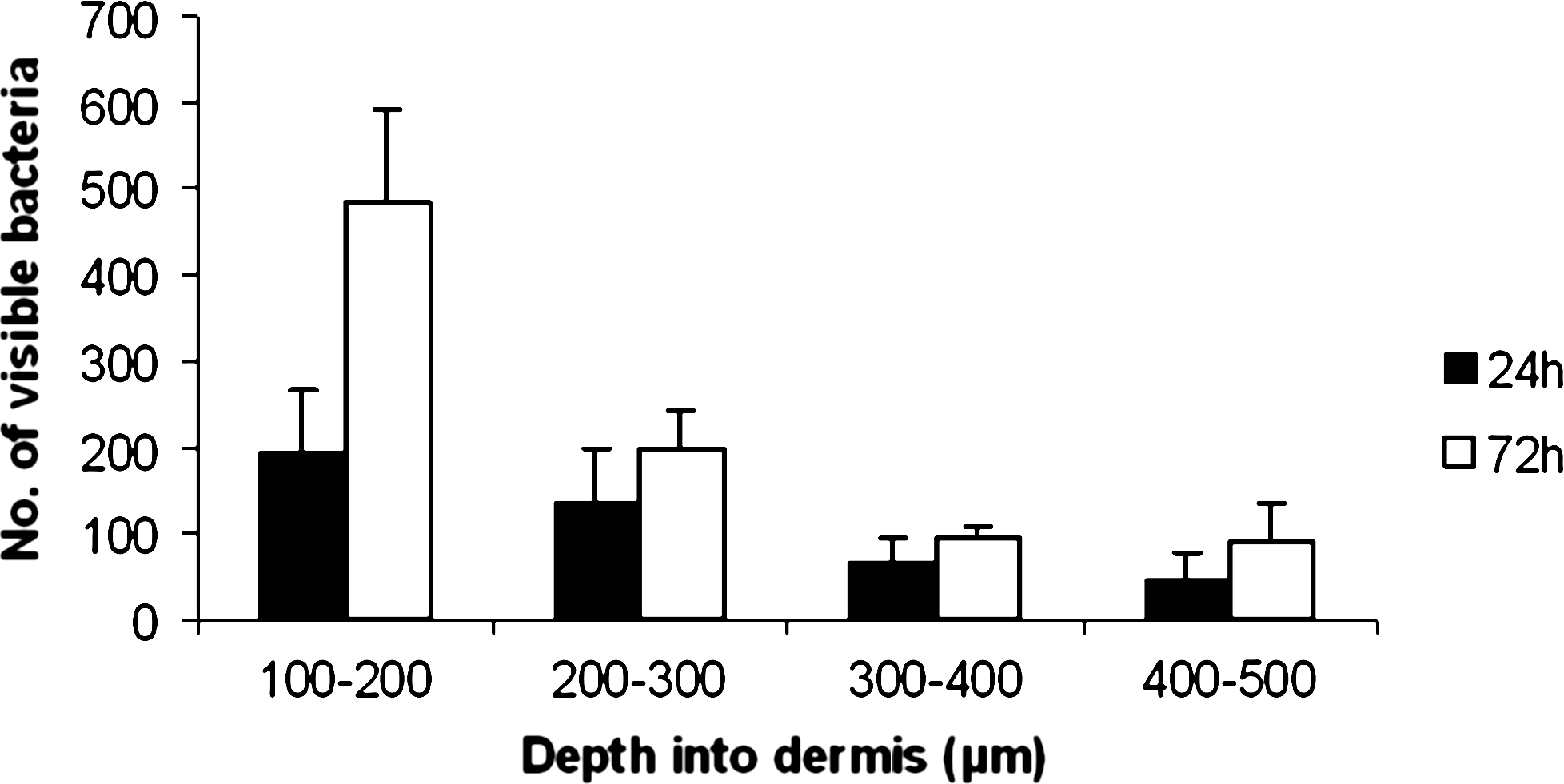
Microscopical quantification of bacterial load in deep layers of the dermis. 0–100 μm = TNTC. Histogram showing number of bacteria visible per × 60 field in relation to depth within the tissue. More P. aeruginosa were visible in deeper layers, further from the epidermis after 72 h than after 24 h. Error bars = SEM, n = 3 separate infected skin constructs for each time-point.
Bacteria multiply within the infected tissue
To determine whether bacteria introduced to the skin constructs were multiplying rather than simply migrating through the skin layers, we performed viable counts of bacteria recovered from the skin sections at two time-points postinfection (Fig. 6). We determined that both species were actively growing within the tissue, as would occur in native skin in vivo. This demonstrates that the species tested are unaffected by the tissue culture medium (Greens') and that there were enough nutrients available to sustain microbiological growth for a period of at least 72 h.

Viable bacterial counts in composite tissue postinfection. Homogenized tissue was serially diluted and cultured onto BHI agar plates to determine numbers of viable bacteria per mg of tissue. Increased numbers of viable bacteria were recovered from tissue after 72 h infection than after 24 h, indicating bacterial growth. Error bars = SEM, n ≥ 2 separate infected skin constructs for each time-point. Viable bacterial counts from unburnt infected samples were extremely low and are not shown on the graph; at 72 h postinfection, numbers of S. aureus recovered from tissue were ∼1000 cfu/mg, while numbers of P. aeruginosa were ∼200 cfu/mg.
