
Abstract
We investigated whether human articular chondrocytes can be labeled efficiently and for long-term with a green fluorescent protein (GFP) lentivirus and whether the viral transduction would influence cell proliferation and tissue-forming capacity. The method was then applied to track goat articular chondrocytes after autologous implantation in cartilage defects. Expression of GFP in transduced chondrocytes was detected cytofluorimetrically and immunohistochemically. Chondrogenic capacity of chondrocytes was assessed by Safranin-O staining, immunostaining for type II collagen, and glycosaminoglycan content. Human articular chondrocytes were efficiently transduced with GFP lentivirus (73.4 ± 0.5% at passage 1) and maintained the expression of GFP up to 22 weeks of in vitro culture after transduction. Upon implantation in nude mice, 12 weeks after transduction, the percentage of labeled cells (73.6 ± 3.3%) was similar to the initial one. Importantly, viral transduction of chondrocytes did not affect the cell proliferation rate, chondrogenic differentiation, or tissue-forming capacity, either in vitro or in vivo. Goat articular chondrocytes were also efficiently transduced with GFP lentivirus (78.3 ± 3.2%) and maintained the expression of GFP in the reparative tissue after orthotopic implantation. This study demonstrates the feasibility of efficient and relatively long-term labeling of human chondrocytes for co-culture on integration studies, and indicates the potential of this stable labeling technique for tracking animal chondrocytes for in cartilage repair studies.
Introduction
Due to limited scientific evidence of the role of implanted chondrocytes in a cartilage defect and of their extent of participation in the repair process, chondrocyte labeling would also be extremely important before cell grafting in animal models. So far, only Dell'Accio et al. 11 used a fluorescent dye, PKH26, to label goat articular chondrocytes and showed that implanted cells persisted for at least 14 weeks in the defects and contributed to structural cartilage repair in a goat model of autologous chondrocyte implantation. This study represents an important milestone in the field, but the method used for labeling was not ideal for long-term studies because of the natural fading of PKH26 and the decrease in intensity of labeling with cell divisions.11,12
With the perspective of performing controlled in vitro or in vivo studies on the interaction between defined chondrocyte populations and other cell systems, a reliable technique for sustained chondrocyte labeling should be used. Besides membrane fluorochromes,11,13 other methods for cell tracking include the use of radioactive isotopes, 14 Y chromosome–specific probes, 15 or delivery of reporter genes. Among these, green fluorescent protein (GFP) is the most common and popular since it can be easily and efficiently viewed by blue fluorescent light excitation 16 or by the use of antibodies. Several methods for gene delivery have already been applied to chondrocytes for in vitro applications and in vivo purposes,17,18 such as plasmids transfection19,20 and infection with adenovirus,21,22 retrovirus,22,23 and baculovirus. 24 Among all viruses, lentiviral vectors based on human immunodeficiency virus offer many advantages, as they integrate to the genome enabling long-term gene expression and have the capacity to transduce both dividing and nondividing cells.25,26
Transduction of articular chondrocytes by lentivirus or retrovirus carrying GFP gene has previously been studied,22,23,27 but long-term gene expression and influence on chondrocyte phenotype have not been systematically described. Lu et al. 23 showed that transduction of rat chondrocytes with GFP lentivirus allowed to initially label about 60% of the cells, while after 6 weeks only 21% of chondrocytes remained labeled. A high efficiency of labeling of 85% on osteoarthritic chondrocytes was achieved and maintained over 15 weeks by Li et al., 22 but whether GFP lentivirus transduction affected chondrocyte redifferentiation was not described. Hirschmann investigated the efficiency of retroviral gene transfer into articular chondrocytes of rabbit, sheep, bovine, and human origin. 27 The expression of the GFP transgene remained stable for at least 4 weeks in vivo when transduced cells were implanted into cartilage defects in knee joints of rabbits. However, the influence of the marking procedure on matrix production was only investigated during chondrocyte expansion in monolayer.
