
Abstract
The use of bioartificial livers (BALs) for the expansion of human adult liver stem cells and the production of growth factors could be a potential strategy for cell-based extracorporeal liver support. The present study aimed to assessing the differentiation of human adult liver stem cells in a rotary BAL. Liver stem cells were seeded into a polysulphone membrane filter at a density of 3 × 108 cells, and the filter was connected to a rotary bioreactor perfusion system (37°C, 50 mL/min, 48 h). Viability, cell differentiation, and metabolic performances were evaluated at 24 and 48 h. Hepatocyte growth factor production from human adult liver stem cells, mature hepatocytes, and mesenchymal stem cells in adhesion and in the rotary BAL conditions was compared. Liver stem cells cultured in the rotary BAL produced the highest amounts of albumin (p = 0.002) and ammonia-induced urea (p = 0.0001), and had an increased cytochrome P450 expression in respect to liver stem cells in adhesion. Remarkably, liver stem cells in the rotary BAL produced very high amounts of hepatocyte growth factor (p = 0.005) in respect to hepatocytes and mesenchymal stem cells. Moreover, the cells lost their stem cell markers and acquired several markers of mature hepatocytes. In conclusion, the rotary BAL favored liver stem cell differentiation into mature hepatocyte-like cells.
Introduction
Recent studies have allowed to recognize in many tissues the presence of resident stem cells that are normally dormant but which can be activated to trigger a full regenerative program under appropriate stimulation. 7 We recently identified a human adult liver resident stem cell (HLSC) population with multipotent differentiation capabilities and regenerative properties. 8 HLSC were characterized by the expression of mesenchymal markers (CD29, CD90, CD73, and CD44) and embryonic stem cell markers (nanog, Oct-4, Sox2, and SSEA-4) and by the absence of hematopoietic (CD45, CD34, CD117, and CD133) or oval stem cell cytokeratin-19 (CK19) markers. 8 In vitro differentiation into mature hepatocytes was made possible only in the presence of a combination of several growth factors such as fibroblast growth factor-4 (FGF-4) and hepatocyte growth factor (HGF).
Various bioartificial liver (BAL) systems have been proposed using different geometries and cell types.9–13 The bioartificial support is commonly considered to provide temporary support until either regeneration, “bridging to regeneration,” is possible or until a donor liver can be found and transplanted, “bridging to transplantation,” thus improving survival of patients with severe liver disease.10,14 However, to date the real clinical benefit of BALs is still questioned. 15
The majority of studies on BAL support is focused on the metabolic performances of a given cell type in the device in view of its potential clinical application.
In this study, we evaluated whether a rotary BAL could be a tool to differentiate the HLSC into mature hepatocytes. To this aim, we evaluated HLSC viability, differentiation, and metabolic performances in BAL in comparison with HLSC cultured in adhesion, human hepatocytes, and mesenchymal stem cells (MSC).
Materials and Methods
Isolation and characterization of HLSC
HLSC isolation and characterization was performed as described. 8 Human cryopreserved normal hepatocytes obtained from Cambrex (Bio Science, Verviers, Belgium) were used. Briefly, hepatocytes were seeded at a density of 1.0–1.5 × 105 viable cells (80% viable cells determined by the trypan blue [Invitrogen, San Diego, CA] per cm2 onto collagen-coated culture plates in Hepatozyme-SFM maintained at 37°C, 5% carbon dioxide [CO2] for 2 weeks). After 2 weeks in culture, hepatocytes died and then medium was substituted by alpha-minimum essential medium/endothelial cell basal medium-1 (3:1) (Gibco–Invitrogen/Lonza Walkersville, Walkersville, MD) supplemented with L-glutamine (5 mM), HEPES (12 mM, pH 7.4), penicillin (50 IU/mL), streptomycin (50 μg/mL) (all from Sigma-Aldrich, St. Louis, MO), fetal calf serum (FCS) (10%), and horse serum (10%; Gibco–Invitrogen). The expanded cells were transferred to a 75-cm2 flask and analyzed when they approached confluence. Characterization of HLSC was performed by cytofluorometric analysis and the following antibodies, all phycoerythrin-conjugated or fluorescein isothiocyanate–conjugated, were used: anti-CD105, -CD29, -CD146, -CD44, -CD73, -CD90, -ICAM-1, -EpCAM, -NCAM, and P450 and phycoerythrin-conjugated or fluorescein isothiocyanate–conjugated goat antibodies against mouse immunoglobulin G (IgG) secondary antibody when needed. For each sample, 105 cells were analyzed on a fluorescence-activated cell sorting (FACScan) cytometer. Gating was constructed based on negative controls, and compensation controls were included in all analyses performed. Population percentages and numbers were generated for gated populations from each experiment using Cell Quest software. Indirect immunofluorescence was also performed on HLSC cultured on chamber slides (Nalge Nunc International, Rochester, NY), fixed in 4% paraformaldehyde containing 2% sucrose, and, when needed, permeabilized with HEPES-Triton ×100 buffer. The following monoclonal antibodies (mAb) were used: mAb anti-alpha-fetoprotein (α-FP), mAb anti-albumin (R&D Systems, Abington, United Kingdom), mAb anti-CK18 (Chemicon International, Temecula, CA), and mAb anti-CK8. Omission of the primary mAb or substitution with an unrelated rabbit serum or mouse IgG was used as controls. Alexa Fluor 488 anti-rabbit IgG and Texas Red anti-mouse IgG (Invitrogen) were used as secondary Ab. Confocal microscopy analysis was performed using a Zeiss LSM 5 Pascal Model Confocal Microscope (Carl Zeiss International, Jena, Germany). Hoechst 33258 dye (Sigma-Aldrich) was added for nuclear staining.
