
Abstract
We have previously established a chamber model of tissue engineering that promotes de novo angiogenesis and vascularization of engrafted cells and tissues when combined with an extracellular matrix. Here we demonstrate that the mouse chamber (MC) model can sustain ectopic grafts of murine fetal thymus lobes and, to a limited degree, human pediatric thymus tissue, resulting in de novo T-cell production. Silicone chambers containing Matrigel® and thymus tissues were placed around exposed epigastric vessels and the ends sealed with bone wax, before implantation into the inguinal fat pad of athymic Balb/c nu/nu (nude) mice. Murine, embryonic day 15 (E15) thymus grafts were found to be well vascularized and viable within the MC upon harvest at week 11. In contrast, engraftment of both adult murine and pediatric human thymus tissue was limited, with only one out of the seven human thymus grafts sustaining mature, murine-derived T-cell development. Increased CD4+ and CD8+ T-cell numbers were observed in the peripheral blood of nude mice within 2 weeks after E15 thymus-MC grafts (n = 8), compared with nude control mice. Peripheral blood T-cell percentage and subset distribution were comparable to mice receiving conventional thymus kidney capsule grafts. T-cell function of both kidney capsule- and MC-E15 thymus grafts was established via successful rejection of major histocompatibility complex (MHC)-mismatched skin grafts. Sustained growth of fetal thymus tissue in the MC provides an alternative model for the study of thymopoiesis and related applications of T-cell–mediated immunity.
Introduction
The thymic stroma is predominately epithelial in nature—interspersed with mesenchymal-derived fibroblasts, macrophages, and antigen-presenting dentritic cells, organized into a three-dimensional (3D) meshwork to facilitate maximal interaction with migrating T cells. 2 During organogenesis, branches of the brachial artery penetrate the thymic capsule and establish an intricate network of vessels and capillaries that provide both nutritional support and continual replenishment of blood-borne T-cell progenitors to sustain postnatal thymopoiesis. 3
The study of thymopoiesis is relatively difficult to explore in vitro. Thymic epithelium requires maintenance of a 3D structure in culture for positive selection of T cells; a monolayer culture destroys this property. 4 As such, functional studies have relied heavily on the use of fetal thymus organ culture systems or reaggregates of fetal thymic stromal cells, in which limited thymopoiesis can be obtained in situ. 5 Ectopic grafting of fetal thymus tissue under the kidney capsule (KC) has further allowed in vivo analysis of thymic function by promoting neovascularization of ectopic tissue and subsequent entry of systemic, host-derived T-cell progenitors to form a functional thymus unit (e.g., Refs.6–8 ). To this end, conjoint, ectopic human fetal liver- and human fetal thymus-KC grafts have been utilized in the establishment of humanized mouse models to facilitate the study of human T-cell–mediated immunity and disease.6,9,10
Recently, in vivo chamber models have been developed in rats and mice as part of a tissue engineering methodology to promote de novo, vascularized tissue formation.11–14 In the mouse chamber (MC) model, a split silicone tube (∼5 × 3 mm) is implanted around the epigastric vessels of the groin vascular bed, which is then filled with an extracellular matrix and sealed at both ends with bone wax. Vascularized, flow-through chamber vessels containing the extracellular matrix, Matrigel™ (BD Biosciences, San Jose, CA), were shown to promote high levels of angiogenesis and generation of de novo adipose, breast-like acini and duct tissue. 14
Here we demonstrate the ability of subcutaneous, inguinal-MC grafts containing Matrigel supplemented with fibroblast growth factor (FGF-2) to support the growth and function of murine fetal thymus lobes for potential application in the study of thymopoiesis and T-cell–mediated immunity. Successful engraftment and vascularization of fetal thymus tissue were demonstrated in nude (Balb/c nu/nu ) mice using the MC model and was comparable to mice receiving conventional thymus grafts under the KC. Sustained repopulation of thymus tissue was evident at 11 weeks after graft, with both groups demonstrating increased peripheral T-cell levels above nude controls by week 2. T-cell function of both thymic-KC and MC engrafted mice was determined via successful rejection of third-party major histocompatibility complex (MHC)-mismatched skin grafts. The MC, however, supported only limited growth of postnatal murine and human thymus tissue, indicating further manipulation of the MC environment may be required to sustain growth in these instances.
