
Abstract
Hematopoietic stem cells (HSCs) are commonly used in clinical transplantation protocols to treat a variety of diseases. However, efficient transplantation requires a substantial amount of HSCs from different sources and may require expansion. Therefore, effective expansion of HSCs remains a technical hurdle blocking the development of advanced cell therapies. The product of the human homeobox B4 (HOXB4) gene was recently demonstrated to effectively expand HSCs from umbilical cord blood (UCB) or bone marrow in either a retroviral or recombinant protein form. Our study purified TAT-HOXB4 proteins and demonstrated their ability to expand UCB and peripheral blood (PB) progenitor cells. The results showed that the TAT-HOXB4 gene product expanded the CD34+ progenitor cells from UCB and PB by ∼7.5-fold. The results from a semisolid cloning assay, a human long-term culture-initiating cell assay, and a nonobese diabetic–severe combined immunodeficiency mice repopulating assay showed that TAT-HOXB4 expanded hematopoietic progenitor cells while retaining their repopulating capacity and multipotency. TAT-HOXB4 protein also expanded engrafted stem cells that were previously expanded in a secondary transplantation assay. The results demonstrated the feasibility of using TAT-HOXB4 to expand UCB and PB progenitor cells, which are readily available to treat different hematological malignancies and nonhematological diseases.
Introduction
Recent evidence suggests that HOX transcription factors are involved in the regulation of normal and leukemic hematopoiesis. 12 Among the HOX genes, HOXB4 is selectively expressed in primitive hematopoietic cells but is downregulated in committed progenitors and more mature hematopoietic cells. 13 Retroviral overexpression of HOXB4 enhances HSC self-renewal and induces a net 40-fold increase in HSCs from mouse BM ex vivo. 9 In addition, the HOXB4-expanded HSCs retained their normal differentiation and long-term repopulation potential, and no hematologic abnormalities were detected in large groups of mice receiving transplants.9,14 Recent results showed that coculture with BM stromal cells genetically engineered to secrete HOXB4 or incubation of recombinant TAT-HOXB4 with either UCB or mouse BM stem cells expanded the stem cell pools 2.5- or 6-fold, respectively.15,16 Recombinant TAT-HOXB4 protein treatment may be useful in stem cell expansion for its simplicity and for its lack of involvement of stromal cells. However, the scale of recombinant TAT-HOXB4 proteins produced was too small to be used in a clinical setting. 16
Our study mass-produced and purified the recombinant TAT-HOXB4 proteins to a clinical scale. The Lin−CD34+ hematopoietic progenitor cells (HPCs) from UCB and PB were isolated using a CliniMacs method and cultured with the purified TAT-HOXB4 proteins. In vitro colony-forming cell (CFC) and long-term culture-initiating cell (LTC-IC) assays showed the multipotentiality of expanded HPCs. The expanded cells showed their nonobese diabetic–severe combined immunodeficiency (NOD-SCID) repopulation capacity after primary transplantation into NOD-SCID mice. Moreover, the CD34+ cells from transplanted NOD-SCID mice also showed CFU and SRC activity after secondary expansion and transplantation, indicating the long-term repopulating capacity of the TAT-HOXB4–expanded Lin−CD34+ HPCs from UCB and PB. The results set the stage for embarking on a phase I clinical trial using recombinant TAT-HOXB4 to expand HPCs from UCB or PB to treat different kinds of hematological malignancies and nonmalignant diseases.
Materials and Methods
TAT-HOXB4 protein purification and one-step desalting/renaturation protocol
The pTAT-HA-HOXB4 plasmid (a gift from Guy Sauvageau, University of Montreal, Montreal, Canada) was transformed into Bl21(DE3)pLysS (Novagen, Darmstadt, Germany), and the purification of TAT-HOXB4 was performed as previously described.