MSC isolation and culture
Human MSC were isolated from bone marrow. 16 Briefly, about 20–30 mL of bone marrow aspirate was collected into a syringe containing 6000 U of heparin to prevent clotting. The bone marrow samples were washed twice with Dulbecco's phosphate-buffered saline (PBS) (Invitrogen), and recovered by centrifugation (at 900 g, 10 min, 20°C). Up to 2 × 108 nucleated cells at a 4 × 107 cells/mL density were loaded onto 25 mL of Percoll (density: 1.073 g/mL in a 50-mL conical tube). Cell separation was accomplished by centrifugation (1100 g, 30 min, 20°C). The nucleated cells were collected from the interface, diluted with two volumes of Dulbecco's PBS (Gibco–Invitrogen), and collected by centrifugation. The cells were resuspended, counted, and plated at 200,000 cells/cm2. The cells were cultured in Dulbecco's modified Eagle's medium (Invitrogen) containing 10% fetal bovine serum (FBS).
The rotary BAL system
In the setup of BAL the medium circuit driven by a peristaltic pump regulates circulation consisting of a set of tubes linked to a reservoir for fresh medium, a waste circuit, air traps, and an oxygenator. The total priming volume was 180 mL. The oxygenator, a gas exchange membrane, enables contact between the medium and the oxygen, nitrogen, and CO2 mixture, allowing medium oxygenation and pH maintenance. The main reactor housing contained the oxygenator, parts of the medium circuit, and the rotation unit. The filter was made of packed Albuflow® (cut-off: 340 kDa; Fresenius Medical Care, Bad Homburg, Germany) hollow fibers (number: 500) (priming volume: 20 mL). The cellular volume was 80 mL. A heating device regulated by a thermostat ensured stable temperature conditions inside the reactor housing. The chamber was flushed with medium, and the bubble point test for integrity of structure was performed before cell inoculation, was placed into the reactor's rotation unit, and linked to the medium circuit.
In vitro culture in BAL
For the experiments in the BAL, HLSC were harvested by trypsinization and were seeded at a density of 300 × 106 on a polysulphone membrane filter that was connected to a bioreactor perfusion system in which temperature was maintained at 37°C and the hollow fiber space was perfused with culture medium at 50 mL/min of flow rate. Then, 1 mM of ammonia was added to the rotary BAL culture. In control experiments, a monoclonal anti-human HGF blocking antibody (MAB294, clone 24612; R&D System) was added to the BAL culture to neutralize HGF production. Culture medium pH was monitored between 7.4 and 7.5. This perfusion system was maintained for 48 h, and aliquots of medium were recovered at 24 and 48 h to evaluate metabolites. In control experiments MSC and human mature hepatocytes were harvested by trypsinization and were seeded in BAL in the same condition.
Metabolic analysis
Albumin production was analyzed by the QuantiChrom BCG albumin assay kit (Gentaur, Brussels, Belgium), a quantitative colorimetric method, according to the manufacturer's instructions. Briefly, 5 μL of diluted standards and collecting medium of HLSC was transferred to a 96-well plate in triplicate, and 200 μL of working reagent containing bromocresol green, which forms a colored complex specifically with albumin, was added. After incubation, the intensity of the color was measured at 620 nm. Albumin concentration was calculated as:
where a and b were found fitting the standard curve using the equation y = ax/(b + x). To exclude bovine albumin interference the absorbance of control medium supplemented with FCS was subtracted to the absorbance of each sample.