Materials and Methods
Animals
C57Bl/6 mice were obtained from Monash Animal Services and housed at Monash University Mouseworks facility (Monash University, Victoria, Australia) according to the institutional guidelines. Balb/c and Balb/c nu/nu mice were obtained from Animal Resource Centre (Perth, Australia) and housed at the Experimental Medical and Surgical Research Unit, St. Vincent's Hospital (Melbourne, Australia).
All surgical experiments were performed under the ethical approval of Monash University and St. Vincent's Hospital Animal Ethics Committee, under the National Health and Medical Research Council (Australia) guidelines.
Human tissue
With patient consent, a human thymus tissue sample was obtained from a child undergoing corrective cardiac surgery at the Royal Children's Hospital, from which thymus tissue is routinely removed to access the heart. Ethical approval was obtained from the Ethics in Human Research Committee at the Royal Children's Hospital (Melbourne, Australia) and used in accordance with the institutional guidelines.
Animal treatments
Anesthesia, sterility, and pain relief
Male, nude mice (20–26 g body weight) were used in this set of experiments. Surgery was performed in sterile conditions under general anesthesia, induced by an intraperitoneal injection of chloral hydrate (0.4 mg/g body weight). The skin was treated with chlor-hexidine and alcohol for sterility and carprofen administered for pain relief.
MC surgery
Tissue engineering chambers consisting of 5 mm lengths of silicone laboratory tubing (3.35 mm internal diameter, 44 μL volume; Dow-Corning, Midland, MI) were inserted into one groin of the mouse as described by Cronin et al's method. 14 Briefly, the superficial epigastric vessels were exposed via a transverse groin incision and dissected free of their investing fascia for a distance of approximately 1 cm, from their origin at the femoral vessels to their entry into the groin fat pad. The split cylindrical tissue engineering chamber was then placed around the vessels and anchored to the underlying muscle with a 10-0 nylon microsuture. The proximal aperture and longitudinal seam of the chamber were sealed with melted bone wax (Ethicon, Somerville, NJ), and the chamber was filled with thymus tissue sample and Matrigel containing 1 μg/mL recombinant human FGF-2 (Peprotech, Burlingame, CA). Following this, the distal aperture of the chamber was sealed with bone wax and the construct carefully returned to the dissected space within the groin. The wound was closed with metal clips. At harvest the constructs were delivered via the transverse groin wound and examined for vessel pedicle patency. Constructs with an absent or infarcted pedicle were discarded from the study.
KC grafts
To perform KC surgery, the mouse was shaved on the left side between the hip and upper rib cage and wiped clean with ethanol. An intraabdominal incision was made (∼0.5 cm), running parallel but just below the spine, in between the hip and the bottom of the rib cage, to expose the kidney. The kidney was attached to a fat pad and brought outside the incision by gently grasping the fat pad with forceps and levering the kidney out. A small hole was made in the KC using two straight fine forceps and the thymus tissue placed under capsule. Kidney was replaced into abdominal cavity and muscle tissue sutured with dexon 4.0 before suturing skin closed. Mice were given Caprofen® (Pfizer, NewYork, NY) for pain and monitored daily. At the end of the experimental period, mice were sacrificed and whole kidney containing thymus graft removed.
Skin grafts
Donor C57Bl/6 mice were first shaved of their hair, before a piece of skin measuring 1.5 cm in diameter was dissected as a full-thickness skin graft and was then transplanted to the back of the recipient nude mice with 6-0 nylon continuous suture. For fixing the skin graft to the recipient mouse, a Wolf–Krause dressing was applied on the top of the skin graft and removed 5 days postoperatively. Animals and skin grafts were monitored daily.
Antibodies and immunoconjugates
The following fluorochrome- or biotin-conjugated antibodies used in this study were purchased from BD Pharmingen (San Diego, CA) unless otherwise stated. Murine antibodies: anti-CD8α (clone 53-6.7), anti-CD4 (clone GK1.5), anti-T-cell receptor beta (TCRβ; clone H57-597), anti-CD44 (clone IM7), anti-CD62L (clone MEL-14), anti-CD31 (clone MEC13.3), and pan-cytokeratin (DakoCytomation, Glostrup, Denmark). The secondary reagent: streptavidin–APC, streptavidin–AlexaFluor488, anti-rat Ig AlexaFluor488, and anti-rabbit Ig AlexaFluor568 (Molecular Probes, Eugene, OR).