Considering the limited density of cells in native cartilage and the typical phase of chondrocyte expansion before implantation or use in a variety of in vitro models, in this work we aimed at determining (i) the effect of GFP lentivirus transduction on human articular chondrocyte proliferation and postexpansion redifferentiation capacity, and (ii) the long-term in vitro or in vivo stability of GFP expression in engineered cartilaginous tissues generated using lentiviral-transduced human chondrocytes. The labeling of chondrocytes was followed by flow cytometry, fluorescence microscopy, or immunohistochemistry against GFP. Chondrogenic redifferentiation of transduced cells was assessed in different well-established models, including culture in micromass pellets28–30 or within hyaluronan-based scaffolds,31–33 as well as ectopic implantation in nude mice34–36 and systematically compared to the one of untransduced chondrocytes by histological, immunohistochemical, and biochemical analyses. Finally, to assess whether our labeling protocol could allow to track chondrocytes after orthotopic implantation in animal models, autologous GFP-transduced chondrocytes were also implanted in articular defects created in goats, a species frequently used for cartilage repair studies.11,37–39
Materials and Methods
Isolation and expansion of human articular chondrocytes
Biopsies of human articular cartilage were collected postmortem from knee joints of two female donors of 37 and 77 years old (respectively donor A and B), and from three male donors of 63, 70, and 76 years old (respectively donors C, D, and E) without macroscopic signs of arthritis, with the approval of the local ethics committee and informed consent by the relatives. Chondrocytes were isolated upon 22-h incubation at 37°C in 0.15% type II collagenase and resuspended in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin (complete medium). Chondrocytes were plated in tissue culture dishes at a density of 104 cells/cm2 and expanded in complete medium supplemented with 1 ng/mL transforming growth factor-β1 (TGF-β1) and 5 ng/mL fibroblast growth factor-2. The use of these growth factors was previously reported to increase chondrocyte proliferation rate and redifferentiation capacity, 28 and not to influence the transduction efficiency with lentivirus. 22 When cells were subconfluent, they were detached by sequential treatment with 0.3% type II collagenase and 0.05% trypsin/0.53 mM EDTA, and replated at 5 × 103 cells/cm2. Cell proliferation rate was defined as the total number of doublings during the expansion phase divided by the time required for the expansion, and was expressed as doublings/day. 40 In some experiments, cells were frozen in presence of 20% FBS and 10% DMSO.
Lentivirus GFP production and titration
The plasmids pMD.2G (Addgene plasmid 12259), pCMVΔR8.91, 41 and pWPXL-EGFP (Addgene plasmid 12257) were kindly provided by D. Trono (Geneva, CH, Switzerland). High-titer GFP lentiviral supernatants were generated by transient cotransfection of three plasmids in 293T cells using Fugene 6 (Roche, CH, Switzerland). About 293T cells (2.5 × 106 cells) were transfected with 12 μg of pWPXL-EGFP, 12 μg of pCMVΔR8.91, and 5 μg of pMD.2G. Supernatants of transfected 293T cells were collected 2 and 3 days posttransfection, filtered through 0.45 μm pore size filters, and stored at −70°C. The viral titer was determined on HeLa cells infected with serial dilutions of each viral supernatant in presence of protamine sulfate (5 μg/mL; Sigma, St. Louis, MO) by flow cytometry analysis. The titer was calculated according to the following formula: Transduction Units (TU)/mL = (Percentage of GFP-positive cells/100) × (Number of cells infected) × (Vector dilution factor), as described previously. 42 Viral titer typically ranged around 3.7 × 106 TU/mL after 2 days and 2.0 ×106 TU/mL after 3 days. The cell lines 293T cells and HeLa cells were maintained in Dulbecco's modified Eagle's medium containing 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin.
Transduction of human articular chondrocytes
Freshly isolated articular chondrocytes (0.56 × 106 cells) were seeded in 56 cm2 dishes and transduced the following day with 2.8 × 106 or 5.6 × 106 TU of GFP lentivirus (multiplicity of infection, MOI, of 5 or 10) in presence of protamine sulfate 5 μg/mL final. The percentage of GFP-positive cells was assessed by flow cytometry, 3 days posttransduction to determine the efficiency of transduction, and after the first and second passages in culture, to follow the percentage of GFP-expressing cells during expansion. Data acquisition and analysis were performed using CellQuestPro software (Becton Dickinson, Franklin Lakes, NJ). Transduced chondrocytes were expanded and cultured in pellets and 3D scaffolds as described below, in parallel with untransduced cells, used as controls. For cytofluorimetry, cells were retrieved from pellets by sequential treatment with 0.3% type II collagenase and 0.05% trypsin/0.53 mM EDTA.