Ammonia-induced urea production was quantified by the QuantiChrom Urea assay kit (Gentaur), according to the manufacturer's instructions. Briefly, 50 μL of water (blank), 50 μL of diluted standard (5 mg/dL of urea), and 50 μL of samples were transferred into wells of a clear bottom 96-well plate in triplicate, and 200 μL of working reagent containing a chromogenic reagent was added. After incubation, the intensity of the color was measured at 520 nm. Urea concentration of samples was calculated as:
where [STD] = urea standard concentration (5 mg/dL).
Glucose consumption was determined using the SYNCHRON CX5 (Beckman Coulter, Fullerton, CA) and expressed as T0 concentration −T24 h (or T48 h concentration), where T0, T24, and T48 h were the concentration of glucose at time 0, 24, and 48 h, respectively. The glucose concentration of the medium was 95 ± 9.21 mg/mL at time 0.
Lactate dehydrogenase (LDH) release was measured by cytotoxicity detection kit (Roche Applied Science, Monza, Italy), according to the manufacturer's instructions. Briefly, 100 μL of water (blank), 100 μL of diluted standard (0.1 U/mL of LDH from Sigma-Aldrich), and 100 μL of cell-free supernatants were transferred into wells of a clear bottom 96-well plate in triplicate and incubated with the substrate mixture from the kit. The intensity of the color was measured at 520 nm. LDH concentration was calculated as:
where [LDH] = LDH standard concentration (0.1 U/mL). The increase in supernatant LDH activity directly correlated with the number of dead cells.
Indocianin green uptake assay
The uptake and the secretion of Indocianine green (ICG; Sigma-Aldrich) is a specific function of mature hepatocytes. To evaluate the liver differentiation of HLSC after culture in BAL, the ICG uptake was performed. Briefly, after washing in PBS (Sigma-Aldrich), ICG solution was added to the cells at a final concentration of 1 mg/mL and incubated for 15 min at 37°C, 5% CO2. Then, cells were washed three times in PBS and the plates were refilled with alpha-minimum essential medium/endothelial cell basal medium-1 (Invitrogen/Lonza Walkersville, MD) containing 10% fluorescence-activated cell sorting (FCS). ICG was completely eliminated from the cells in the supernatants after 30 min.
Periodic acid solution reaction
Glycols are oxidized to aldehydes when treated with periodic acid (Sigma-Aldrich), and after reaction with Schiff's reagent (Sigma-Aldrich), a mixture of pararosaniline and sodium metabisulfite, a pararosaniline adduct is released that stains the glycol-containing cellular components. Periodic acid solution (PAS) reaction was performed on paraffin-embedded tissue section of aggregates recovered after BAL culture. Briefly, sections were deparaffinized, hydrated, and incubated in periodic acid solution for 5 min at room temperature. After several rinsing, sections were incubated for 15 min at room temperature in Shiff's reagent. Sections were counterstained in hematoxylin solution for 90 s, washed to remove reagents, and examined by image acquisition with Zeiss Axioscope (Carl Zeiss International).
Fluorimetric determination of P450 activity
Fluorescence-based P450 assays were performed by direct incubation of cells cultured in rotary BAL with substrate 7-ethoxy-4-trifluoromethylcoumarin, supplied by (Ultrafine Chemicals, Manchester, United Kingdom). Substrates were added in acetonitrile solution in concentrations for which the final solvent concentration did not exceed 0.5% (v/v) when added to incubation medium. After 24 and 48 h at 37°C, the metabolite formed during the activity assays, 7-hydroxy-4 trifluoromethylcoumarin, were evaluated hydrolyzing by incubation of supernatants with β-glucuronidase/arylsulfatase (150 Fishman units/mL and 1200 Roy units/mL, respectively) for 2 h at 37°C. Finally, samples were diluted (1:2) in the corresponding quenching solution (0.25 M Tris in 60% [v/v] acetonitrile), and fluorescent metabolite formation was quantified fluorimetrically by means of a Spectra Max Gemini XS fluorescence microplate reader at 510 nm (Molecular Devices, Sunnyvale, CA). Results were expressed as nanomoles of metabolite formed per hours per 106 cells.