Histology
Immunohistochemistry
MC contents were harvested from the body and cleaned to remove excess fat and connective tissue. Thymic grafts were suspended in Tissue-Tek O.C.T compound (Sakura Finetek, Zoeterwoude, The Netherlands) and snap frozen in a liquid nitrogen and isopentane slurry. Sections of 8–10 μm were cut on a Leica CM1850 cryostat (Leica Microsystems, Bannockburn, IL) and left to air dry at 4°C for 30 min. Primary antibodies were added to sections, and slides were incubated at room temperature for 15 min followed by washing in phosphate-buffered saline for 5 min. Secondary antibodies were then applied to sections, which were incubated for 15 min, washed, and then mounted with a coverslip using fluorescent mounting medium (DakoCytomation). Images were acquired on a Bio-Rad MRC 1024 confocal microscope and analyzed using LaserSharp2000 software (Bio-Rad, Hercules, CA).
Hematoxylin and eosin
MC contents or kidney–thymus grafts were removed and cleaned to remove excess fat and connective tissue. Tissue was fixed overnight in 4% formaldehyde before being processed and embedded into paraffin. Sections of 5 μm were fixed onto the slides and dried overnight in a 37°C incubator, before staining with hematoxylin and eosin.
For costaining with CD31, tissues were fixed in paraffin, cut into 5 μm sections, and then subjected to a 3% H2O2 block followed by antigen retrieval by Proteinase K (DakoCytomation) for 8 min. Slides were then subjected to a protein block for 30 min before incubation with a primary rat anti-mouse CD31 antibody (BD Pharmingen) at a concentration of 1:1150 for 60 min at room temperature. Slides were washed with tris-buffered saline and incubated with a rabbit anti-rat biotinylated secondary antibody (DakoCytomation) at a concentration of 1:200 for 30 min at room temperature. Cells were then washed with tris-buffered saline, and the target was detected using a VECTASTAIN Elite avidin: biotinylated enzyme complex (ABC) Kit and DAB chromogen (Vector Laboratories, Burlingame, CA) counterstained with hematoxylin, cover slipped using dystyrene plasticizer and xylene (DPX) and left to dry overnight.
Enzymatic digestion of thymus-MC grafts
The contents of MC-thymus grafts were subjected to an enzymatic digestion to help dissociate the thymus tissue from the adipose tissue. Chamber contents were minced into small pieces and incubated with a Collagenase/DNase mix (Roche, Basel, Switzerland) in RPMI-1640 for 30 min at 37°C. To aid tissue dissociation, chamber contents were manipulated with a glass pipette of decreasing bore size. Resulting cell suspensions were passed through 100-μm mesh to remove debris and centrifuged to recover cells. Cell counts were performed using a Z2 Coulter counter (Beckman Coulter, Fullerton, CA).
Flow cytometry
For flow cytometric analysis, 5 × 106 cells from murine- or human thymus-MC digests were stained with appropriate antibodies. Cells were acquired on a FACScalibur (BD Biosciences, San Jose, CA) using up to four fluorescent channels and analyzed using CellQuest software (BD Biosciences).
Statistical analysis
Statistical analysis was performed with SPSS version 15.0 software (Chicago, IL) using the Mann–Whitney U test.
Results
MC supports ectopic murine fetal thymus grafts
To investigate the potential of the MC model in supporting the growth of thymus tissue, chambers were surgically implanted around the epigastric vessels of the groin fat pad of Balb/c nu/nu (nude) mice, as described previously. 14 The athymic, nude mouse model was used to study thymus engraftment as it harbors a defective foxn1 gene, which inhibits development of the endogenous thymus organ; however, their hemopoietic stem-cell compartment remains capable of repopulating normal donor thymus tissue. After implantation, MC was filled with Matrigel and engrafted with one Balb/c embryonic day 15 (E15) thymus lobe. As a comparison, nude mice also received an E15 thymus lobe graft under the KC. Nude mice received only one MC- or one KC-E15 thymus graft per mouse; n = 8 per MC and KC group. Peripheral blood was collected at 2, 4, and 8 weeks after graft and analyzed via flow cytometry for the presence of mature αβ T cells, characterized by the expression of the TCRβ subunit. At week 8, MC and KC nude mice groups were grafted with MHC-mismatched C57Bl/6 donor skin grafts. As a control, donor skin was also grafted onto untreated nude mice (n = 5). Both MC and KC grafts were harvested at 3 weeks after skin graft (week 11).