16
Briefly, cell lysates were suspended in buffer A (8 M urea, 20 mM HEPES [pH 8.0], and 100 mM NaCl), clarified by centrifugation (20,000 rpm for 30 min at 20°C), adjusted to 10 mM imidazole, and loaded on HisTrap chelating columns. Bound proteins were washed with 50, 100, and 250 mM imidazole in buffer A and eluted with 500 mM imidazole in buffer A. The fractions containing TAT-HOXB4 were loaded on a MonoSP column in buffer A without imidazole, eluted with 1 M NaCl and 20 mM HEPES (pH 8.0), and desalted using a one-step desalting/renaturation protocol. Briefly, the fraction containing denatured TAT-HOXB4 protein was mixed with
TAT-HOXB4 transduction ability
TAT-HOXB4 transduction, preparation of nuclear extracts, and western blot analysis were performed as previously described. 16
Electrophoretic mobility shift assay
Electrophoretic mobility shift assay was performed as previously described. 17 Purified TAT-HOXB4 protein (50 ng) was incubated with a [γ-32P]ATP end-labeled oligonucleotide (5′-CTGCGATGATTGATGACCGC-3′) containing the HOXB4-binding consensus sequence 18 and then loaded on 6% nondenaturing polyacrylamide gel. For competition assays, the protein was preincubated with 1–50-fold molar excess of unlabeled oligonucleotide.
RNA purification and quantitative real-time polymerase chain reaction analysis
The procedures were performed to evaluate the mRNA expression levels of Bambi and cdk6 in cultured cells, as previously described.17,19,20
TAT-GFP plasmid construction and TAT-GFP protein purification
The pTAT-HA-GFP plasmid was generated by inserting a 711-bp fragment of the GFP cDNA from pEGFP (BD Bioscience, San Jose, CA) into the pTAT-HA expression vector. The expression, purification, and desalting/renaturation of TAT-GFP was performed as described in the purification of TAT-HOXB4.
Endotoxin depletion
Endotoxin was removed by Detoxi-Gel™ Endotoxin Removing Gel (Thermo Scientific, Rockford, IL). The preparations were tested endotoxin free by the Limuslus Amebocyte Lysate QCL-1000 endotoxin detection kit (Cabrex Bioscience, Walkersville, MD).
Purification of HSCs, TAT-HOXB4 transduction, and expansion of HSCs
UCB samples were obtained from Taiwan Advance Bio-Pham Inc. (Taipei County, Taiwan), and PB samples were obtained from Taipei Veterans General Hospital (Taipei, Taiwan). Informed consent was obtained before procurement of UCB or PB samples. This study was approved by the Taipei Veterans General Hospital Institutional Review Board in 2005. All the frozen blood samples were thawed in 37°C water bath. Mononuclear cells (MNCs) were isolated using Ficoll-Paque (GE Health Care, Uppsala, Sweden) density gradient centrifugation. The red blood cells of the treated MNCs were removed by lysis at 4°C in 0.83% ammonium chloride with 0.1% sodium bicarbonate (pH 7.0). The CD34+ cells were purified using the CliniMacs procedure (MiltenyiBiotec, Auburn, CA). The purified CD34+ progenitor cells were enriched by negative depletion of lineage-positive cells with nine different markers, including CD2, CD3, CD14, CD16, CD19, CD24, CD56, CD66b, and glycophorin A, using a StemSep device (StemCell Technologies, Vancouver, Canada). The purified cells were cultured for 2 days in stem cell medium (Dulbecco's modified Eagle's medium (DMEM) with 10% fetal calf serum [FCS], 5 ng/mL interleukin-3, 10 ng/mL interleukin-6, 100 ng/mL stem cell factor) and then for 4 days in stem cell medium containing 15 nM HOXB4, 15 nM TAT-GFP, or 1% BSA. On day 3 (day 0 of treatment), 4 × 105 cells/mL was resuspended in stem cell medium supplemented with BSA, TAT-GFP, or HOXB4. Fresh BSA, TAT-GFP, or TAT-HOXB4 proteins (50% initial protein amount in 5% of total culture medium) were then added every 3 h as previously described. 16 After a 12-h period, FCS and cytokines were added to correct for the resulting 20% dilution of culture media. After 24 h, cells were resuspended in fresh medium containing BSA, TAT-GFP, or TAT-HOXB4. On days 0, 2, and 4 of treatment, CD34+ cells were counted using ISHAGE procedure (CD45/CD34 staining plus side and forward scatter evaluation).
Semisolid cloning assays
Human CFCs were assayed by plating cells in 1 mL standard methylcellulose (StemCell Technologies, Vancouver, Canada) supplemented with 1% methylcellulose (4000 cps), 30% FCS, 1% BSA, 0.1 mM 2-mecraptoethanol, 2 mM
LTC-IC assays
LTC-IC assays using human hematopoietic cells (1000 cells/cm2) and preirradiated (30 Gy using a 137Cs-γ irradiator) MS-5 cells (3 × 104 cells/cm2) as feeder layer were performed in a long-term culture medium (DMEM supplemented with 12.5% FCS, 12.5% horse serum, and 10−4 M β-mercaptoethanol) using limiting dilution. Cells from cultures were counted every week and cloned for CFC assay as previously described.15,16 Total LTC-IC number was obtained by multiplying the frequency of LTC-ICs, as determined in secondary cultures, by the total number of cells present after the 5-week primary long-term culture.
