
Abstract
We have developed an efficient decellularization process for the isolation of extracellular matrix (ECM) from native cardiac tissue. The isolated ECM exhibited desirable mechanical properties in terms of elasticity, strength and durability–properties required from scaffolds used for cardiac tissue repair. This study further investigates the potential use of this scaffold for cardiac tissue engineering in terms of interactions with seeded cells and biocompatibility. We used the commonly studied fibroblasts, cardiomyocytes, and mesenchymal stem cells, which were isolated and seeded onto the scaffold. Cell density and distribution were followed by 3,3'-dioctadecyloxacarbocyanine perchlorate staining, and their proliferation and viability were assessed by AlamarBlue® assay and fluorecein-diacetate/propidium iodide staining. Fibroblast-seeded scaffolds shrank to 1–2 mm3 spheroids, and their glycosaminoglycans significantly increased by 23%. The expression of ECM remodeling-related mRNAs of collagens I and III, matrix metalloproteinase 2, and type 1 tissue inhibitor of metalloproteinases was quantified by real-time polymerase chain reaction, and was found significantly elevated in fibroblast-seeded scaffold, compared with the control cells on plates. Fibroblast-seeded scaffolds lost some flexibility, yet gained strength compared with the acellular scaffolds, as shown by mechanical testing. Scaffold seeded with cardiomyocyte began to beat in concert few days after seeding, and the myocytes expressed typical functional cardiac markers such as α-actinin, troponin I, and connexin43. The cells revealed aligned elongated morphology, as presented by immunofluorescent staining and scanning electron microscopy. Mesenchymal stem cell–seeded scaffolds maintained viability over 24 days in culture. These findings further strengthen the potential use of acellular cardiac ECM as a biomaterial for heart regeneration.
Introduction
In recent years there has been an increasing interest in natural acellular extracellular matrix (ECM) for tissue engineering application of various organs.28,29 ECM from various sources has been rendered acellular by implementation of different decellularization techniques for tissue engineering of heart valves, blood vessels, urinary bladder, and more.29–31 Acellular urinary bladder ECM has also been studied for the regeneration of infracted hearts.32–36 However, ECM isolated from different tissues and organs possess different properties in terms of collagen type, content and density, glycosaminoglycans (GAG) composition, and susceptibility to different decellularization procedures. 37 Recently, cardiac-derived ECM has been reported for the engineering of heart tissue. 38 The cardiac ECM was isolated from porcine and rat heart, and the feasibility of the construct was demonstrated by seeding neonatal rat cardiomyocytes on ECM isolated from the rat heart.
In this study, we describe a new and efficient decellularization process for cardiac tissue, which allows the complete removal of the cellular component while maintaining the mechanical and biochemical properties of the isolated cardiac ECM. The cardiac ECM was obtained by decellularization of porcine myocardium slices, using a combined enzymatic and chemical process, developed and optimized for this purpose in our laboratory (Patent Cooperation Treaty patent). To study the potential of such isolated ECM patch for cardiac tissue reconstruction, we have addressed the ECM's intrinsic biochemical and mechanical properties. The biological functionality of the isolated ECM was evaluated using adherence, proliferation, and long-term viability studies of seeded fibroblasts, cardiomyocytes, and MSCs. We have also evaluated the promotion of normal cell phenotype, functionality, and the ability of populating cells to remodel the scaffold, as well as the potential immunogenecity of such scaffold.
Materials and Methods
Scaffold preparation: Decellularization of porcine heart tissue
Hearts of commercial healthy slaughter-weight female pigs were harvested and immersed in cold, sterile phosphate-buffered saline (PBS), supplemented with antibacterial and antimycotic antibiotics (200 U/mL penicillin, 0.2 mg/mL streptomycin [Pen-Strep]; Biological Industries, Beit Haemek, Israel) and 2 μg/mL amphotericin B (Fungizone®; Gibco, Long Island, NY). The atria, right ventricle, and any excess fat were excised from the left ventricle, which was horizontally cut into three. The two upper rings were cut into three or four (depending on the ring diameter), and each part was sliced parallel to the epicardial plain into 3 mm-thick tissue slices. The slices were washed in hypertonic 1.1% NaCl–0.02% ethylenediaminetetraacetic acid (EDTA; Sigma, St. Louis, MO) for 2 h and then in hypotonic 0.7% NaCl–0.02% EDTA for 2 h. Tissue slices were subjected to two 24 h cycles of enzymatic digestion using 0.05% trypsin (Sigma)–0.02% EDTA in PBS at pH 7.4 and at 37°C, supplemented with Pen-Strep and Fungizone. Tissue slices were then agitated in a fresh detergent solution of 1% Triton®-X-100 (polyethylene octyl phenyl ether) and 0.1% ammonium hydroxide in PBS for four consecutive 48 h cycles. The acellular matrices were washed extensively in sterile saline, immersed in 70% ethanol overnight, washed in sterile water, and lyophilized.
Histological staining was performed to verify complete acellularity of the ECM scaffolds. Fresh cardiac tissue and decellularized ECM were frozen in Tissue-Tek® optimal cutting temperature (OCT) compound (Sakura Finetek, Torrance, CA), cross sectioned to 10 μm, and stained to detect any intact cell nuclei, membrane residues, and cytoskeletal actin using (4′,6-diamidino-2-phenylindole [DAPI], 0.1 μg/mL; Sigma), and phalloidin–fluorescein isothiocyanate (FITC) (0.1 μg/mL; Sigma) in PBS. Stained sections were washed twice in PBS and viewed by fluorescent microscopy (Nikon Eclipse TE2000-s with a DXM1200 camera, Nikon instruments, Melville, NY). Representative images were obtained for the entire sample widths.
Immunofluorescent staining of collagen I, collagen III, and fibronectin
Acellular ECM samples were frozen in optimal cutting temprature (OCT), sectioned to 10 μm sections, and fixed in cold methanol. The sections were incubated for 1 h at room temperature (RT) with either primary mouse anti-collagen I or anti-collagen III antibody (1:100; Sigma). Samples were reincubated with a secondary Cy3-conjugated goat anti-mouse antibody (1:250; Jackson Immuno Research Laboratories, Inc., West Grove, PA), washed in PBS, and incubated with a FITC-conjugated goat anti-fibronectin F (ab′) 2 fragment (1:200; Cappel-MP Biomedicals, Solon, OH). Control slides were incubated only with the secondary antibody under similar conditions. Slides were washed in PBS, mounted with Fluoromount-G (Southern Biotech, Birmingham, AL), and imaged by inverted fluorescent microscope (Nikon Eclipse TE2000-s with a DXM1200 camera). Digital deconvolution was performed on the images using the imageJ software. Relative picture data analysis was performed using the NIS elements ver. 3.0 software (Nikon Instruments, Melville, NY).