Upon surgical removal of E15 thymus grafts at week 11, a patent pedicle was observed for all MC grafts, and contents were found to be well vascularized (Fig. 1 and data not shown). Both KC- and MC-thymus grafts were observed by hematoxylin and eosin staining (Fig. 2). An intact lymphocyte-filled structure was found in all samples from the KC and MC groups with E15 MC-thymus grafts being of similar size, and in some samples, larger than the KC-thymus grafts (representative histology shown in Fig. 2A, B). The grafts of both groups presented with a classic thymus morphology, exhibiting darker staining cortical areas and lighter staining medullary regions due to the greater density of lymphoid cells in the cortex than the medulla (Fig. 2C, D). Vascularization of thymus tissue from both graft sites was confirmed by counterstaining with anti-CD31 antibodies (Fig. 2E, F).
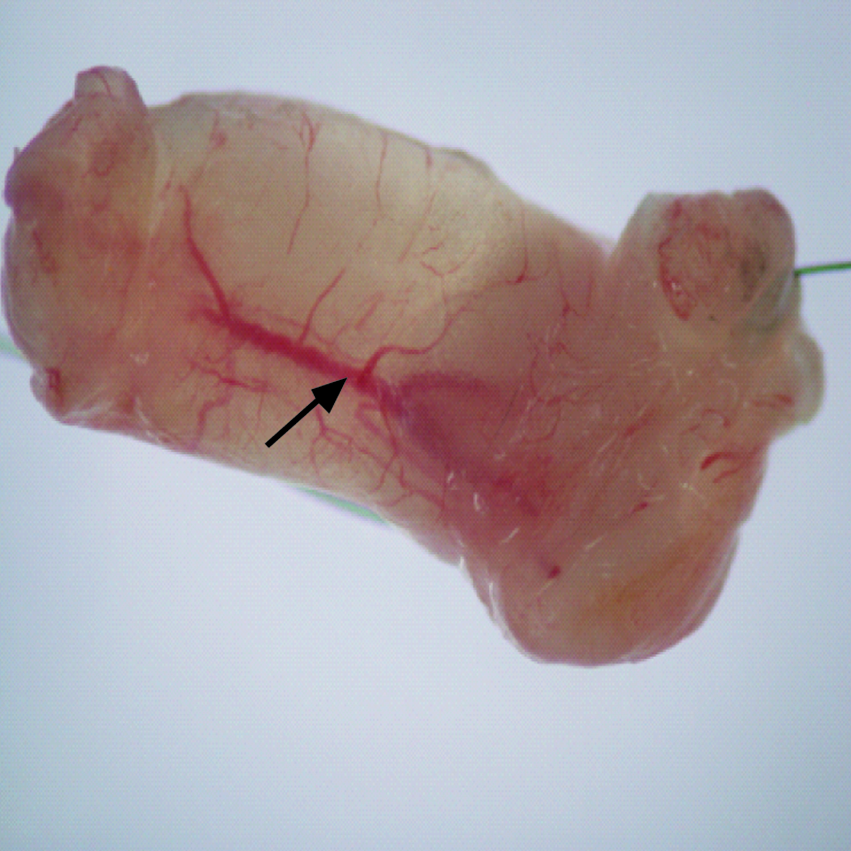
Mouse chamber (MC) contents at week 11 after embryonic day 15 (E15) thymic lobe graft showing evidence of vascularization (arrow).