Quantitative real-time polymerase chain reaction
First-strand cDNA was produced from 2 μg RNA using high-capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA). Briefly, 2 μg of mRNA, 2 μL of RT buffer, 0.8 μL of dNTP mixture, 2 μL of RT random primers, 1 μL of MultiScribe reverse transcriptase, and 4.2 μL of nuclease-free water were used for each cDNA synthesis. After the reverse transcription, cDNA was stored at −20°C. Relative quantization by real-time polymerase chain reaction (PCR) was performed using SYBR-green detection of PCR products in real-time using a 48-well StepOneTM real-time system (Applied Biosystems). Sequence-specific oligonucleotide primers (purchased from MWG-Biotech AG, Ebersberg, Germany) were as follows:
HNF4—forward, 5′-TGCGTGGTGGACAAAGACAA-3′; reverse, 5′-TCTGGACGGCTTCCTTCTTC-3′ C/EBP—forward, 5′-GCACACTGTGGGCAGACAGCCATG-3′; reverse, 5′ CTCAAGGCTATCTTTGTTCACTGC-3′.
Power SYBR® Green PCR Master Mix was purchased from Applied Biosystems. Thermal cycling conditions were as follows: activation of AmpliTaq Gold® DNA polymerase LD at 95°C for 10 min, followed by 45 cycles of amplification at 95°C for 15 s and 60°C for 1 min. To detect the log phase of amplification, the fluorescence level (quantification of product) was determined at each cycle. The cycle at which the fluorescence reached threshold was recorded, averaged between triplicates, and normalized to the averaged cycle of threshold value for glyceraldehyde 3-phosphate dehydrogenase.
Drug metabolism PCR array
Drug metabolism quantitative real-time PCR was used to analyze cytochrome mRNA expression in HLSC and human mature hepatocytes. Assay was purchased from SuperArray (SuperArray Bioscience Corporation, Frederich, MD) and was performed following the manufacture's instructions. Briefly, 1 μg RNA was retrotranscribed using RT first-strand kit; once the cDNA was obtained, it was mixed to RT real-time PCR Master Mix containing SYBR green as reference dye and was dispensed in aliquots across the PCR plate containing a panel of 96 primer set for a thoroughly researched set of 84 relevant pathway- or disease-focused genes, plus five housekeeping genes and three RNA and PCR quality controls. Thermal cycling conditions were as follows: 95°C for 10 min, followed by 40 cycles of amplification at 95°C for 15 s and 60°C for 1 min. Data analysis template was downloaded from Superarray Web site (
Cytokines secretion
To evaluate HGF secretion, an aliquot of medium was recovered after 24 and 48 h and analyzed by antibody sandwich enzyme-linked immunosorbent assay for the quantitative determination of human HGF concentration (R&D Systems, catalog number: DHG00) according to the manufacturer's instructions.
Statistical analysis
Data are presented as mean ± standard deviation. Statistically significant differences were determined using t-test or Newmann-Keuls multicomparison test when appropriate. A p-value <0.05 was considered significant.
Results
Isolation and characterization of HLSC
HLSC were obtained from cryopreserved hepatocytes by culturing under stringent conditions. HLSC characterization was performed by FACS and immunofluorescence. As shown in Figure 1A, HLSC expressed several stem cell markers as CD29, CD105, CD90, CD73, CD44, and CD146; HLSC also expressed hepatocyte-specific markers such as albumin, CK8, α-FP (Fig. 2), which is considered a marker of hepatocyte precursors, and CK18 (Fig. 2). Moreover, HLSC expressed ICAM-1, but not EpCAM and NCAM (Fig. 1A). As previously described by Herrera et al., 8 HLSC were negative for hematopoietic stem cell markers CD45, CD34, CD117 (c-kit), and CD133 and for CK19, a marker of oval cells (data not shown). These results indicated that HLSC expressed markers of both MSC and immature liver cells (α-FP). Absence of CK19 expression suggested that HLSC are a population of liver progenitor cells different from oval cells.