Primary cell isolation and culture
Cardiac fibroblasts were isolated from a healthy adult (6 years old) sheep myocardium. Tissue samples from the left ventricle wall were diced into small pieces of approximately 1 mm3, washed in sterile PBS, and placed in the culture plates. Culture medium—Dulbecco's modified Eagle's medium high (Sigma), 10% fetal calf serum (Biological Industries), 100 U/mL penicillin, 0.1 mg/mL streptomycin, and 1 μg/mL Fungizone—was slowly added after 10–12 min to the plates. The seeded plates were incubated in a humidified 37°C, 8% CO2 incubator, until fibroblasts outgrew from the tissue pieces. Cells from passages 4 and 5 were used for different experiments.
Cardiac myocytes were isolated from 1- to 2-day-old Sprague-Dawley rats. Excised hearts were minced, and the cardiac cells were dissociated by gentle agitation in 200 U/mL collagenase type II (Worthington Biochemical cooperation, Lakewood, NJ) in PBS-G (PBS, Pen-Strep, 0.1% glucose) at 37°C for 10 min. Cell suspensions were centrifuged at 1000 rpm for 5 min, suspended in culture medium (same as for fibroblasts), and filtered through a 100 μm pore sieve to remove clusters. Cell suspension was preplated in culture dishes and incubated for 1 h to allow adherence of fibroblasts. Nonattached myocyte-enriched cell suspension was collected, centrifuged as before, and resuspended in F-10 nutrient mixture supplemented with 5% fetal calf serum, 5% donor horse serum (Biological Industries), Pen-Strep, Fungizone, and 1 mM CaCl2. Cardiomyocytes were cultured in a humidified 37°C, 8% CO2 incubator. Proliferation of fibroblasts was eliminated by the addition of 25 μg/mL bromo-deoxy uridine (BrdU; Sigma) to the culture medium during the first days of culture, as previously described.39,40
Bone marrow MSCs were isolated from the femurs and tibiae of 3-month-old Sprague-Dawley rats. Marrow was flushed out of the bones with standard culture medium using a syringe and 18-gauge needle. Marrow clusters were dispersed by gentle pipetting, and the cell suspension was centrifuged (1000 rpm, 5 min) and resuspended in the culture medium. Cells from each bone were separately seeded in the culture plates and cultured in a humidified 37°C, 5% CO2 incubator. Nonadherent cells were removed by several washes with fresh medium, 1 day after seeding. MSCs were cultured in standard medium (Dulbecco's modified Eagle's medium low; Biological Industries), and at 70–80% confluence, the cells were harvested and replated (P1) for an additional week. Cells were analyzed for surface cell markers (CD90+CD44+CD29+ and CD34−) by FACSCalibur™ (BD (Biosciences, San Jose, CA) unpublished data).
Culture, distribution, and viability of primary cells seeded onto the acellular cardiac scaffolds
Lyophilized scaffolds were rehydrated in culture medium for 24 h before seeding. Cardiac fibroblasts, cardiomyocytes, or MSCs (5 × 105 cells) were pipetted slowly directly onto the 10 × 10 mm scaffolds, and 1 h later the culture medium was carefully added to the plates. Culture medium was changed every other day.
To follow cell distribution on seeded scaffolds, cells were stained with DiO (5 μg/mL in culture medium for 2 h; Molecular Probes) before seeding onto the scaffold. Seeded scaffolds were imaged at different time points by inverted fluorescent microscopy. The seeded surface was kept upward, and so the translucence of the scaffold enabled to view the cells on its surface as well as in its subsurface. To view the morphology of the cytoskeleton, actin filaments were stained with FITC-conjugated phalloidin and imaged by fluorescent microscopy.
The viability of the seeded fibroblast and myocyte cells was evaluated using the noncytotoxic AlamarBlue (Serotec, AbD Serotec, Kidlington, United Kingdom) according to the manufacturer's recommendations (n = 12 for each cell type). Preliminary experiments were carried out to establish optimal volume and incubation time for each cell (data not shown). The seeded scaffolds were incubated with the AlamarBlue reagent at different time points in the experiment, after which they were washed in fresh medium and recultured. Scaffolds seeded with fibroblasts and MSCs were moved to new culture plates before each viability measurement, to avoid deviations due to cells, which had migrated from the scaffold and adhered to the plate surface. As control, cells were seeded onto plastic 12-well plates, and their viability was measured at the same time points. The viability values presented here are a generalized combination of the proliferation, vitality, and mortality of cells at each time point.
To further qualitatively assess cell viability, scaffolds seeded with fibroblasts and MSCs were subjected to a fluorecein-diacetate (FDA)/propidium iodide (PI) staining assay. Seeded scaffolds were immersed for 10 min in 0.5 μg/mL FDA (Fluka) and 1 μg/mL PI (BD Biosciences) in PBS at RT in the dark. Samples were washed twice in PBS and imaged by fluorescent microscopy.
Immunophenotyping of cardiomyocytes on scaffolds
Scaffolds seeded with cardiomyocytes were fixed in 4% paraformaldehyde, washed with PBS, blocked, and incubated with primary mouse anti-α-actinin, anti-connexin 43 or anti-troponin I (1:250, 1 h at RT; Chemicon). Cy3-conjugated goat anti-mouse served as secondary antibody (1:500, 1 h at RT; Jackson). Control slides were incubated with only the secondary antibody. Confocal images were captured using a (Bio-Rad, Hercules, CA) Radiance 2000 equipped with a Nikon E600 upright microscope, at Z-plane depth of 50 μm.
Scanning electron microscopy
Decellularized and native matrices were washed in distilled water and snap-frozen in liquid nitrogen. To observe the acellular matrix, samples were fractured while frozen and were then lyophilized overnight. Seeded ECM samples were washed in PBS, fixed overnight in 0.1% formalin and 2 h in 2.5% glutaraldehyde, washed extensively in PBS, dehydrated in ascending concentrations of ethanol, and air dried. Samples were gold sputtered and mounted for imaging on a JSM-5400 scanning electron microscope (Jeol, Tokyo, Japan).