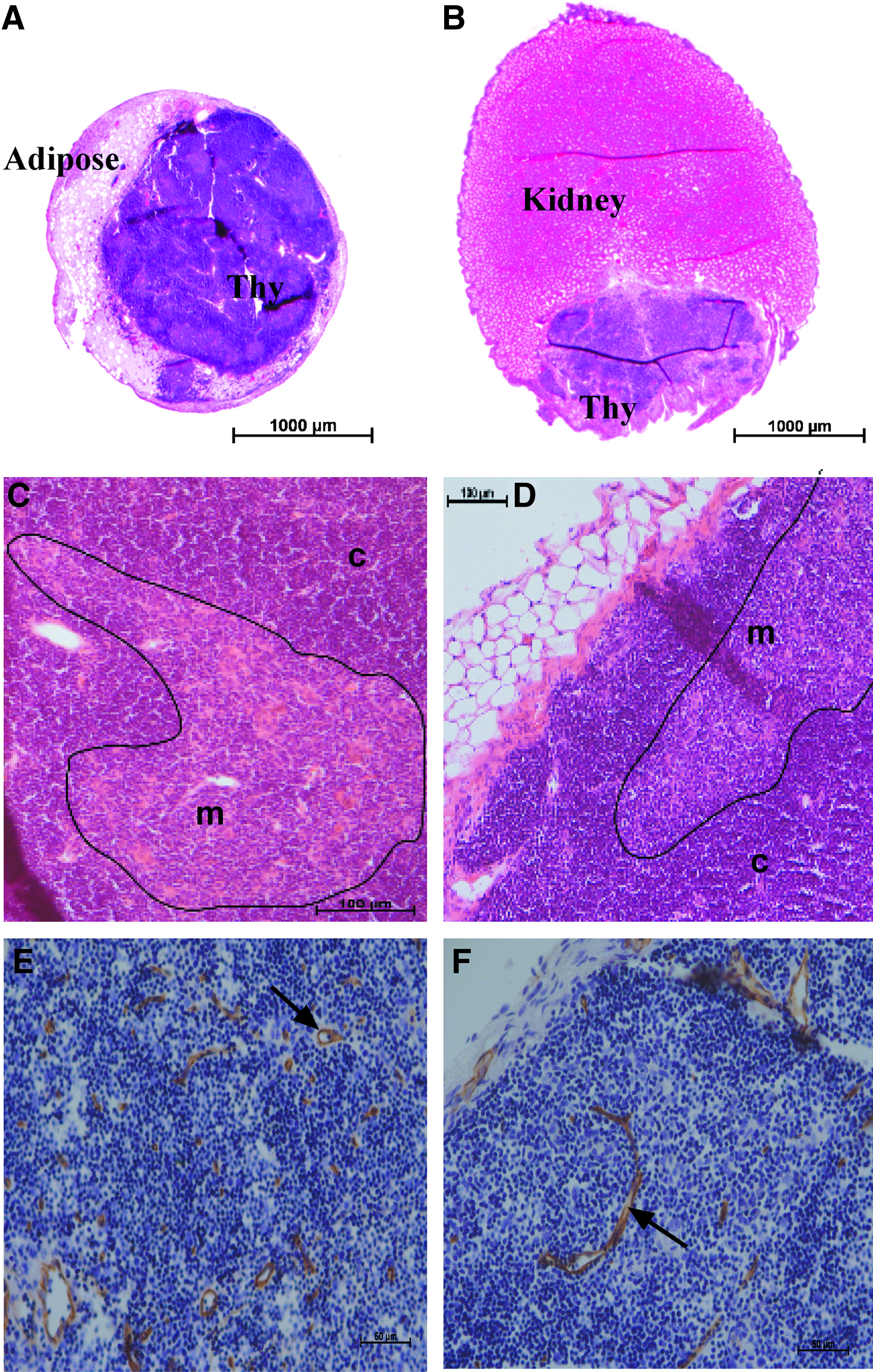
Representative histology of MC contents (
Assessment of αβTCR+ T-cell proportion in the peripheral blood of nude mice receiving either MC- or KC-E15 thymus grafts demonstrated significantly increased proportion of T cells in both groups, at 2, 4, and 8 weeks after grafts, compared with nude control mice (Fig. 3A). It is interesting to note that at 2 weeks, significantly less peripheral T cells were found in the MC group compared with the KC group. This may be due to an increased time taken to vascularize the thymus tissue within the MC compared with the KC, and thus delayed export of T cells into the circulation. At week 8, the peripheral T-cell pool of the MC- and KC-E15 thymus groups was still markedly lower compared with control Balb/c levels. This is most likely explained by the large size difference, and therefore T-cell output, between an embryonic thymus and the endogenous, adult thymus of Balb/c control mice. De novo T-cell production requires 2–3 weeks, from immature to mature T cell, and thus only limited T-cell production would have occurred in the embryonic thymus graft within the 8-week period of this experiment.15,16 Despite the lower peripheral T-cell percentage, analysis of TCRβ+ CD4+ and CD8+ mature T-cell subset distribution within the MC and KC groups revealed that both subsets were represented at a similar proportion to Balb/c control mice, indicating that comparable T-cell development was occurring at both ectopic graft sites (Fig. 3B).