Fluorescence-activated cell sorting (FACS) analysis of HLSC in adherent culture condition and in BAL. (
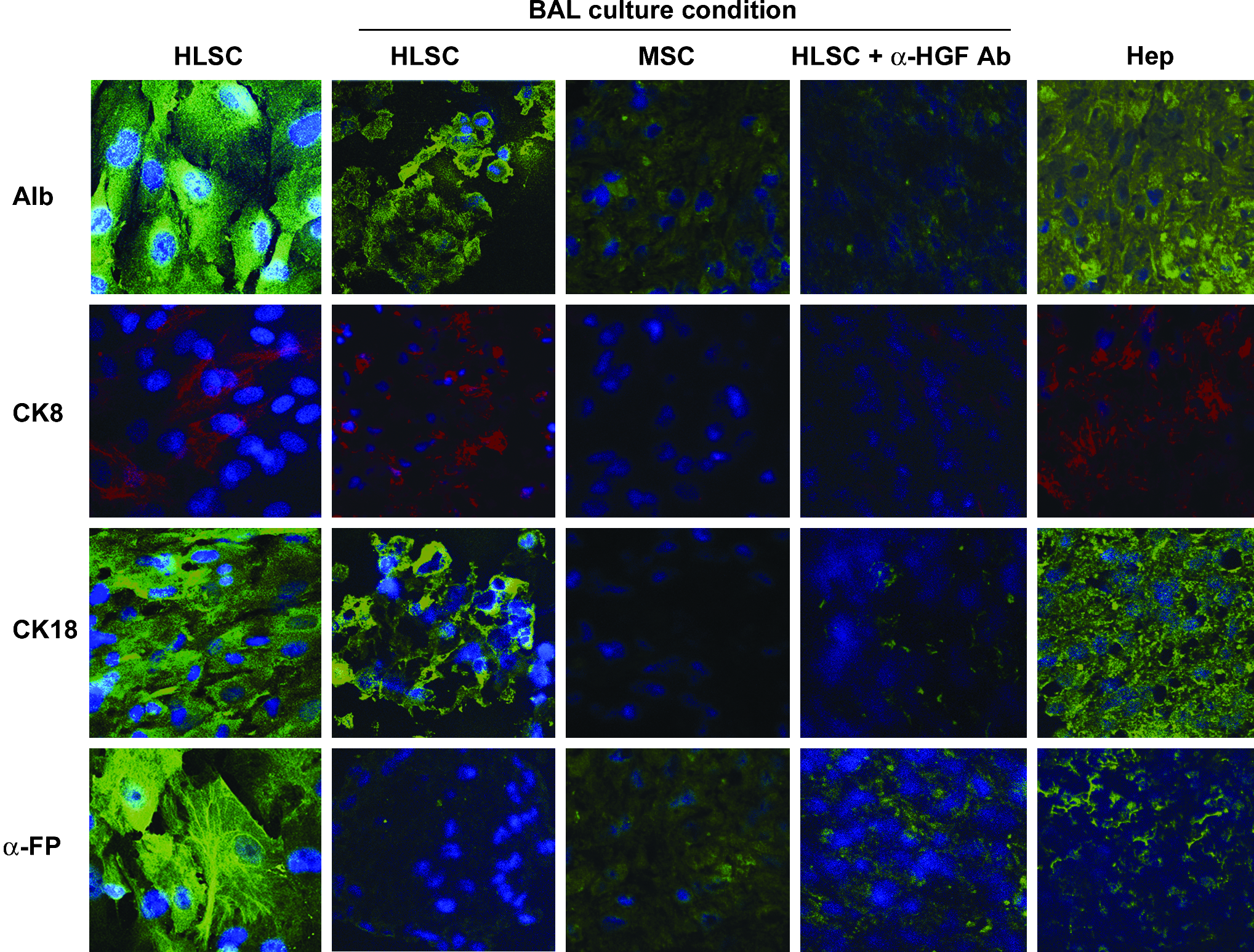
Immunofluorescence analysis of HLSC, MSC, and human hepatocytes. Hepatic differentiation of HLSC was evaluated by immunofluorescence on HLSC both in adherence and BAL culture and compared with MSC in BAL culture and human hepatocytes. In some experiments HLSC were incubated with a monoclonal blocking antibody against HGF. Cells were stained with antibodies against albumin, CK8, α-FP, and CK18. The nuclei of the cells were stained with Hoechst dye 33258. Original magnification, × 400. MSC, mesenchymal stem cells; HGF, hepatocyte growth factor; CK, cytokeratin, FP, fetoprotein.