Culture of human articular chondrocytes in micromass pellets
Second-passage cells, in aliquots of 5 × 105, were centrifuged in polypropylene conical tubes (Sarstedt, Numbrecht, Germany) to form spherical pellets, which were cultured for 2 weeks in 0.5 mL of a chemically defined serum-free medium containing 10 ng/mL TGF-β1 and 10−7 M dexamethasone, as previously described. 28
Culture of human articular chondrocytes on porous 3D scaffolds
HYAFF®11 nonwoven meshes made of esterified hyaluronic acid (Fidia Advanced Biopolymers, Abano Terme, Italy) were cored as discs (6 mm diameter, 1.5 mm thick), and placed on dishes coated with a thin film of 1% agarose. Second-passage chondrocytes (5 × 106/scaffold) were distributed on the top surface. Constructs were statically cultured from 2 to 20 weeks in complete medium supplemented with 10 μg/mL insulin, 0.1 mM ascorbic acid 2-phosphate, and 10 ng/mL TGF-β3, with culture medium replaced twice a week. These supplements were previously shown to enhance chondrogenesis of dedifferentiated human chondrocytes during culture into HYAFF11 nonwoven meshes. 35 Constructs (n = 4 per condition and time point) were either precultured for 2 weeks and further implanted in nude mice as described below or were harvested at the time points 2, 4, 8, 12, 16, and 20 weeks, bisected, and analyzed by histology, immunohistochemistry, biochemistry, and fluorescence microscopy.
Ectopic implantation of human articular chondrocyte constructs
Human articular chondrocyte–HYAFF11® constructs cultured for 2 weeks (with control or transduced cells) were implanted in the back of two nude mice (CD-1 nu/nu, athymic, 6–8-week-old females) in a pocket between excised muscle fascia and subcutaneous tissue. All animals in this study were cared for and treated according to institutional guidelines. Each mouse received four grafts, two generated by control cells and two generated by transduced cells. Constructs were harvested after 8 weeks and processed for histological and immunohistochemical analyses and fluorescence microscopy.
Generation and orthotopic implantation of autologous goat articular chondrocytes constructs
Surgical procedures were performed following protocol approval by the cantonal veterinary office (Amt für Landwirtschaft des Kantons Bern) using sterile techniques and under general anesthesia. Anaesthetic induction was obtained by intramuscular injection of xylazin (Vetoquinol, Ittigen, Switzerland), followed by intravenous application of midazolam (Roche Pharma, Basel, Switzerland), and the intraoperative anesthesia was maintained by isofluoran inhalation. Postoperative antibiotic (Amoxycillin; GlaxoSmithKline AG, Münchenbuchsee, Switzerland) and analgetic (Flunixin; Biochema, Uzwil, Switzerland) therapy was applied for another 3 days. In a first surgery, both posterior legs of two skeletally adult female Saanen goats (older than 18 months) were laterally operated. To harvest the biopsies, four circular defects of 6 mm in diameter and 0.4–0.5 mm in depth were created in the cartilage of the trochlea groove using a disposable biopsy punch (Stiefel, Offenbach am Main, Germany). Chondrocytes isolated from the excised cartilage were transduced at MOI of five using the protocols previously described for human chondrocytes. Transduced chondrocytes were then expanded for two passages in complete medium supplemented with 5 ng/mL fibroblast growth factor-2. 43 The percentage of GFP-positive cells was quantified cytofluorimetrically at the end of the second expansion as previously described. Collagen type I/III membranes (Chondrogide®; Geistlich, Wolhusen, Switzerland) were cored as discs (6 mm diameter), and cells were statically seeded at a density of 2 × 106 cells/scaffold on the porous side of the dry membrane, the smooth side being rather impermeable to cells. Constructs were then cultured for 2 weeks in a complete medium supplemented with 10 μg/mL insulin, 0.1 mM ascorbic acid 2-phosphate, and 10 ng/mL TGF-β3, with the culture medium replaced twice a week. Four weeks after biopsy withdrawal, in a second surgical intervention the biopsy sites in the trochlea femoris were re-cored to a final size of 6 mm diameter × 2 mm thickness and filled with the in vitro precultured constructs, which were sutured to the surrounding cartilage with four stitches. Goats were sacrificed 4 weeks postoperatively by injection of an overdose of phenobarbital, and samples were harvested for the immunohistochemical detection of GFP-labeled chondrocytes.