NOD-SCID mice repopulating cells (SRCs) assays
An SRC assay was performed using the methods previously described with slight modification. 16 All the mice were handled in sterile conditions and maintained in germ-free isolators located in the Taiwan GlycoNex Inc. (Taipei County, Taiwan). A total of 104 initial cells were diluted in a limiting dilution series and then expanded by BSA or TAT-HOXB4. The expanded cells were then collected and injected into 4–6-week-old irradiated (2.5 Gy using a 137Cs-γ irradiator) NOD-LtSz-scid/scid (NOD-SCID) mice along with 105 CD34-irradiated (30 Gy using a 137Cs-γ irradiator) accessory cells. Fourteen to 16 weeks after transplantation, the percentages of human CD45+ cells in the murine BM cells were analyzed by flow cytometry. Mice were scored as positive if more than 0.1% of total murine BM cells were CD45+.
Analysis of human cell engraftment in NOD-SCID mice by flow cytometry
The repopulated human HPCs in murine BMs was determined as previously described.21,22 Briefly, repopulated cells were determined by detecting the number of cells positively stained with a fluorescein isothiocyanate-conjugated anti-human CD45 monoclonal antibody (Becton Dickinson, San Jose, CA). The cells were also stained with a phycoerythrin (PE)-conjugated anti-human CD34 monoclonal antibody (Becton Dickinson, San Jose, CA) and Allophycocyanin (APC)-conjugated monoclonal antibody for human lineage-specific antigens, including CD19-PE and CD14/15-PE (both from Becton Dickinson), to detect specific subsets of human hematopoietic cells. BM cells were suspended in Ca2+- and Mg2+-free PBS containing 2% FCS after lysis of red blood cells. The cells were then incubated with human immunoglobulin G, followed by staining with the aforementioned monoclonal antibodies.
Secondary transplantation
Secondary transplantation was performed as previously described. 23 Briefly, murine BM cells were obtained from the tibiae and femurs of highly engrafted primary recipient mice 14–16 weeks after transplantation. BM cells were purified and expanded as described earlier. The expanded cells were subsequently transplanted into irradiated secondary recipient mice. Fourteen weeks after transplantation, the presence of human CD45+ cells in the secondary recipient mice was analyzed by flow cytometry as described in primary transplantation.
A new serum-free medium for expansion of HSCs
A new serum-free medium containing TAT-HOXB4 to support in vitro expansion of HSCs was developed, and the recipe was constituted of DMEM with 5% recombinant human serum albumin, 5 ng/mL interleukin-3, 10 ng/mL interleukin-6, 100 ng/mL stem cell factor, 30 ng/mL thrombopoietin (TPO), 50 ng/mL Flt-3 ligand, and antibiotics (penicillin/streptomycin). The serum-free medium containing 15 nM HOXB4 or 1% BSA was used in experiments.
Statistical analysis
All data were analyzed with SPSS 13 software (SPSS, Chicago, IL). The results were presented as means with standard errors. Statistical significances were declared at p < 0.05.
Results
Generation of clinically scaled and biologically active TAT-HOXB4
Our study used a recombinant TAT-HOXB4 fusion protein as described to expand HSCs in vitro through direct protein delivery. 16 For clinical applications, a one-step desalting/renaturation protocol was used to purify the mass-produced TAT-HOXB4 proteins (see Materials and Methods section). The scale of TAT-HOXB4 production after purification consistently reached ∼1.5 mg/L of bacterial culture after the one-step desalting/renaturation protocol. To verify that the biochemical property of TAT-HOXB4 was not changed, 293T cells were incubated with TAT-HOXB4 proteins to measure its ability to translocate into the nucleus. Figure 1A showed that TAT-HOXB4 retained this nuclear translocation ability. To demonstrate that TAT-HOXB4 is biologically active, the electrophoretic mobility shift assay was used to test its DNA-binding activity. The results showed that TAT-HOXB4 was able to bind the HOXB4-binding consensus sequence 18 ; this binding could be competed away by an excess amount of unlabeled oligonucleotides (Fig. 1B). TAT-HOXB4 was also incubated with HPCs to test its ability to activate downstream targets. Figure 1C shows that two of the HOXB4 downstream targets, Bambi and cdk6,19,20 were activated after the cells were incubated with TAT-HOXB4, using a quantitative real-time polymerase chain reaction analysis. These results showed that the purified recombinant TAT-HOXB4 proteins are biologically active, and the biochemical properties of purified TAT-HOXB4 resembled those of the original TAT-HOXB4 previously described. 16 The range of TAT-HOXB4 concentrations required for HSCs expansion was tested, and the optimal concentration required to expand both total MNCs and CD34+ cells is 15 nM (data not shown).