(
To assess the function of the peripheral T cells, both the KC and MC groups received allogeneic C57Bl/6 donor skin grafts to assess their ability to mediate rejection of the MHC-mismatched tissue. Donor skin was initially accepted within 1–3 days after surgery (data not shown) in all the three MC, KC, and nude control mouse groups. Donor skin started showing signs of rejection between days 10 and 14 after skin graft in both MC and KC groups (Fig. 4B) and had been completely rejected (i.e., detached from mouse body) by 3 weeks. In contrast, unmanipulated nude control mice completely accepted the donor skin grafts (Fig. 4A).

(
MC supports limited growth of murine adult thymus lobes and pediatric human thymus
Murine fetal thymus lobes can be readily sustained both in culture and as an ectopic tissue graft. Growth of adult thymic tissue, however, is more difficult given their larger size and reduced plasticity. To assess the ability of the MC to support growth of the adult murine thymus tissue, we placed 8-week-old Balb/c thymic lobes (n = 7) into separate MC vessels containing Matrigel and engrafted the chamber into the groin fat pad of nude mice (deemed A1–7), as described previously for E15 thymic grafts. We also engrafted small pieces (∼2 × 2 mm) of a 4-month-old human thymus tissue sample (n = 7) into separate MC-implanted nude mice (deemed H1–7) to assess the ability of the MC to support human thymus function. Due to potential human T-cell xenoreactivity, mice were monitored for signs of xenograft-versus-host disease. During the 12-week period, no mice that received a human thymus-MC graft exhibited obvious signs of disease, such as wasting, lethargy, or increased infections. At 12 weeks, mice were culled and MC contents and spleen removed. MC containing murine adult thymus tissue (A1 and A2) and human thymic tissue (H1 and H2) was prepared for histological analysis. MC A3–7 and MC H3–7 were enzymatically digested and stained with T-cell markers to assess presence of thymocyte subsets by flow cytometry.
As the thymic microenvironment is predominately epithelial, we used a pan-cytokeratin Ab to observe the thymus-MC graft after 12 weeks. Histological examination of graft A1 and A2 via immunohistochemistry revealed the presence of keratin-positive areas within the MC graft indicative of thymus tissue. Unlike the E15 thymus grafts, however, where an intact thymus organ was observed, the adult thymus grafts were not well formed and were often fragmented (Fig. 5). Subsequent staining with anti-CD31 to label endothelial cells revealed that some keratin-positive tissue appeared vascularized (Fig. 5A), whereas other fragments remained avascular (Fig. 5B). In the case of the two human thymus-MC samples chosen for histological analysis, both samples H1 and H2 revealed no visibly intact thymus and prominent adipocyte infiltrate (data not shown).

(
Intrathymic thymocyte development occurs via differentiation of immature T-cell progenitors double negative (DN) for CD4 and CD8, into a subset double positive (DP) for both markers, before selecting either CD8 or CD4 expression to become mature, single-positive (SP) T cells. 17 Upon enzymatic digestion of MC contents, samples A3 and A5–7 as well as H3, H4, and H7 appeared to consist entirely of fatty tissue and did not stain for anti-mouse T-cell markers: αβTCR, CD4, or CD8 by flow cytometry (data not shown). In contrast, MC thymic grafts A4, H5, and H6 demonstrated the presence of CD4 and CD8 SP populations as well as a DP subset, indicating that a degree of intrathymic T-cell development had occurred and was still persisting within both the adult murine- and pediatric human thymus-MC grafts after 12 weeks (Fig. 6A). Representation of thymocyte subsets, however, was skewed in both A4 and H6 samples, with a decreased proportion of DP cells and an increased number of cells of DN phenotype. Interestingly, human sample, H5, was found to have a highly comparable thymocyte subset profile to that of an endogenous control Balb/c thymus, indicating that murine, bone marrow–derived T-cell progenitors had seeded the human thymus-MC graft and differentiated within the thymic microenvironment (Fig. 6A). Thymocyte subsets upregulate TCRβ as they progress from an immature DN subset to mature CD4/CD8 SP phenotype. 18 TCRβ histogram profiles revealed similar low, intermediate, and high TCRβ peaks between the endogenous Balb/c thymus and the H5 thymic graft, whereas MC-thymic grafts A4 and H6 had detectable, but reduced TCRβ peaks (Fig. 6B). Further subgating on sample H5 revealed that the high-level TCRβ expression was restricted predominately to the SP T-cell subsets, as would be expected during normal thymopoiesis (data not shown).