QuantomiX® WET-SEM™
Scaffolds (10 × 10 mm) seeded with 5 × 105 cardiac fibroblasts were cultured for 28 days as described above, washed in saline, stained with 1% tannic acid and 0.1% uranyl acetate, mounted on a QX-302 tissue capsule, and imaged at QuantomiX Laboratories (Nes-Ziona, Israel;
Collagen and GAG quantification
Total collagen was quantified in seeded matrices at different time points (1, 5, 12, 19, and 24 days after seeding, n = 6 for each time point) using the hydroxyproline assay as described previously. 41 GAG contents were quantified in seeded and nonseeded matrices at the same time points using the safranin O method as described previously (n = 6 for each time point). 42
Expression of ECM remodeling-related genes
The expression of collagen I (α1 chain), collagen III (α1 chain), matrix metalloproteinase 2 (MMP2), and type 1 tissue inhibitor of metalloproteinases (TIMP1) by fibroblasts seeded onto the scaffolds with time was quantitatively studied by real-time polymerase chain reaction (PCR). Fibroblasts were seeded onto the ECM (n = 4) as described above. Total RNA was isolated from the seeded cells at different time points using Tri-reagent (Sigma) following manufacturer's instructions, and 0.5 μg RNA was used for reverse transcription (first strand reverse-iT; ABgene, Epsom, United Kingdom). Primers were designed for sheep to specifically amplify genes' cDNA as follows (5′ to 3′): CCAGTCACCTGCGTACAGAA and GCACGGAAATTCCTGTTGAT for collagen Iα1; CACAAGGAGTCTGCATGTCT and GTTCACCAGGCTCACCAGCA for collagen IIIα1; CTGATGGCGCCCATTTATAC and GATGAACCGGTCCTTGAAGA for MMP2; CCACCCCACCCACAGACG and CGCAGGGACTGCCAGGTG for TIMP1; and ATGCTGGTGCTGAGTACGTG and ATCCACAGTCTTCTGGGTGG for glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as intrinsic housekeeping gene control. Reactions were prepared with Power SYBER® Green PCR Master Mix, run on ABI 7300, and analyzed using the 7000 SDS software (Applied Biosystems, Foster City, CA).
Mechanical properties of seeded and unseeded scaffolds
To study the effect of seeded cells on the scaffold tensile properties, acellular scaffolds (∼30 × 20 mm) were seeded with cardiac fibroblasts. Cell density was 5 × 105 cells/cm2 as in previous experiments. One week after seeding, seeded (n = 4) and unseeded (n = 12) scaffolds were cut into 25 × 10 mm samples. Uniaxial measurements were performed using a rheological measurement instrument (TA500; Loyd Instruments, Bognor Regis, United Kingdom). Sample edges were clamped so that the exposed area between the clamps was 18 × 10 mm. Device control and data acquisition were performed using Nexygen software (Lloyd Instruments). Throughout the experiments, samples were immersed in PBS (pH 7.4) at RT. Samples were subjected to three sequential mechanical assays, with 10-min intermissions between each assay.
Cyclic strain
Samples were stretched at a fixed rate of 0.05 mm/s to 15% strain (from l0 = 18 mm to lmax = 20.7 mm) and then released at the same rate to the starting point (0 strain). The strain–release cycles were repeated 10 times and served as preconditioning of the macromolecular structure of the matrix. After a 2-min rest at starting point, three more strain–release cycles were performed, and a stress–strain curve was plotted.
Stress–relaxation
Samples were stretched rapidly (0.5 mm/s) to 20% strain (from l0 = 18 mm to lmax = 21.6 mm) and held at that displacement for 10 min, allowing relaxation of stress, and a stress versus time curve was plotted.
Strain to break
Samples were stretched at a fixed rate of 0.05 mm/s until torn, and a stress versus time curve was plotted. The near-linear part of the curve between 10% and 30% strain was divided into four consecutive sections (10–15%, 15–20%, 20–25%, and 25–30%) for which Young's modulus (Y) was calculated as Y = Δσ/Δɛ (Δσ is the subtraction of stress values and Δɛ is the subtraction of the representative dimensionless strain displacement values [(l − l0)/l0] for each section). Complete break was defined as a decrease of 40% stress within 10% strain. Ultimate strength of the matrix was defined as the maximum stress reached before complete break.
Macrophage stimulating assay
To evaluate the immunogenic potential of the acellular scaffold, macrophage cells were stimulated with ECM in vitro, and nitric oxide quantification was performed based on Griess reaction. RAW cell line (TIB-71™; American Type Culture Collection (ATCC), Manassas, VA) cells were seeded in six-well culture plates and cultured in 3 mL of culture medium (same as used for fibroblasts). One day after seeding, the medium was replaced with low-serum medium (2% serum). When 70% confluence was reached, cells were exposed to one of the stimulating agents: acellular cardiac ECM (n = 6) and fresh cardiac tissue (n = 6). Each of these was lyophilized, minced with a scalpel, and 20 mg was gently dispersed in each well. Lipopolysaccharide (LPS; from Pseudomonas aeruginosa, 1 μg/mL; Sigma) served as a positive control (n = 4), and wells with no added agent served as negative control (basal NO secretion, n = 4). The stimulating agents were incubated with the RAW cells for 16 h, and the secreted NO was measured as the free stable nitrite form (NO2−) in the medium, using Griess Reagent System (Promega, Madison, WI), according to the manufacturer's instructions. Nitrite concentrations (μM) were interpolated from a nitrite standard reference curve (0–100 μM sodium nitrite in low-serum medium).
Statistical analysis and micrograph representation
The number of samples is stated individually for each experiment. Data are expressed as mean value ± standard deviation. Two-tailed Student's t-test was used for comparisons at single time points, whereas standard analysis of variance was used when groups were compared through time. p < 0.05 was considered significant for all comparisons. Representative micrographs were chosen randomly from all samples in each experiment.
Results
Validation of acellularity
Complete ECM acellularity was validated using phalloidin-FITC or DAPI staining performed on ECM sections and fresh native tissue. As seen in Figure 1A and B, no evidence for cells (phalloidin-FITC) or cell nuclei residues (DAPI) could be detected in the decellularized scaffolds compared with the native tissue. Camera exposure and microscope operational parameters were identical for both the captured images.