Histological and immunohistochemical analyses
Cell pellets, engineered tissues, ectopic implants, or reparative tissues were fixed in 4% formalin, embedded in paraffin, cross sectioned (5 μm thick for pellets and 7 μm thick for engineered tissues and implants), and stained with Safranin-O for sulfated glycosaminoglycans (GAG). In some experiments, pellets or constructs were cryo-embedded; 7-μm-thick sections were assessed for the expression of GFP by direct observation using a fluorescent microscope (Axiophot; Zeiss, Jena, Germany), using transmitted light images of the same microscopic field to view all cells.
For immunohistochemical assessment of GFP expression, sections were deparaffinized and incubated with 0.1% trypsin for 20 min at 37°C and afterward with rabbit serum (dilution 1:20) for 10 min. For detection of GFP in tissues generated using human chondrocytes, sections were incubated for 1 h with a goat polyclonal antibody against GFP (dilution 1:200) (ab6673; Abcam, Cambridge, United Kingdom), followed by a secondary antibody, rabbit anti-goat biotinylated (dilution 1:400) (E0466; Dako, Carpenteria, CA). For detection of GFP in tissues generated using goat chondrocytes, sections were incubated for 1 h with a rabbit polyclonal antibody against GFP (dilution 1:200) (ab290; Abcam), followed by a secondary antibody, horse anti-rabbit biotinylated (dilution 1:100) (BA-1400; Vector, Burlingame, CA). The avidin–biotin complex coupled with alkaline phosphatase (ABC/AP complex; Dako) was used for the staining according to manufacturer's instructions followed by incubation with Fast Red substrate and Levamisole (Dako). The percentage of GFP-positive chondrocytes was derived by manual counting of the number of cells positively stained by Fast Red and of the number of total cells, which were viewed by counterstaining with hematoxylin. The implants were surrounded by a thin layer of fibroblastic cells, which was not considered for analysis, since most likely formed by mice cells. Four microscopic fields for each cross section, corresponding to an average of 323 ± 35 cells/field, were assessed.
For collagen type II immunohistochemistry, sections were processed using an antibody against type II collagen (II-II6B3, Hybridoma Bank, University of Iowa) as previously described. 43
Biochemical analysis
Cell pellets and engineered tissues were digested with proteinase K solution as previously described. 44 GAG content was measured spectrophotometrically after reaction with dimethylmethylene blue dye, 45 with chondroitin sulfate as a standard. The GAG content was normalized to the DNA amount, which was measured using a CyQUANT cell proliferation assay kit (Molecular Probes, Eugene, OR), with calf thymus DNA as a standard.
Results and Discussion
Lentiviral transduction of chondrocytes
Human articular chondrocytes (donor B) were transduced with the recombinant GFP lentivirus at MOI of 5 and 10. In a preliminary experiment, the percentage of chondrocytes positive for GFP as analysed by flow cytometry 6 days posttransduction was not significantly higher when the transduction was performed at MOI 10 as compared to MOI 5 (75.6% vs. 75.0%). For this reason, further experiments were all conducted at MOI 5.
Three days posttransduction with GFP lentivirus, labeled chondrocytes could be detected by fluorescence microscopy (Fig. 1A, B). The percentage of GFP-positive cells 3 days posttransduction was 71.1 ± 2.6% as determined by cytofluorimetry with cells from donors B, C, D, and E. At passage 2, the fraction of labeled cells could be quantified by a clear positive peak by cytofluorimetry (Fig. 1E). The percentage of GFP-positive cells after the first passage (corresponding to, respectively, 2.8 and 3.9 doublings for cells from donors A and B) was 73.6 ± 1.4%, and remained essentially stable (73.4 ± 0.5%) after the second passage (corresponding to, respectively, 7.2 and 7.5 doublings for cells from donors A and B).

Representative pictures of expanded chondrocytes untransduced (control) (
Cell proliferation rate and redifferentiation capacity in pellets
The proliferation rate of chondrocytes, assessed until the second passage of expansion, was only slightly variable using cells from the two donors, and was not significantly different when cells were untreated (control) or transduced with GFP lentivirus (Fig. 2). These results again confirm the limited intradonor variability in the response of chondrocytes to GFP transduction.