(
To examine whether the adult murine and pediatric human thymus-MC grafts could contribute to the peripheral T-cell pool, we examined splenocyte populations from the engrafted nude mice groups. Thymic-derived T cells are exported to the periphery as naive, unstimulated T cells and can be defined by the positive expression of CD3 and CD62L, with negative to low levels of CD44 (CD3+ CD62L+ CD44−/lo). 19 Predictably, splenocytes from athymic nude mice displayed negligible presence of naive T cells, whereas approximately 15% of the T cells from wild-type Balb/c mice were defined as naive (representative plots shown in Fig. 6C). No significant increases in naive T-cell proportion was found in any nude mouse receiving adult murine thymus-MC grafts (A1–7) compared with nude control splenic levels, including mouse A4 that displayed limited thymopoiesis (Fig. 6C and data not shown). Increased percentage of αβTCR+ T cells, however, was found within the spleen of mouse A4 (13.2%) compared with nude control levels (1.8%) (Fig. 6D). Analysis of splenocytes from mice engrafted with human thymic tissue samples (H1–7) revealed negligible naive T-cell staining in all samples (data not shown) except H5, in which a relatively small naive T-cell population was detected (1.7%) (Fig. 6C). A prominent splenic TCRβ+ T-cell peak was also detected in nude mouse H5 (4.7%) and H6 (2.4%), albeit at a much lower percentage than control Balb/c levels (29%) (Fig. 6D).
Discussion
The MC model of in vivo tissue engineering creates a vascularized chamber to promote angiogenesis and de novo tissue formation. It is a relatively technically simple procedure, involving subcutaneous implantation of a split silicone chamber around the epigastric vessels, which can then be filled with an extracellular matrix scaffold, such as Matrigel, and growth factors, like FGF-2, to enhance angiogenesis of the engrafted tissue and provide a growth-conducive environment. We demonstrate a novel application of the MC model in supporting growth of embryonic thymus tissue for the potential study of T-cell–mediated immunity and other related applications.
The MC supported growth of fetal murine thymus grafts at comparable levels to conventional thymus-KC grafts, resulting in vascularized, lymphocyte-filled tissue of typical thymus organization, sustained over a period of 11 weeks. Increased peripheral T-cell production was observed from 2 weeks postectopic thymus engraftment, with comparable CD4 and CD8 mature T-cell subset distribution, indicating that normal thymopoiesis was occurring at both graft sites. The embryonic thymus does contain a small population of immature T cells; upon engraftment, these cells may continue to mature in the thymus graft and contribute to the peripheral T-cell pool. To sustain thymopoiesis, however, continual replenishment of the thymus graft with host-derived, blood-borne T-cell progenitors is required. The vascularized and well-formed thymus found in the MC upon harvest after 11 weeks indicates that indeed sustained thymopoiesis had occurred. Development of the thymus microenvironment from the primary cortical phenotype of the embryonic thymus,8,20 to exhibit large medullary regions, is further evidence of thymus maturation and the ability to select functional T cells, capable of mediating allogeneic skin graft rejection as demonstrated.