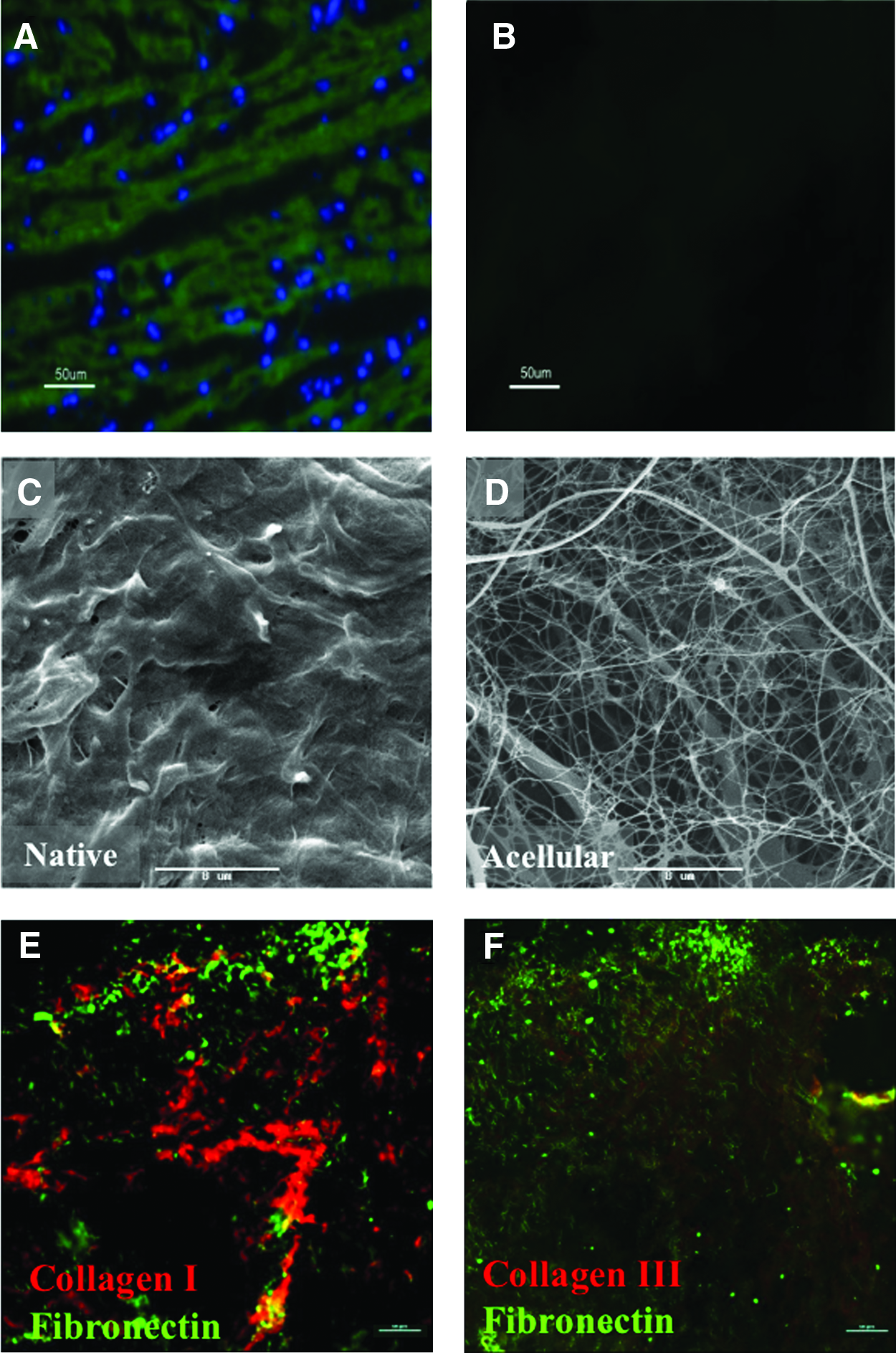
Acellular cardiac scaffold structure and composition. Sections of (
Acellular ECM structure
Scanning electron microscopy (SEM) analysis performed on native and acellular ECM heart tissue (Fig. 1C, D) shows that the resulting acellular structure of the ECM scaffold is a dense interconnective porous network of weaved and strut fibers (Fig. 1D), which do not contain any cellular materials compared with the native tissue imaged under the same parameters (Fig. 1C).
Immunohistochemistry of collagen and fibronectin
Strong immunofluorescent staining for collagen I was obtained in the isolated ECM (Fig. 1E), whereas collagen III resulted in a weaker staining (Fig. 1F). Image data analysis performed using NIS elements software (Nikon Instruments) revealed that the ratio of collagen type I to collagen type III in the acellular ECM is 3.6. This ratio is in accordance to the ratio in the native human heart. 43
Another important ECM constituent that was analyzed is fibronectin. As qualitatively demonstrated in Figure 1E and F, high amounts of fibronectin were found in the acellular scaffold.
Distribution and viability of cells seeded onto the scaffolds
The potential of the ECM matrix to serve as a cardiac scaffold was studied by seeding cardiac fibroblasts, cardiomyocytes, and MSCs on the scaffolds and following their distribution and viability with time. Scaffolds seeded with cardiac fibroblasts prelabeled with DiO exhibited a uniform cell distribution (Fig. 2A–E). By the second week of culture, a continuous process of shrinking of the scaffold was evident. By the fourth to fifth week after seeding, scaffolds became densely populated, forming ∼1- to 2-mm3 spheroids as also demonstrated by SEM imaging (Fig. 2F). Quantomix Wet-SEM images of a fibroblast-seeded matrix 28 days after seeding revealed that the cells populating the scaffold organize in a tissue-like fashion and secrete new fibers—presumably collagen fibers—to their proximate surrounding (Fig. 2G, H). The viability of cells seeded onto the scaffolds was measured at different time points using the noncytotoxic AlamarBlue assay, which enabled assaying each sample at every time point throughout the experiment (Fig. 3A–C). The viability of fibroblasts was approximately 95% after 3 weeks in culture (p < 0.05), showing an increase in tendency for up to 98% between the forth and sixth weeks of culture (Fig. 3A). In comparison, fibroblasts cultured on plate showed elevated viability values after 1 and 2 weeks, which decreased to 60% after 3 weeks. The viability of cardiomyocytes gradually decreased with time and reached 94% and 92% after 3 and 4 weeks, respectively (p < 0.05; Fig. 3B). The viability of control cardiomyocytes showed a similar profile, with 95% and 94% 3 and 4 weeks after seeding, respectively. The viability of MSCs remained approximately the same during the first 2 weeks, reaching 95% by day 24 after seeding (Fig. 3C). The viability of control MSCs increased by the first week, but decreased to 82% and 69% at days 16 and 24, respectively.