Cell proliferation rate (as doublings/day) of chondrocytes isolated from cartilage biopsies of the two donors, transduced with GFP lentivirus or untransduced (control) and expanded for two passages.
The redifferentiation capacity of expanded chondrocytes (donor A) was assessed in 3D micromass pellet cultures. The intensity of staining for sulfated GAG (Safranin-O staining, Fig. 3A) was similar in pellets generated from control chondrocytes as compared to GFP-transduced chondrocytes. The GAG/DNA ratios were not significantly different in pellets generated from control chondrocytes as compared to GFP-transduced chondrocytes (Fig. 3A), consistently with the Safranin-O staining. Observation of pellet cryosections under fluorescent light demonstrated a positive signal when chondrocytes were transduced with the GFP lentivirus (Fig. 3B). Our results are generally in agreement with previous studies indicating that the expression of GFP protein does not influence the chondrogenic differentiation of murine mesenchymal progenitor cells.46,47
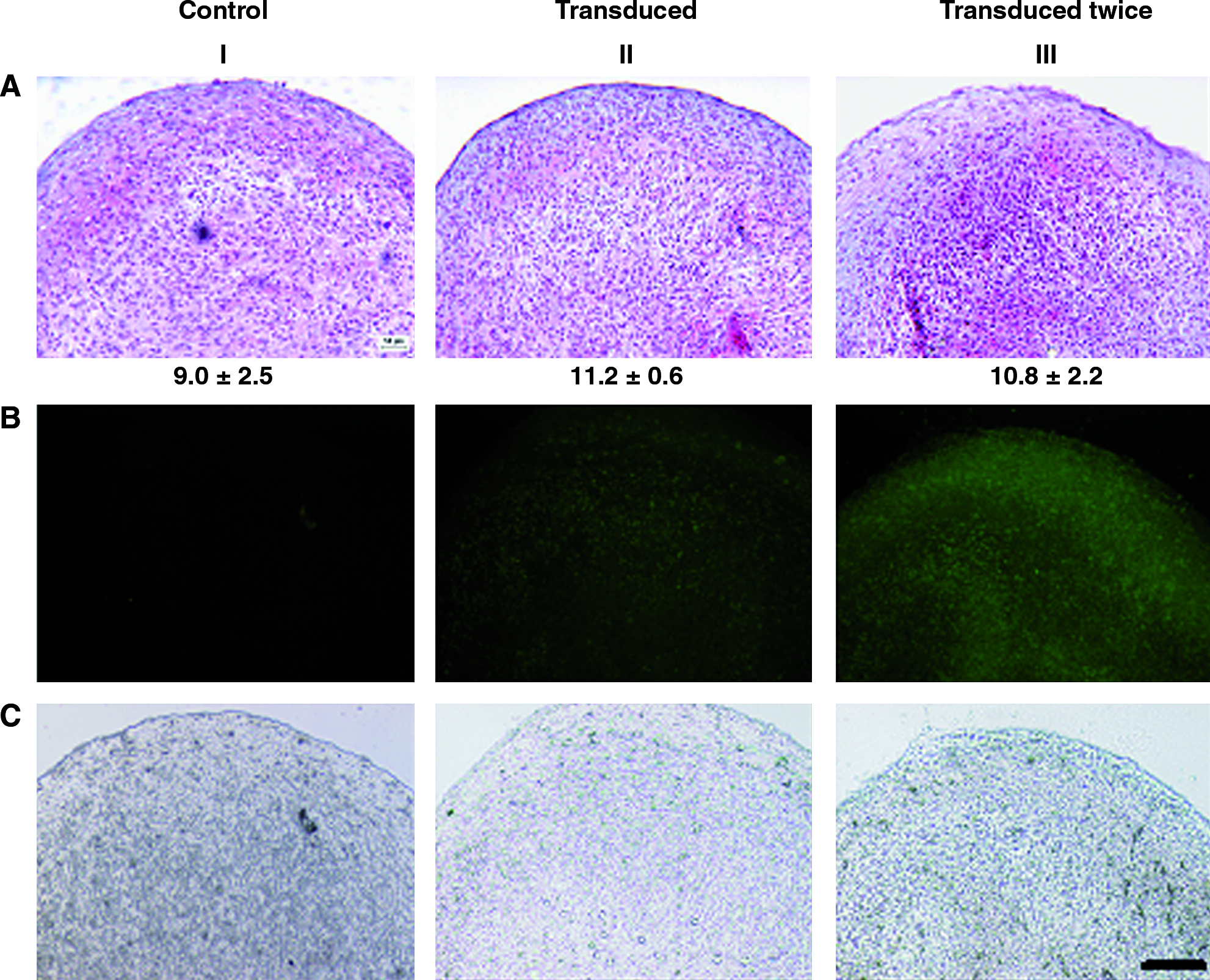
Chondrocyte redifferentiation in 3D pellets. Pellets generated from untransduced (control) (I), transduced (II), or twice transduced (III) chondrocytes from donor A were stained by Safranin-O (
We next investigated the influence of cell freezing after GFP transduction on the percentage of labeled chondrocytes and on their redifferentiation capacity in pellets. The introduction of a cell freezing step did not modify the percentage of isolated fluorescent chondrocytes, as quantified by cytofluorimetry (73.1% positive cells after the freezing step vs. 73.6% before cell freezing). Our findings are in line with those by Li et al., 22 who reported that the expression of GFP in transduced osteoarthritic chondrocytes was maintained after freezing and re-culture.