High-density HLSC culture in BAL
We compared the ability of HLSC to differentiate into mature hepatocytes in two different conditions (BAL vs. adhesion) in the presence of the same culture medium with no addition of specific differentiation growth factors (HGF and FGF-4). In the BAL, the seeded cells formed aggregates within 24 h. HLSC cultured in BAL for 48 h lost MSC markers such as CD73, CD146, CD29, CD105, and CD44 (Fig. 1B), and cells acquired the expression of cytochrome P450 activity, that was not present in HLSC in adhesion, as evaluated by FACS analysis (Fig. 3B). Moreover, by fluorimetric determination of the metabolism of the 7-ethoxy-4-trifluoromethylcoumarin substrate for P450 indicated that HLSC cutured in BAL, but not in adhesion, converted the substrate in a manner comparable to that of human hepatocytes (Fig. 3A). By RT-super array, we observed a comparable expression of CYP11B2, CYP17A1, and CYP2F1 in HLSC cultured in BAL and in human hepatocytes. CYP19A1 was more expressed by HLSC than by human hepatocytes, whereas CYP1A1, CYP2B6, CYP2C19 CYP2C6, CYP2C9 CYP2D6, CYP2E1, CYP2J2, and CYP3A5 were more expressed in human hepatocytes. Moreover, HLSC after culture in rotary BAL maintained the expression of liver tissue–specific markers such as albumin, CK8, CK18, and down-regulated α-FP (Fig. 2). MSC cultured in the same condition showed a slight expression of albumin and α-FP but not CK8 and CK18 (Fig. 2). HGF blockade in HLSC cultured in BAL inhibited the expression of albumin, CK8, and CK18 (Fig. 2). Moreover, by quantitative real-time PCR, the HLSC cultured in BAL expressed both liver-specific transcription factors hepatocyte nuclear factor 4 and C/EBP mRNA. HGF blockade inhibited the expression of HFN-4 but not C/EBP. MSC cultured in BAL expressed slight amount of C/EBP, but not hepatocyte nuclear factor 4. High positivity for proliferating cell nuclear antigen indicated robust cell proliferation (Fig. 4A) in the absence of LDH releases in the supernatants (0.01 ± 0.01 in 24 h and 0.02 ± 0.02 in 48 h mU/h/106 HLSC) as shown in Table 1. In addition, HLSC cultured in rotary BAL showed periodic acid solution (PAS) positivity (Fig. 4B) and ICG retention and release (Fig. 4C, D) that were not characteristics of HLSC cultured in adhesion. As previously described by Herrera et al., 8 HLSC in adhesion differentiate only after addition of growth factors into the culture medium.

Cytochrome P450 expression and activity in HLSC after culture in rotary BAL. (
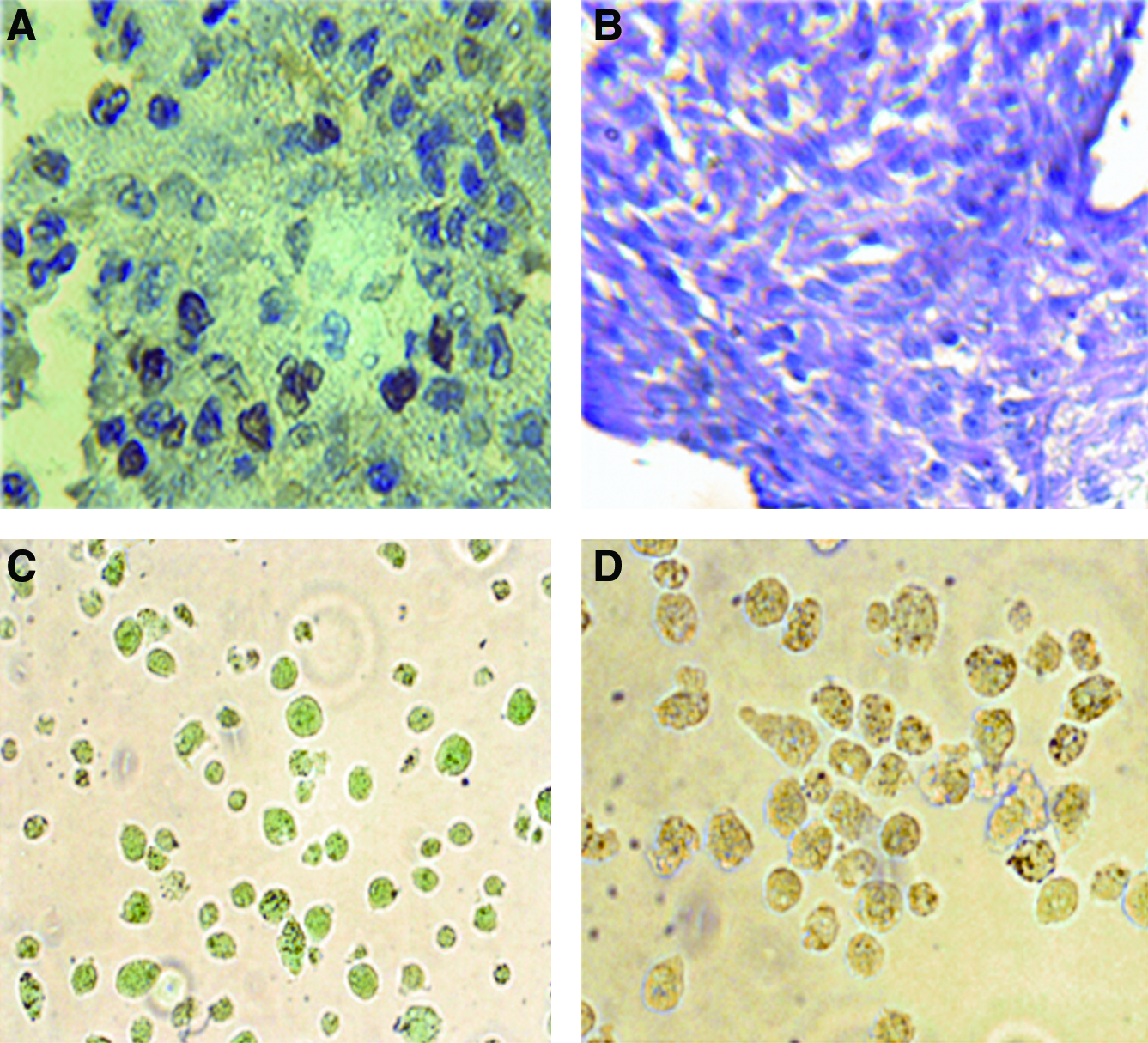
Immunocytochemistry analysis, HGF production, and expression of hepatocyte nuclear factor 4 and C/EBP mRNA of differentiated HLSC, (

Comparison between HLSC in adherence culture and HLSC, MSC, and human hepatocytes in rotary BAL in terms of metabolic activity displayed as glucose consumption, urea production, albumin secretion (all expressed as μg/h/106 cells), LDH release (mU/h/106 cells), and HGF production (ng/mL/106 cells). Values are means ± standard deviation. Statistical analysis was performed using Student's t-test.