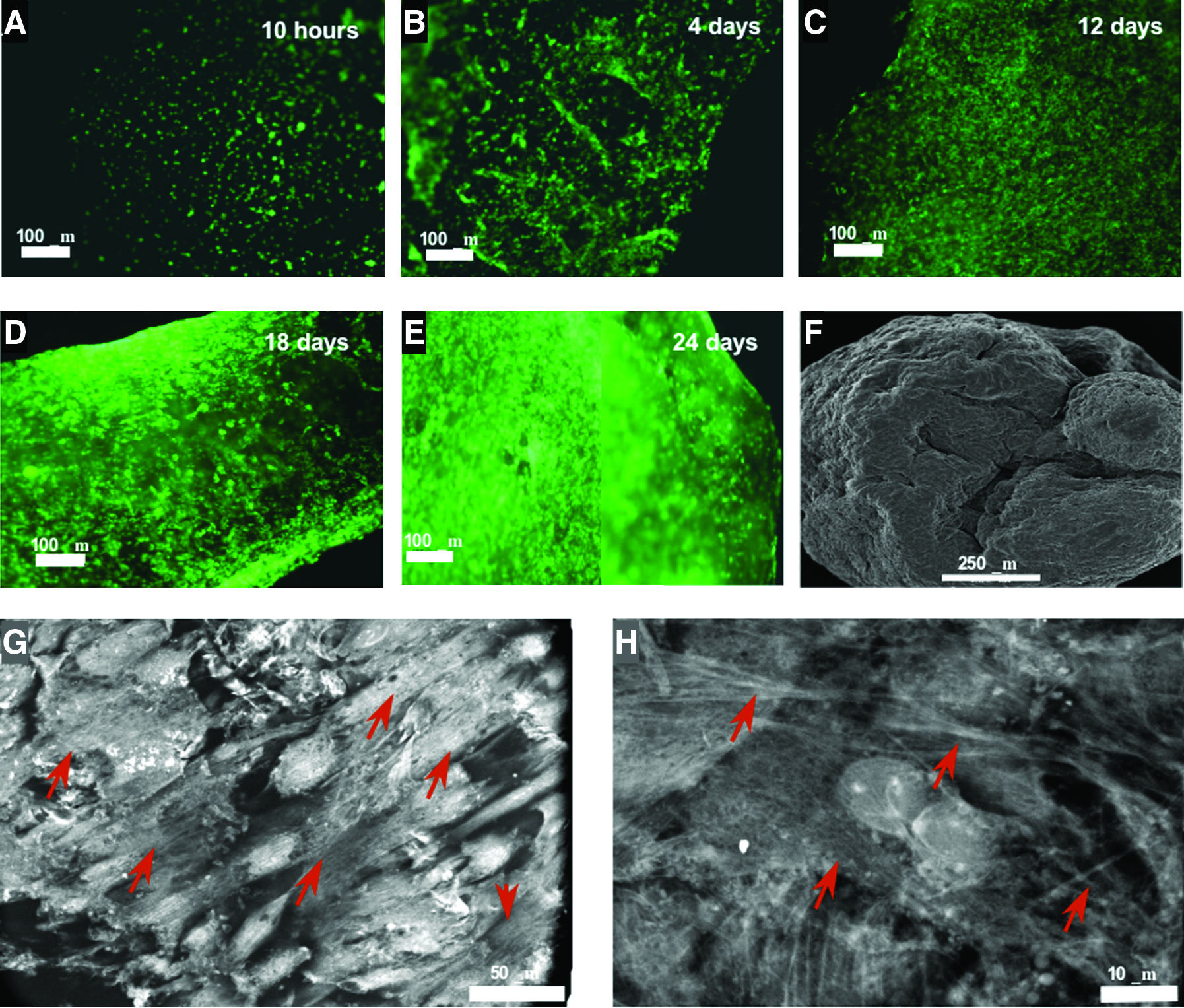
Cardiac fibroblasts seeded onto the acellular cardiac scaffolds. Acellular ECM 10 × 10 mm scaffolds were seeded with 5 × 105 cardiac fibroblasts, to study cell adhesion and distribution on the scaffold. Cells were stained with 3,3'-dioctadecyloxacarbocyanine perchlorate (DiO) before seeding. (

Viability of seeded cells on the acellular cardiac scaffolds. Cell viability was measured using the AlamarBlue assay at different time points after seeding. (
The qualitative fluorescent viability staining assay using FDA/PI revealed that the vast majority of the seeded MSCs (24 days after seeding; Fig. 3D) and fibroblasts (29 days after seeding; Fig. 3E) were viable, as demonstrated by the uptake of FDA. Only few cells were stained positively with PI and were located mainly in dense cell clusters (Fig. 3E).
Immunophenotyping, morphology, and contractility of cardiomyocytes seeded onto the scaffolds
Confocal images of F-actin, cardiac connexin 43, α-actinin, and troponin I indicate that the seeded cardiomyocytes resumed typical differentiated phenotype, expressing cardiac-specific proteins and creating functional gap junctions between adjacent cells, 1 week after seeding (Fig. 4A–D). These images present a focal plane 50 μm deep from the surface, which also demonstrates infiltration of cells from the surface toward the scaffold core. Two weeks after seeding, myocytes exhibited aligned elongated morphology as demonstrated by SEM imaging (Fig. 4F) as compared with their initial arrangement 2 days after seeding (Fig. 4E). Scaffolds seeded with neonatal rat cardiomyocytes began to exhibit spontaneous pulsatile contractions by the second to third day of culture, becoming concert by the third to fourth day (view Supplementary Movie Online, available online at

Cardiomyocytes seeded onto the acellular cardiac scaffolds. Confocal images (50 μm deep) of cardiomyocytes, 1 week after seeding were stained for (
Remodeling of the ECM scaffold by seeded fibroblasts
Collagen and GAG deposition by the seeded fibroblasts was estimated using the hydroxyproline assay and safranin O assay, respectively. As shown in Figure 5A, the scaffold collagen contents increased by 5% after 2 weeks and remained approximately at this level for the rest of the experimental time; however, this increase was not statistically significant (p > 0.05). Total GAG content in fresh cardiac tissue was 3.52 ± 0.37 mg/g fatless dry weight, whereas the acellular scaffolds consisted of 25.46 ± 3.36 mg GAG per gram fatless dry scaffold weight. Scaffolds seeded with fibroblasts resulted in an increase of 23% in the amount of GAGs after 2 weeks of culture (p < 0.05; Fig. 5B), and this level was retained for the rest of the time. Real-time PCR analysis of the expression of ECM remodeling-related genes resulted in different expression patterns for collagens I and III, MMP2, and TIMP1 at the mRNA level (Fig. 6A–D): collagen I increased fourfold during the first week (p < 0.05) and decreased approximately to its initial level by the second week (p < 0.05; Fig. 6A); collagen III level elevated gradually during the first and second week and remained steady during the third week, at 1.8 times its initial level (Fig. 6B); MMP2 increased fivefold by the second week (p < 0.05) and slightly decreased (Fig. 6C); TIMP1, a major MMP inhibitor, rose approximately threefold by the first week (p < 0.05), maintaining this level during the rest of the time (Fig. 6D). Despite the different patterns, the expression of all these genes was elevated throughout the 3 weeks studied, and it was higher than that obtained from control cells seeded on culture plates.