Although the MC model sustained function of murine fetal thymus tissue, it supported only limited growth of adult murine thymus. Thymic fragments were found in two adult thymus samples after a period of 12 weeks; however, they appeared disorganized and were often avascular, suggesting that parts of the adult thymus tissue had either failed to vascularize or were not sustained. Flow cytometric analysis revealed that the progression to mature SP thymocytes, through intermittent DP phenotype, was evident in only one adult thymic-MC graft (A4); however, increased DN and reduced DP proportion further suggest inefficient thymopoiesis and limited survival of the adult thymus lobe. The increased percentage of αβTCR+ T cells found within the spleen of mouse A4, however, does suggest an initial export of thymic-derived T cells from the MC-adult murine thymus graft and may correlate with the limited vascularization, observed by histology, in some adult thymic fragments. The absence of naive T cells within this peripheral T-cell population may be explained by their rapid expansion into memory/activated phenotype as a result of homeostatic pressures associated with the lymphopenic environment of a nude mouse, and the fact that thymopoiesis was not sustained.
The reduced ability of the adult thymus tissue to survive in the MC compared with its younger, embryonic, counterpart is not surprising given the inherent decrease in plasticity associated with aging tissue. Reduced proliferative capacity of thymic epithelium is evident with age4,20,21 and may interfere with the ability to sustain tissue at an ectopic graft site. Factors that may influence survival of the much larger, adult thymic lobes are reduced access to supplemented media and oxygen, resulting in hypoxia and loss of 3D structure. 22 A study of chamber hypoxic levels in the rat indicated that more than 7 days is required for adequate, angiogenic-dependent oxidation within the chamber microenvironment. 23 It is likely that delayed vascularization of larger adult lobes within the MC impinges on survival of the thymus tissue resulting in limited and inefficient thymopoiesis. Significant necrosis also occurs alongside hypoxia and may explain the avascular, adult thymic fragments found in MC A1 and A2, remnant after 12 weeks. Enhanced survival of adult murine thymus tissue may occur if the tissue is first cut into smaller pieces and then placed or sutured into the MC, within close proximity of the flow-through pedicle, to allow rapid and more thorough vascularization of the thymus tissue. In addition, supplementation of MC matrix with other thymic growth factors, such as keratinocyte growth factor, may increase the chances of thymic tissue survival. 24
Although only one human thymus tissue graft was sustained (n = 7), the finding was interesting, in that full development of murine-derived T cells, progressing through defined intermediate thymocyte subsets, occurred within this human thymus-MC graft and a small, but detectable, naive T-cell population could be found within the peripheral T-cell pool of this mouse. Although studies have reported a degree of success in the development of T cells via cross-species interaction, 5 current data suggest that proper development and self-tolerance of T cells require complex interaction with MHC I- and II-expressing thymic cortical and medullary epithelium of the same species, to ensure efficient positive and negative selection. 25 In this study, the MHC restriction of murine T cells, which developed in the human thymus, remains unresolved, but possible interaction with both human stromal elements and murine-derived antigen-presenting cells are the subject of ongoing studies. 26
Overall, the data suggest that the MC model has significant potential in sustaining ectopic thymus tissue grafts, particularly fetal thymus, and may provide a beneficial alternative, compared with renal capsule grafting, for the study of thymopoiesis and T-cell–mediated immunity in small animal models. The chamber is constructed from a silicone material, penetrable via percutaneous needle. Therefore, it would be possible to modulate the environment of an established MC-thymus graft via injection of cells or substances, for example, hemopoietic stem-cell subsets or HIV. In addition, the chamber offers a prototype for a new model of cell and tissue delivery in humans, where the KC would be considered generally unacceptable due to the invasive nature and the inappropriateness of repeated access. The subcutaneous chamber provides an isolated, yet readily accessible chamber with an independent blood supply, suitable for harvest, retrieval, manipulation, and retransplantation if necessary, without animal sacrifice nor donor site morbidity, given the redundancy of vasculature in this region. Depot drug and hormone implants are a well-tolerated and effective treatment modality in humans and would be akin to the chamber described.
In summary, although sustained growth of fetal thymus and functional T-cell production were obtained in this study, the successful growth of postnatal thymic tissue, however, is still relatively low in the MC model. Further manipulation of the MC microenvironment, including more rapid tissue vascularization, supplementation of matrix with thymic-specific growth factors, and addition of T-cell progenitors, may further enhance the survival of postnatal thymus tissue and T-cell genesis.
Footnotes
Acknowledgments
This research was supported by grants from the Australian National Health and Medical Research Council and the Australian Stem Cell Centre. The authors would like to thank Jason Palmer for his microscopic analysis.
Disclosure Statement
R.L.B. is the Chief Scientific Officer of Norwood Immunology Ltd.