LDH, lactate dehydrogenase; HGF, hepatocyte growth factor; BAL, bioartificial liver; MSC, mesenchymal stem cells; HLSC, human adult liver resident stem cell.
Production of metabolites
As shown in Table 1, glucose consumption indicated an active metabolism of HLSC cultured in BAL in the absence of a significant LDH release, as a marker of cell lysis. The acquisition of mature hepatocyte functions was indicated by the secretion of albumin and production of urea comparable to that of mature human hepatocytes cultured in the same condition. In adherent culture conditions, HLSC were not able to produce urea in the absence of growth factors, but they secreted low amounts of albumin. MSC cultured in BAL had a comparable glucose consumption but lower secretion of albumin and production of urea.
HGF secretion
HLSC, at variance to hepatocytes and MSC, secreted 81.36 ± 0.6 in 24 h and 232.8 ± 2.2 ng/mL/106 cells of HGF when cultured in BAL, whereas the amount of HGF produced by HLSC was significantly lower when they were cultured in adhesion (4.56 ± 0.03 in 24 h and 23.40 ± 0.02 in 48 h ng/mL/106 cells) (Table 1 and Fig. 4). The amount of HGF produced by MSC and human hepatocytes cultured in BAL was significantly lower than that of HLSC cultured in BAL.
Discussion
The results of the present study indicate that HLSC spontaneously differentiate into a mature phenotype when cultured in a rotary BAL. Contrary to adhesion culture conditions, the culture in rotary BAL offered a better microenvironment for the cells. These results may be due to the lack of compartmentalization of metabolites by low gravity due to rotation ensuing in a more uniform concentration of endogenous growth factors.
HLSC is an adult pluripotent stem cell population characterized by the expression of mesenchymal markers (CD29, CD90, CD73, and CD44), embryonic stem cell markers (nanog, Oct-4, Sox2, and SSEA-4) and by the absence of hematopoietic or oval stem cell markers. 17 Indeed, they did not express CD45, CD34, and CD133 and CK19, which is a marker of oval cells. Moreover, the HLSC grow for up to 30 passages in culture, whereas the oval cells have not been so far isolated and cultured from human liver. In vitro differentiation into mature hepatocytes was made possible only in the presence of a combination of several growth factors such as FGF-4 and HGF. Further, HLSC were shown to differentiate into osteogenic, adipogenic, endothelial, and insulin-producing cells when cultured in appropriate conditions. 17
This study showed that after 48 h in the rotary BAL, HLSC, purified as previously described from human isolated cryopreserved mature hepatocytes, acquired markers typically expressed by mature hepatocytes like cytochrome P450, maintained the expression of liver tissue–specific markers such as CK18, CK8, and albumin, and lost fetal liver markers, that is, α-FP and MSC markers (CD73, CD146, CD29, CD105, and CD44). In the adhesion culture condition, none of these phenotypic changes occurred.