Pellets generated from transduced chondrocytes with or without freezing exhibited similar intensity of staining for sulfated GAG (data not shown) and GAG/DNA ratios (10.5 ± 0.1 with freezing step vs. 11.2 ± 0.6 without freezing step). After 2 weeks in pellets, the fluorescence intensity of transduced chondrocytes was comparable if a freezing step was performed or not during previous cell expansion (data not shown).
To possibly increase the percentage of GFP-labeled cells, a second cycle of transduction was performed on already transduced chondrocytes. This second cycle of transduction increased the percentage of fluorescent chondrocytes from 73.6% to 85.6 ± 1.5% at passage 1 as detected by cytofluorimetry. This is consistent with Li et al., 22 reporting that after two cycles of retroviral transduction the percentage of labeled human chondrocytes increased from 85% to 96%. Pellets generated from double-transduced chondrocytes exhibited a similar intensity of staining for sulfated GAG as well as similar GAG/DNA ratios as compared to those generated from cells transduced only once (Fig. 3A). Observation of pellet cryosections under fluorescent light qualitatively indicated a more homogenous green staining in pellets generated from double-transduced chondrocytes as compared to cells transduced only once (Fig. 3B). The percentages of single- or double-transduced chondrocytes enzymatically retrieved from pellets after 2 weeks culture and still positive for GFP averaged, respectively, 62.5 ± 2.3% or 75.9 ± 4.6%, as assessed by cytofluorimetry.
Cell redifferentiation capacity in 3D scaffolds
As an independent model of chondrogenic differentiation and tissue formation, untransduced (control) or transduced chondrocytes (donor A), after 2 weeks of expansion, were cultured in HYAFF®11 scaffolds for up to additional 20 weeks. Engineered cartilaginous tissues continuously matured in vitro until 8 weeks, as indicated by Safranin-O staining, but longer culture time led to progressive degradation of the tissue, as typically occurs under static culture conditions. 48 After 4 weeks of culture, generated constructs displayed faint staining for Safranin-O, which was of similar intensity for control and transduced chondrocytes (Fig. 4A). Immunohistochemistry for GFP protein indicated a strong positive staining in constructs generated from transduced cells (Fig. 4B), in correspondence both of elongated cells at the construct periphery and of more rounded cells in the central part of the tissue. After 8 weeks or longer culture time, cells localized at the tissue periphery were still predominantly positively stained for GFP, whereas the construct core was essentially necrotic, not allowing reliable assessment of positivity for GFP staining (data not shown).

Analysis of constructs cultured for 4 weeks and generated from untransduced (control) (I) or transduced chondrocytes (II) from donor A. Cross sections were stained by Safranin-O (
By fluorescence analysis of cryosections, HYAFF®11 fibers appeared unspecifically fluorescent in constructs generated from control or transduced chondrocytes (Fig. 4C). The same microscopic fields were thus observed also in light microscopy to distinguish fluorescent HYAFF®11 fibers from fluorescent cells (Fig. 4D). This analysis allowed to confirm that only constructs generated using transduced chondrocytes contained GFP-positive cells.