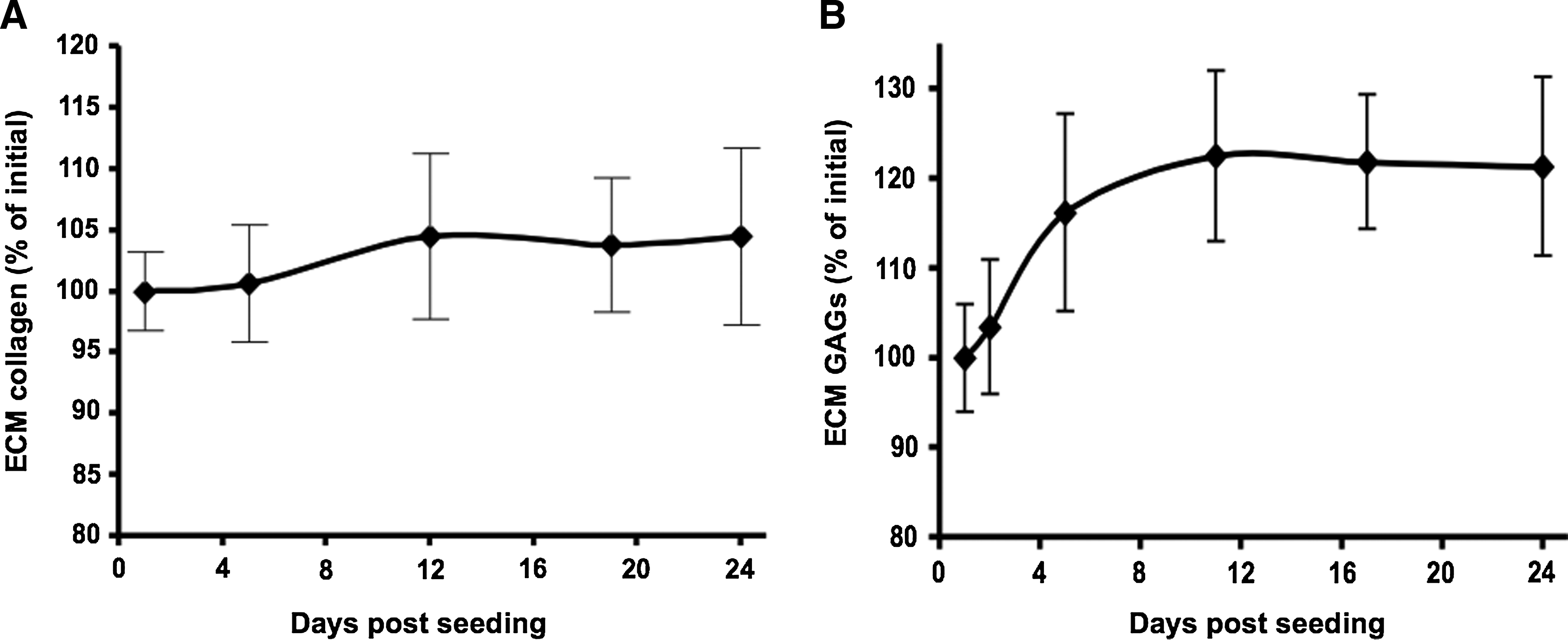
Collagen and glycosaminoglycan contents in fibroblast-seeded scaffolds. Cardiac ECM scaffolds were seeded with 5 × 105 fibroblasts/cm2. (

Expression of ECM remodeling-related genes by fibroblasts. Analysis of gene expression by fibroblasts was performed using real-time polymerase chain reaction for (
Mechanical properties of seeded and unseeded scaffolds
To study the effect of the seeded cells on the tensile properties of the scaffold, scaffolds seeded with cardiac fibroblasts were subjected to cyclic strain, stress–relaxation, and strain to break assays. Unseeded scaffolds served as controls. Figure 7 depicts the results for representative individual samples. Seeded scaffolds qualitatively exhibited reduced elasticity and higher hysteresis, as evident by the distance between the ascending (straining) and descending (releasing) curves in the cyclic strain assay; the reduction of peak stress values for consequent cycles; and by the reduction of minimum stress at starting point (below 0 Mpa; Fig. 7A), compared with the unseeded samples (Fig. 7B). In the stress–relaxation assay, seeded scaffolds exhibited a gradual stress decrease from 0.222 ± 0.058 MPa to 0.18 MPa after 2–3 min. The stress stabilized after 5–6 min and remained 0.16 ± 0.021 MPa for the rest of the time (Fig. 7C). In comparison, the initial stress of 0.211 ± 0.031 MPa in unseeded samples was insignificantly lower (p > 0.05) than that of the seeded samples, yet it dropped quickly to ∼0.14 Mpa, stabilized, and remained significantly lower at 0.132 ± 0.025 MPa for the rest of the time (p < 0.05; Fig. 7C). In the strain to break assay, the seeded scaffolds exhibited a similar profile as with the nonseeded acellular scaffolds. Seeded scaffolds were strained only up to 34.1 ± 6.2% before failure, compared with the 41.3 ± 3.3% for unseeded matrices (p < 0.05), yet exhibited a slightly higher (nonsignificant) ultimate strength value of 0.427 ± 0.103 MPa, compared with the 0.393 ± 0.096 MPa for unseeded matrices (p > 0.05; Fig. 7D). Native cardiac tissue could only be strained to 5.6 ± 1.6% and was torn at 0.745 ± 0.238 MPa (Fig. 7D). The calculated mean Young's modulus was Y = 1.63 ±0.13 MPa for unseeded scaffolds and slightly increased to Y = 1.84 ± 0.27 MPa (p < 0.05) for seeded scaffolds 1 week after seeding.

Mechanical properties of fibroblast-seeded scaffolds versus nonseeded scaffolds. Seeded scaffolds (n = 4) were clamped in a custom-designed apparatus and mounted on a TA500 rheological measurement instrument. All assays were performed after sample preconditioning. (
Macrophage stimulating assay
To achieve a preliminary indication on the biocompatibility of the acellular cardiac ECM, we performed a macrophage stimulating assay in vitro. As shown in Figure 8, the basal nitrite concentration, secreted to the medium by the nonstimulated negative control RAW cells, was 1.47 ±0.34 μg/mL. Stimulation by acellular scaffold resulted in 1.40 ± 0.46 μg/mL nitrite, practically similar to the level obtained by the negative control (p ≫ 0.05). Fresh cardiac tissue stimulated the RAW cells to secrete 6.09 ± 0.66 μg/mL, more than four times the basal concentration (p ≪ 0.05). The positive LPS control strongly stimulated RAW cells to secrete 25.25 ± 1.56 μg/mL nitrite, 17 times the basal level (p ≪ 0.05).