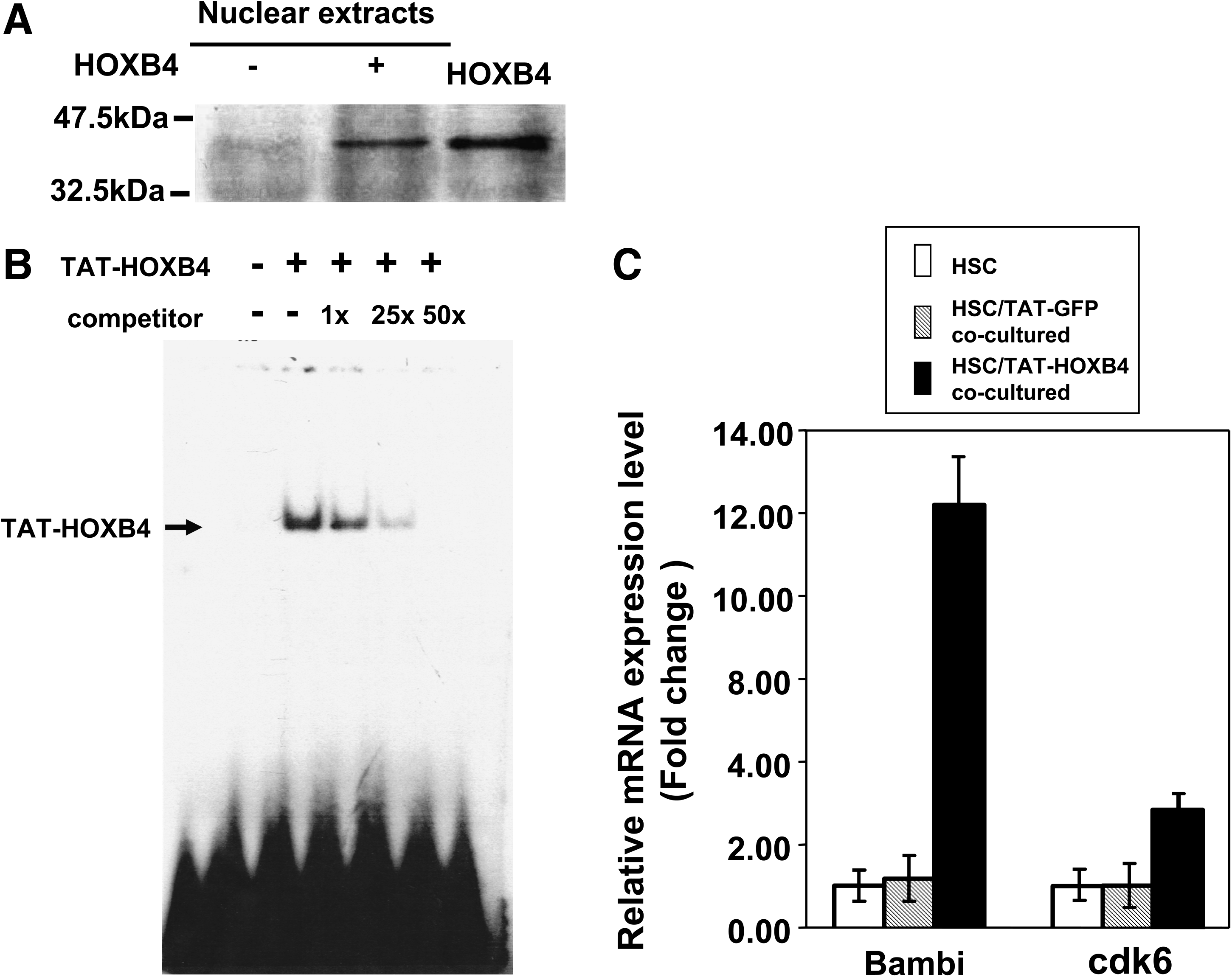
Nuclear translocation of TAT-HOXB4 proteins and demonstration of the biological activity of TAT-HOXB4. (
TAT-HOXB4 expanded hematopoietic progenitors of UCB and PB in vitro
UCB progenitor cells were used to test the expansion capability of TAT-HOXB4. After MNCs were incubated with TAT-HOXB4 (at the concentration of 15 nM), their numbers increased ∼2.5-fold compared with the BSA control (Fig. 2A). The total Lin−CD34+ cells from UCB were purified using the ClinMacs/StemSep method (see Materials and Methods section) to test whether they can be expanded by TAT-HOXB4. The percentage of CD34+ cells in UCB was found to be ∼1% before purification and ∼99% after purification. The number of total CD34+ cells increased ∼7.4-fold when compared with the BSA control after incubation with TAT-HOXB4 for 4 days (Fig. 2B). Similar procedures were performed on the Lin−CD34+ cells from PB. The number of MNCs increased ∼2.3-fold compared with the BSA control (Fig. 2A). The percentage of CD34+ cells in PB was found to be ∼1.2% before purification and ∼99% after purification. TAT-HOXB4 also expanded the PB CD34+ cells ∼7.6-fold compared with the BSA control (Fig. 2B). These results showed that recombinant TAT-HOXB4 was able to expand CD34+ cells in both UCB- and PB-derived cells in vitro. To assess whether TAT-HOXB4 can amplify human hematopoietic progenitors, assays of CFCs and LTC-ICs were performed after the progenitors were incubated with TAT-HOXB4 or BSA for 4 days. Figure 2C shows that total CFCs increased ∼10.6-fold in UCB and ∼9.5-fold in PB progenitor cells. Figure 2D shows that total LTC-ICs increased ∼12.8-fold in UCB and ∼9.5-fold in PB progenitor cells. These results indicated that TAT-HOXB4 expanded CD34+ cells and effectively amplified progenitor cells from UCB and PB. To exclude the possibility that the recombinant proteins were contaminated with endotoxin, the same procedures were used to purify recombinant TAT-GFP to serve as a negative control. Using TAT-GFP versus TAT-HOXB4 to expand UCB and PB progenitors, a similar degree of expansion was achieved by TAT-HOXB4 but not by TAT-GFP (Supplemental Fig. S1), available online at