Cell redifferentiation capacity and tracking after ectopic implantation in nude mice
Expanded, transduced, or untransduced chondrocytes (donor A) were cultured in HYAFF®11 scaffolds for 2 weeks and further implanted subcutaneously in nude mice for 8 weeks. The chondrogenic redifferentiation capacity was assessed via staining for sulfated GAG, which appeared to be of strong and similar intensity in implants generated using untransduced or transduced chondrocytes (Fig. 5A). A similar intensity of immunostaining for collagen type II was observed in implants generated from control or transduced chondrocytes (Fig. 5B). Collagen type II positive staining was detected in the same regions stained for sulfated GAG. GFP expression was demonstrated by immunohistochemistry in implants generated using transduced chondrocytes (Fig. 5C), indicating that in vivo cells were still labeled and detectable. Manual counting of cells positively stained for GFP and of all cells with hematoxylin-stained nuclei was performed on histological sections of implants generated using transduced chondrocytes. Such counting demonstrated that 73.6 ± 3.3% of the cells in the implants were positive for GFP after 12 weeks of transduction, corresponding to a complete maintenance of labeling by the transduced cells. Fluorescence analysis of cryosections of the implants confirmed the presence of GFP-expressing chondrocytes (data not shown).
Analysis of constructs implanted in nude mice for 8 weeks and generated from untransduced (control) (I) or transduced chondrocytes (II) from donor A. Cross sections were stained by Safranin-O (
Transduced chondrocytes implanted in nude mice did not show an apparent reduction in the number of positive cells after 12 weeks, while transduced chondrocytes cultured for 2 weeks in 3D pellets exhibited around 10% reduction of GFP positivity. In the two experimental designs, we used different culture systems (scaffolds vs. 3D pellets) as well as distinct methodologies to determine the number of labelled cells (direct counting after immunohistochemistry vs. cytofluorimetry after enzymatic digestion of pellets). The methodologies employed in this study could explain relative variations observed in the number of GFP-positive chondrocytes.
Cell tracking after orthotopic implantation in a goat model
The percentage of GFP-positive goat articular chondrocytes after the second passage was slightly higher after a single transduction than that measured in human articular chondrocyte cultures, averaging 78.3 ± 3.2% as measured by cytofluorimetry. This percentage is in the same range of that reported by Ueblacker et al. 49 and indicates that the protocol used for labeling human chondrocytes is efficient also for the transduction of chondrocytes from other species.