Discussion
We have developed a three dimensional, easily reproducible model of human skin infection, using the common dermopathogens S. aureus and P. aeruginosa, which to our knowledge is the first of its kind.
Although S. aureus colonizes the skin and is present in the anterior nares of around 25–30% of healthy people,44–46 it is also the most common cause of skin and soft tissue infections, 47 and carriage is a risk factor associated with various morbidities and mortalities, including surgical complications. 44 The emergence of antibiotic resistant strains of S. aureus, as well as concurrent failure of antibiotic treatment of infection, points to a need for novel therapies, development of which could, in part, take advantage of the model proposed in this study.
Previously, investigations into skin infection have largely relied on small animal models. Although such studies have yielded significant amounts of information, there are many differences between, for example, rodent skin and human skin. Adult murine epidermal layers are thinner and contain much higher numbers of densely packed hair follicles than human. 48 Use of porcine models better approximates human skin and its infection49–53 but also has limitations such as the high cost. Both animal models raise ethical questions about the numbers of animals that are needed for research, so an in vitro model that reduces the extent of animal experimentation is very useful.
In their early porcine infection model, Mertz et al. concluded that an initial innoculum of 1 × 108 cfu/mL of P. aeruginosa or S. aureus provided maximal attachment of bacteria to the wound bed, which is the bacterial concentration used here. 49 Interestingly, studies by both Davis et al. 53 and Svedman et al. 50 demonstrated that in the porcine model, S. aureus infection was localized to the wound without apparent systemic infection. In our model, after 72 h infection S. aureus appeared to remain largely in the upper layers of the skin, whereas P. aeruginosa did clearly penetrate with greater efficiency into deeper tissue. The latter could be due to the release of Pseudomonas elastase and alkaline proteases, which are capable of degrading collagen and have elastolytic activities. 54 The digestion of structural proteins of the skin could enhance the invasive capabilities of P. aeruginosa. The use of in vivo models of skin contamination, such as an early murine model studying protease-deficient P. aeruginosa in infection of burn wounds, 55 has established that P. aeruginosa protease markedly enhances invasiveness of the organism.
In addition to invasion, infection with P. aeruginosa appeared to result in a loss of epidermal layers. We observed areas in our histological sections where the bacteria invade underneath the epidermis (Fig. 3a). We propose that the bacteria gain entry to the dermis via either the burn wound or the exposure of dermis in the larger skin constructs, and then extend both vertically and horizontally at the epidermal–dermal junction. In the engineered skin construct, this is potentially the weakest point. Burning of the skin constructs resulted in a partial loss of epidermis (Fig. 2c, 3f), which accounted for a portion of the epithelial damage, but did not explain the total loss observed in skin that was both burnt and infected. Incursion of bacteria along the epidermal–dermal junction and the consequent release of proteases could instigate a weakening of the junction to the extent that the epidermis peels away, which was in fact seen. In samples where the epidermal layer was not burnt before infection and therefore remained intact, bacteria were not visible within the dermis on Gram staining of sections. Additionally, numbers of viable bacteria recovered from the tissue even 72 h postinfection were extremely low. This indicates a requirement for a break in the epidermal barrier before effective bacterial infection and invasion of the tissue can occur, comparable with the situation in vivo.
After infection, the bacterial species used in this model thrived for at least 72 h within the tissue, indicating sufficient availability of nutrients. The lowest viable count of bacteria recovered from infected tissue was equivalent to around 2 × 107 cfu/g tissue, which is well above the criterion for infection of wounds proposed by Loebl et al. of >1 × 105 cfu/g tissue.56,57
In the burn wound model, we have created a reasonably economical system for studying infection by culturing skin within small inserts. A feature of the tissue culture inserts is a porous membrane at the base, through which medium bathes the dermis while keeping the epidermis at the air–liquid interface. Future applications of the model could include the introduction of different components of the innate immune system, such as lymphocytes, chemokines, or cytokines, to the infection model via the subdermal pores. This would allow the manipulation and study of discreet or mixed immune system components in the infection setting, potentially providing a wealth of information. Recently, McFarland et al. 58 found that epidermal cells in their skin construct model, which differs by having a collagen–polymer scaffold rather than a dermal base, reacted to inflammatory cytokine insults such as IL-1α, TNF-α, and IL-17 by upregulating expression of genes involved in both innate and adaptive immunity, including those encoding anti-microbial defensins. That engineered construct cells behave similarly at the gene expression level to native cells in their study is encouraging in terms of the realistic results potentially achievable with our model.
The model described here will allow researchers to collect information on the interactions and responses of skin cells and microorganisms to each other and the tissue matrix in a three-dimensional environment. In its present form the model is already being used to study approaches to reduce the extent of bacterial burden in infected wounds, and in the future it could be used to develop targeted anti-bacterial therapies and diagnostic tests.
Footnotes
Acknowledgments
This research was supported by a grant from the Engineering and Physical Sciences Research Council and the Defence Science and Technology Laboratory.
Disclosure Statement
No competing financial interests exist.