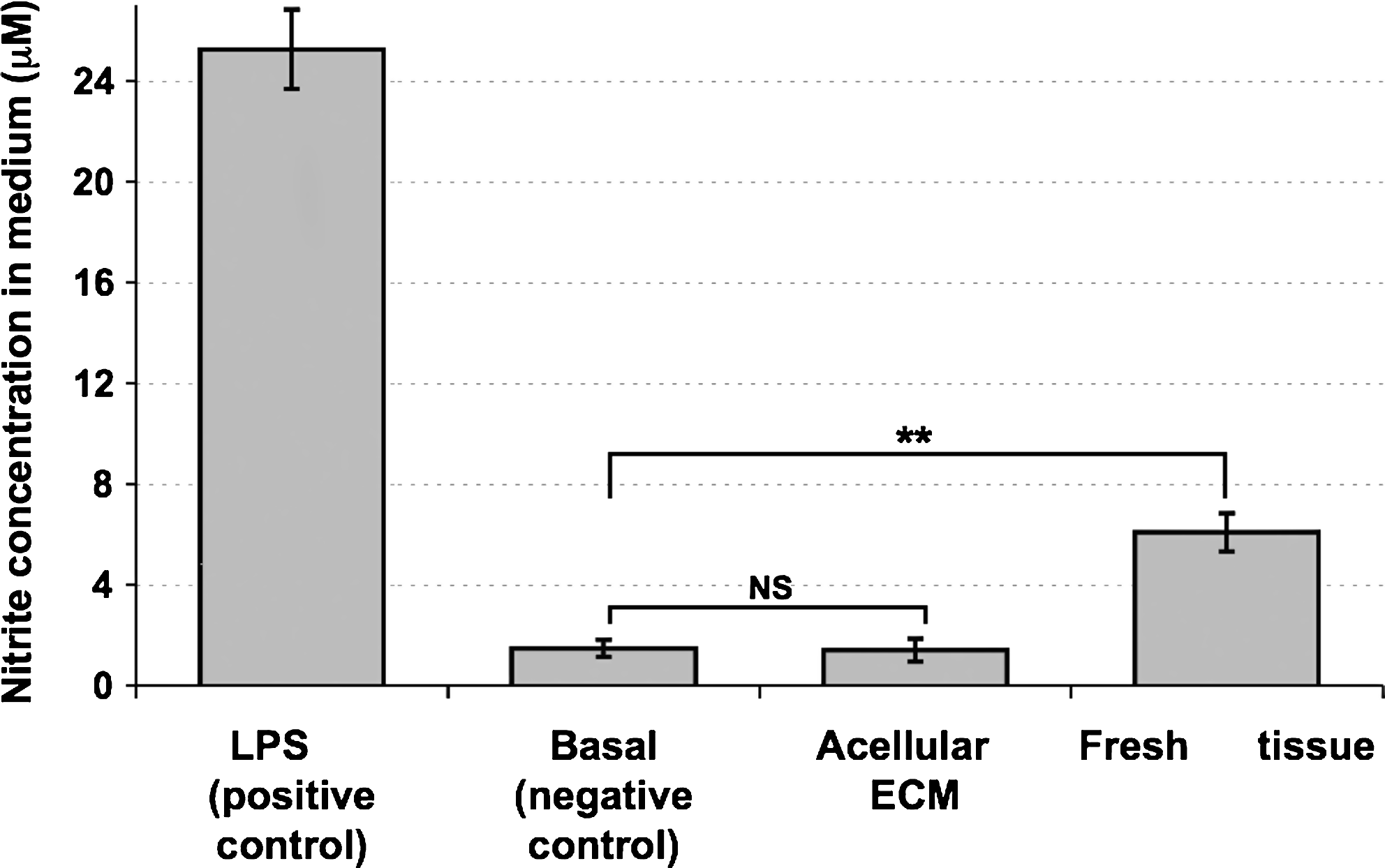
Macrophage stimulation assay in vitro. RAW cells were exposed to stimulating agents for 16 h, and nitric oxide levels in the medium were then measured: acellular ECM scaffold and fresh cardiac tissue (n = 6 for each). Stimulation with lipopolysaccharide (LPS) served as positive control (n = 4) and nonstimulated cells served to set the basal nitric oxide level (n = 4). **p < 0.05.
Discussion
Several decellularization protocols have been reported for the isolation of ECM from different organs, which include mechanical, enzymatic (DNAs, RNAs, collagenase), and detergent (sodium dodecyl sulfate, Triton-X-100)-based procedures.29,37,44 Recently, a work performed by Ott et al. described the acellularization of cardiac-derived ECM that was also based on SDS perfusion. 38 This group has demonstrated the efficacy of such ECM for the remodeling of rat heart, achieving an undisrupted cellular growth for 8 days. 38
In this study, we describe a decellularization process of porcine cardiac tissue based on a combination of hyper–hypotonic washes, enzymatic digestion, and nonionic detergent treatment with Triton-X-100. Although enzymatic digestion had been suggested to disrupt the ECM structure, 29 we have found that the acellularized porcine cardiac ECM exhibited structural weaved and strut fibers as expected from the myocardial architecture (Fig. 1D), with no apparent traces of cellular components, as depicted in the native tissue. Furthermore, the acellular cardiac ECM consisted of fibronectin and collagen type I to collagen type III ratio similar to that of the native human heart (Fig. 1E, F), preserving 23% of the its GAG content (unpublished data).
To validate the potential of this decellularization procedure and the resulting ECM suitability to serve as a scaffold for cardiac tissue engineering, we have studied its biochemical and mechanical properties and the interaction of this specific ECM with different types of seeded cells. For these studies, several primary cells including adult sheep cardiac fibroblasts, neonatal rat cardiomyocytes, and adult rat bone marrow MSC cells were used. These cells were chosen as they had been commonly used in different cardiac tissue engineering studies.10–12 For these studies, we have used AlamarBlue, a metabolism-based assay, which indicate for cell vitality and proliferation rate of the cells at any time point, rather than only proliferation. The change in the viability during the period tested was mostly insignificant for the myocytes (8%) and MSCs (5%). The viability of myocytes on the scaffold was similar to that measured on culture plate controls during the 4 weeks tested, most likely because these cells lack the ability to divide and propagate. The rapidly propagating control fibroblasts expanded on the plate to complete confluence within a week. Nevertheless, after 3 weeks this culture became overcrowded and the viability levels dropped dramatically. On the other hand, fibroblasts seeded onto the scaffold propagated at a significantly lower rate, which explains the stability in their viability on the scaffold for an extended period of time. Seeded MSCs retained their viability during the first 2 weeks, and their propagation rate was much lower than that of the fibroblasts, which explains the smaller viability increase after 1 week in the control cells. The seeded MSCs remained viable throughout the experiment, whereas the control decreased. The qualitative “live/dead” assay, using FDA/PI fluorescent staining, which enables to estimate cell death during culture, indicated that even after 3–4 weeks after seeding, damaged or dead cells (positive for PI) are scarcely found on the scaffold. The results from these assays indicate that the scaffold supports the viability of all the three seeded cell types for prolonged periods of time compared with the control cells seeded on the plate.