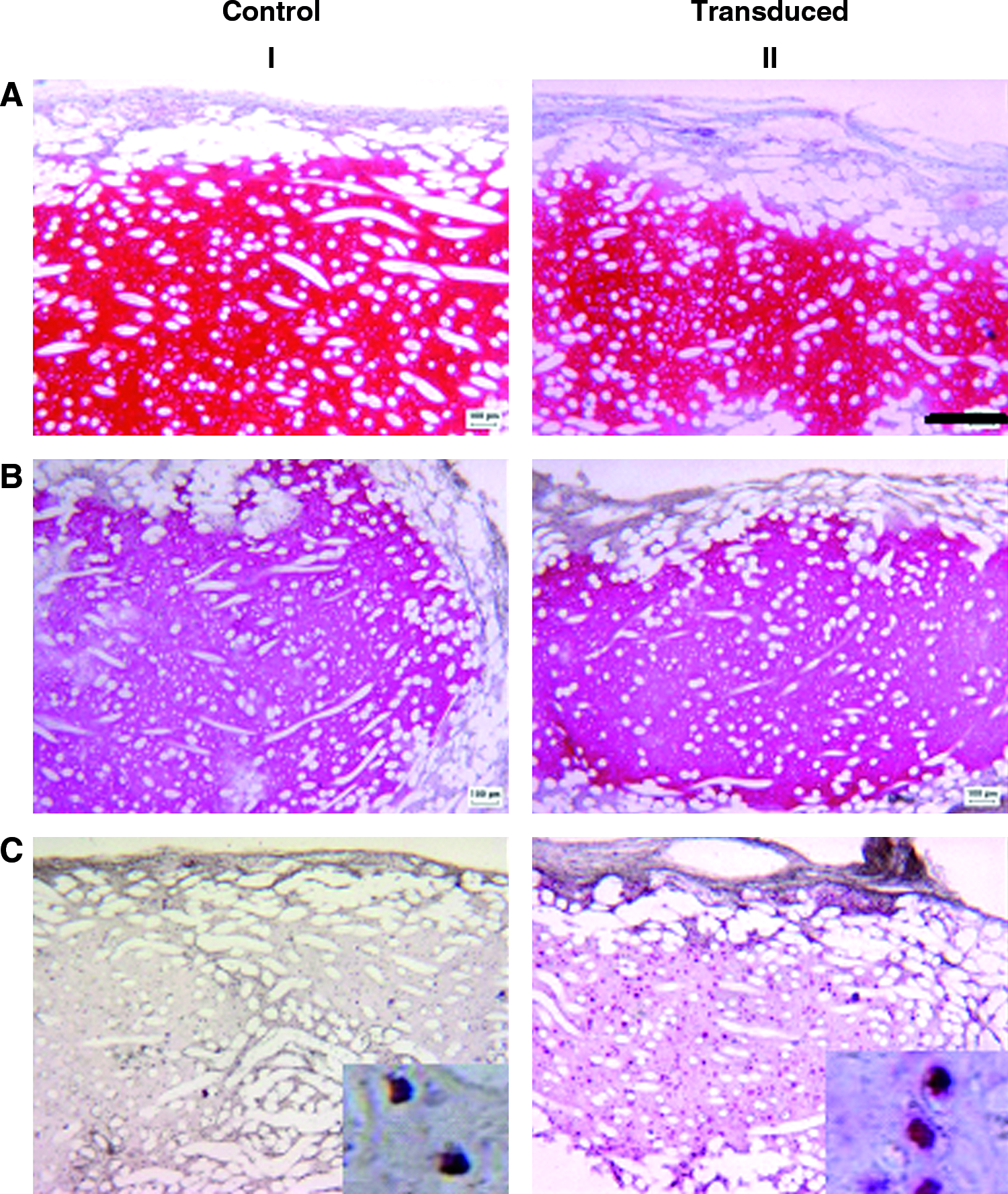
GFP-labeled chondrocytes were cultured in Chondrogide membranes for 2 weeks and then implanted into defects generated in the trochlea of goats (Fig. 6A). After 4 weeks, visual observations showed an apparent good filling of the defect with a relatively smooth surface (Fig. 6B). No inflammatory reactions, possibly related to scaffold degradation products, or blood vessel ingrowth were observed in the repair tissue. Immunohistochemical analysis demonstrated a large amount of GFP-positive cells (Fig. 6C, D) in the reparative tissue, indicating that the grafted chondrocytes remained viable in the defect. Quantification of GFP-positive cells from these sections (as percentage of total) appeared not reliable, due to the possible infiltration of cells from other compartments (e.g., the subchondral bone or synovial membrane) in the grafted material, which could not be distinguished from nonlabeled implanted cells. Although in our study only two goats have been used and a relative short follow-up time was considered, the results represent a proof of principle that the proposed GFP transduction method would enable to track implanted chondrocytes in orthotopic cartilage repair studies and possibly to evaluate the contribution of grafted cells in the repair process.

Analysis of the constructs implanted in goat cartilage defects for 4 weeks. (
Conclusions
We report that primary human articular chondrocytes can be transduced by a GFP lentiviral vector at high efficiency, with labeled cells detected up to 22 weeks after transduction using in vitro 3D cultures, and up to 8 or 12 weeks in in vivo models, namely, orthotopic implantation in goats and subcutaneous implantation in nude mice. Importantly, chondrocyte transduction did not affect the cell proliferation rate or the postexpansion redifferentiation capacity. The developed and validated technique could be used as a tool to engineer labeled cartilage tissues, and investigate the interaction between chondrocytes and other cell populations during crucial processes (e.g., migration, differentiation, and integration) in in vitro systems or immunodeficient animal models. We additionally proved that this method could be extended to the transduction of chondrocytes from other species, thus allowing evaluating the fate of implanted cells in orthotopic cartilage repair studies in large animal models.
Footnotes
Acknowledgments
We would like to thank Fidia Advanced Biopolymers for supplying HYAFF®11 nonwoven meshes, Geistlich for supplying Chondrogide membranes, Prof. D. Trono (Geneva) for kindly providing plasmids for the generation of GFP lentivirus, and Ms. Francine Wolf for her technical support with the immunohistochemical analyses.
Disclosure Statement
No competing financial interests exist.