This study showed that at 24 and 48 h in the rotary BAL, HLSC could perform metabolic activities such as glucose consumption and cleavage of the CYP450 substrate that is typical of mature hepatocytes. A very relevant finding was the strikingly increased production of albumin already at 24 h, which was even more prominent at 48 h. Under the adhesion culture condition, only glucose was metabolized at a significantly lower rate than in rotary BAL (at 48 h). No production of urea and cleavage of the CYP450 substrate were observed. Albumin was produced but only in very low quantities.
Since HGF is known to favor liver regeneration, 17 we hypothesized that it would be important to test the possibility that the production of HGF may be enhanced in the low-gravity conditions of the rotary BAL. A remarkable result was the prominent production and release of HGF by HLSC in rotary BAL but not in adherence culture conditions. The quantitative difference was 20-fold higher in HLSC in rotary BAL than in adhesion. Moreover, the comparative analysis of HGF production among HLSC in rotary BAL, and in adhesion culture condition versus other cell types, that is, MSC and mature hepatocytes clearly showed that HLSC are able to produce the highest amount of HGF. HGF blockade inhibited differentiation of HLSC in BAL, suggesting an autocrine action of HGF in HLSC differentiation. HGF is a heterodimer molecule composed of the 69 kDa α-subunit and the 34 kDa β-subunit. It would therefore be that HGF could pass by simple diffusion from the cellular into the circuit. Wurm et al. 18 using the same rotary BAL with setups differing in size and layout of exchange area, found that mass transfer time for urea and several ions was around 30 and 50 min, whereas complete exchange of albumin was delayed up to 20 h. These data highlight the dependence of mass transfer on both area of exchange as well as size of molecule. Based on our studies, the final concentration (at 48 h) in the cellular and circuit compartments would be 69.120 μg/180 mL (0.384 μg/mL), considering a 50% equilibration between the two compartments.
Taken together, these results indicate that HLSC, but not MSC, differentiated in BAL paralleled human hepatocytes for metabolism, secretion of albumin, production of urea, CYP450 activity, and expression of liver-specific transcription factors HFN-4 and C/EBP, but differed for the pattern of cytochromes expressed as evaluated by super array and for the production of HGF.
Further studies are, however, needed to evaluate the rate of mass transfer of this molecule as well as other potentially relevant molecules in liver regeneration, for example, chemokines and other growth factors.
Our studies may suggest that HLSC could be a potential cell source for BAL. Human hepatocytes should be the optimal cell for BAL devices. However, the shortage of donor tissue usually renders this approach impractical. Several groups have investigated BAL devices using normal or immortalized liver cells from various mammalians.19–22 A remarkable trend toward the use of porcine hepatocytes23–25 is noticeable, mainly because of the similarity in their physiology to that of humans and the abundance of available organs.
A few years ago, it was reported that researchers were able to isolate sufficient amounts of viable human hepatocytes from organs not suitable for transplantation 26 (also reviewed in Refs.12,27). However, no further progress in this area was reported.
Our recent identification and characterization of HLSC from normal adult livers has shown that dormant resident pluripotent stem cells exist in the hepatic parenchyma in a dormant state. 8 The expansion of these cells in standard adhesion conditions requires growth factors and a lag phase. Our data clearly show that HLSC are able to undergo proliferation and differentiation into mature hepatocytes, together with the ability to express their full metabolic activity.
The results bear a potential application for rotary BAL as a bridge to regeneration. The production and release of regenerative factors such as HGF with concentration not otherwise achievable may be very relevant in clinical medicine. In our study, the acquisition of mature hepatic phenotype without the need of exogenous-specific differentiation growth factors may be relevant in cutting costs for the application of this device.
Our study has some limitations. Our observation was limited to 48 h. Differentiation and HGF production were already evident at 24 h and more pronounced at 48 h, a time frame that could allow a potential clinical application even under urgent conditions. Another limitation is that the results obtained with HLSC in the rotary BAL may be not exportable to another BAL device that may offer different microenvironmental conditions that may be optimal for mature hepatocytes but may be not for the growth and differentiation of HLSC.
Finally, the rotary BAL has implied that resident stem cells such as the HLSC, despite their multipotency, may be in fact committed to hepatocytes under appropriate favorable conditions.
In conclusion, we have shown that liver stem cells can acquire the phenotypic and metabolic features of mature hepatocytes when cultured in a low gravity, rotary BAL. These properties may be exploited as a device to provide regenerative factors in clinical application or as laboratory tool to differentiate HLSC or possibly other cell types.
Footnotes
Acknowledgment
This work was supported by Fresenius Medical Care GmbH.
Disclosure Statement
Ciro Tetta is a full time employee of Fresenius Medical Care GmbH.