The interaction between the scaffold and seeded cardiomyocytes was studied by immunofluorescence staining of key factor proteins, which are commonly used to characterize these cell's phenotype. One week after seeding, cardiomyocytes expressed proteins such as α-actinin, troponin I, and connexin43. These are representative markers for the contractile action of the cells as well as the action potential conductivity through functional cytoplasmic gap junction. 45 Most importantly, several days after seeding, spontaneous contractions of the scaffold were clearly visible, becoming synchronized beating shortly after. Evidence for the cardiac-like tissue morphology was further supported by SEM images taken 2 weeks after seeding. Cardiomyocytes on the ECM scaffolds developed aligned elongated morphology, which resembles the morphology of mature cardiac myofibers, and is significantly different than the typical morphology found when cardiomyocytes are cultured on the plate.18,46 This phenomenon may be attributed to longitude “guidance” directed by major elongated collagen fibers (assumed to be type I) of the natural cardiac ECM.
The ability of cells to remodel ECM-based scaffolds is of great importance and has been discussed before.47,48 Macroscopically, it was evident that the fibroblasts can remodel the scaffold as scaffold shrinking became apparent two weeks post-seeding. At first, this may appear to be due to the loss of scaffold collagen mass, as a result of enzymatic digestion by the seeded cells. However, the collagen content of seeded scaffolds has been maintained during culture. Further, newly secreted fibers—presumably collagen fibers—can be seen surrounding the cells, as depicted by the Quantomix Wet-SEM images of fibroblast-seeded scaffold after 4 weeks in culture. Importantly, the total GAG content in fibroblast-seeded scaffold has been increased significantly during culture (23% by day 12), which may be as a result of the beneficial interaction between the scaffold and the populating cells.16,49,50 Considering these evidences, it is clear that the shrinkage of fibroblast-seeded scaffolds with time occurred due to the biochemical and physical rearrangement of scaffold structure and volume, and not due to the loss of constituent's mass.
We have also studied the mRNA expression profile of representative ECM remodeling-related genes, using real-time PCR. We chose to focus on collagens I and III, the main cardiac ECM constituents, MMP2, which is one of the main ECM decomposing factors, and TIMP1, a major MMP inhibitor. Together, these genes can help to create a general picture of the “building versus breaking” status of the cells on the scaffold. Generally, it can be stated that the gene expression levels of the four genes studied were higher than those of the control cells that were cultured on the plates. These findings further support our observation that the seeded cells interact with the scaffold. The significant increase in the collagen I mRNA level after 1 week may explain the preservation of collagen I in the scaffold. The increase in the MMP2 mRNA level, peaking at the second week, may explain the change in the scaffold volume, which begins approximately at that time. However, the mRNA level of TIMP1 was elevated after 1 week, which may seem contradicted to the activity of MMP2. The level of collagen III mRNA was gradually elevated when compared with the collagen I mRNA level, probably because it is a more minor ECM constituent. Thus, we hypothesize that the anabolic and catabolic processes resulting from the remodeling of the scaffold by the fibroblasts are the end result of a complex feedbacked cascade, of which these genes are only representatives. Further investigation of this cascade at the protein level may clarify the enzymatic activity and inhibition kinetics.
To assess the effect of seeded cells on the mechanical properties of the scaffold, we have measured the tensile properties of scaffolds seeded with fibroblasts and compared them with the tensile properties of the unseeded scaffolds. The assays performed in our study are of passive uniaxial nature and do not attempt to mimic the active contraction properties of a live functional myocardium, where myocytes are responsible for muscle contractions, whereas fibroblasts and ECM contribute more to the passive relaxation. The seeded scaffolds seemed more hysteretic compared with the nonseeded scaffolds (cyclic straining) and their strength was increased, whereas their elasticity decreased (strain–relaxation and strain to break). Not surprising, therefore, was the observation that the native tissue exhibited much higher strength and much lower elasticity values (strain to break). Similar observations were found by Feng et al., who studied the mechanical properties of contracted collagen gels by means of uniaxial tensile testing. 51 In the study by Ott et al., the stress–strain curve of native as well as decellularized rat left ventricle was demonstrated. The calculation of the tangential modulus and mechanical stiffness was performed at 40% strain in both the longitudinal and circumferential directions. The mechanical data presented by Ott et al. cannot be compared to our data as our slices, originated from porcine origin, which are by far thicker and larger than those tested by Ott et al. In addition, the strain used by Ott et al. (40%) does not characterize this biomaterial in a manner which resembles the physiological conditions of normal cardiac tissue, and is beyond the plastic deformation of both decellularized as well as native tissue as obtained from our study.
Finally, we have attempted to examine the biocompatibility of the isolated cardiac ECM, as this is a major requirement from a tissue-engineered scaffold. The ultimate study of immunogenic potential should eventually be performed in vivo. 52 However, some in vitro immunological assays can be performed before in vivo studies, as indicatory assays. Therefore, we have used a macrophage stimulation assay that was performed on macrophage cell line. Once the cells are stimulated they secrete nitric oxide that is indicative of the level of their stimulation, thus to the immunogenic potential of the stimulating agent. From the results, it is clearly evident that the acellular ECM scaffold is inert to the RAW cells. This is in contrast to the stimulation observed with native tissue or LPS (positive control), leading to a significant release of NO. The results further emphasize the importance of an efficient decellularization protocol and acellularity assays for biomaterials that originate from xenogeneic tissues. Further in vitro investigations of the immunogenic potential of the acellular cardiac ECM scaffold are required, such as stimulation of macrophages to express proinflammatory genes (interleukins, tumor necrosis factor, etc.) and are currently performed in our laboratory.29,52
In conclusion, this study shows that the porcine acellular cardiac ECM is a nonimmunogenic scaffold that can support the prolonged viability of cardiac fibroblasts, cardiomyocytes, and MSCs. The cardiac ECM promotes mature and functional cardiomyocyte phenotype, triggers remodeling, and secretion of GAG. These findings clearly indicate the advantages of porcine acellular cardiac ECM as a biomaterial scaffold for cardiac tissue engineering.
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.