Expansion of UCB and PB stem cells by recombinant TAT-HOXB4. (
TAT-HOXB4 amplifies NOD-SCID mice repopulating cells (SRCs) which retains multipotency
SRCs are considered to be the most primitive hematopoietic cells experimentally identifiable in humans.21,22 To quantitatively test the effects of TAT-HOXB4 protein on SRCs, our study employed limiting-dilution experiments to determine the frequency and repopulating activity of CD34+ UCB and PB treated with TAT-HOXB4 or BSA for 4 days. The SRC frequency increased from 1 in 20,000 (range defined by ±1 standard errors (s.e.): 16,000–25,000; BSA) to 1 in 1600 (range defined by ±1 s.e.: 1100–2600; TAT-HOXB4) CD34+ cells in UCB (Fig. 3A) and from 1 in 21,000 (range defined by ±1 s.e.: 18,500–27,000; BSA) to 1 in 2100 (range defined by ±1 s.e.: 1500–2300; TAT-HOXB4) in PB (Fig. 3B). These results indicated that TAT-HOXB4 expanded the repopulating cells by 12.5-fold (UCB) and 10-fold (PB), respectively, over the CD34+ cells treated with BSA. After 4 days of treatment with TAT-HOXB4 versus BSA, SRC-derived human cells were found in 84% of mice injected with the cell progeny of 1000 cells under TAT-HOXB4 expansion compared with 8% of mice injected with the same number of cell progeny in cultures treated with BSA (Table 1). Similar experiments were performed using CD34+ PB cells, and the difference in the percentage of engraftment is also significant (80% for TAT-HOXB4 vs. 8% for BSA; Table 1). The ability of TAT-HOXB4 to increase the repopulating capacity of human HSCs was shown by the significant enhancement of the level of chimerism in mice transplanted with CD34+ cells treated with TAT-HOX4 versus BSA (6.85% for TAT-HOXB4 vs. 0.28% for BSA in UCB and 6.18% for TAT-HOXB4 vs. 0.45% for BSA in PB; Fig. 3C). Finally, phenotypic analysis showed that TAT-HOXB4–induced expansion of UCB and PB did not alter the differentiation program of hematopoietic progenitors in vivo. The engrafted cells from expanded UCB and PB were able to differentiate into different lineages such as myeloid progenitor (CD14/CD15), B-lymphoid (CD19), and mature myeloid (CD45) cells (Fig. 3D and data not shown), demonstrating their ability to differentiate into cells from different lineages.
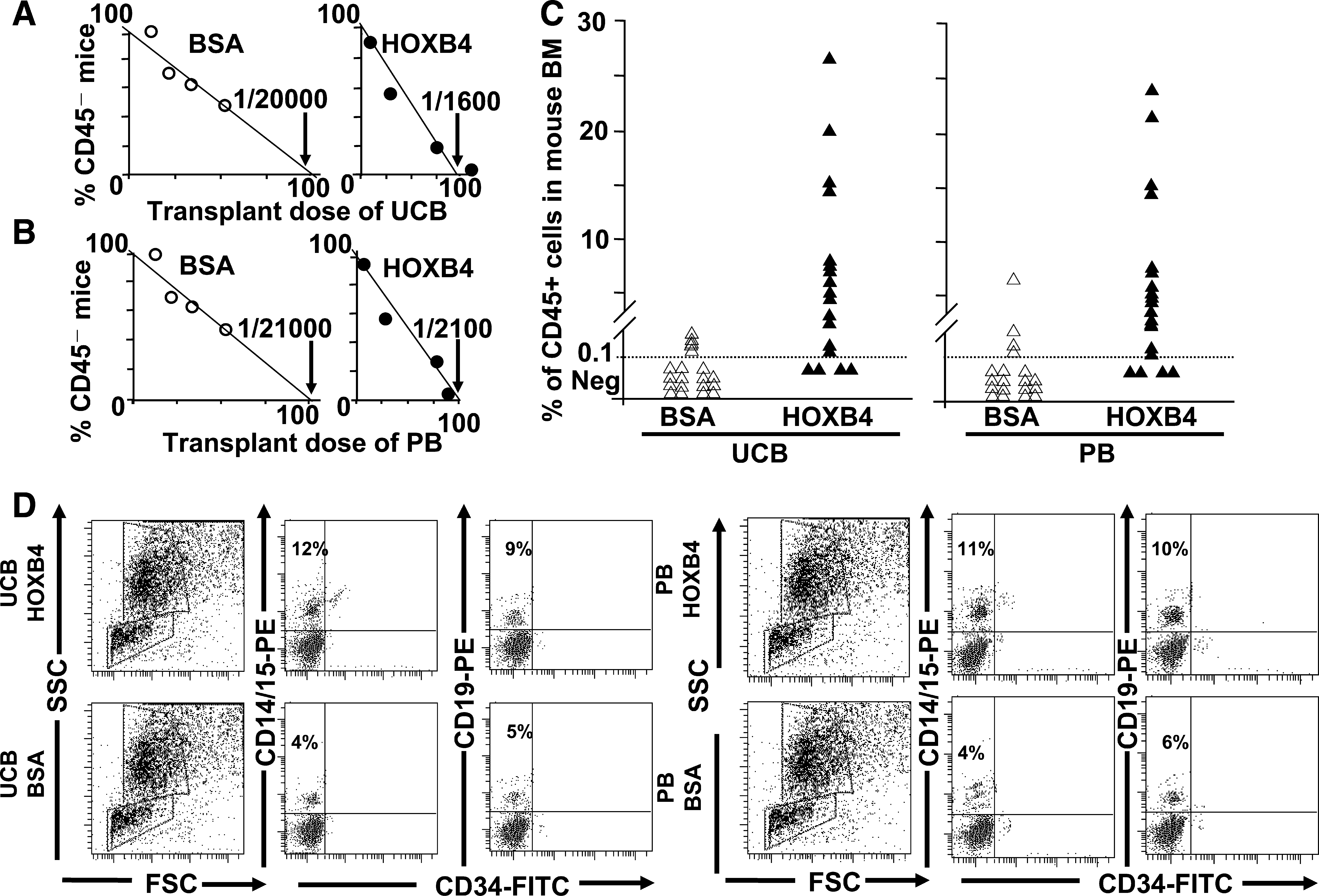
In vitro expansion of SRCs by TAT-HOXB4. (
Initial cell number was 1500.
BSA, bovine serum albumin; HOXB4, homeobox B4.
Expansion of CD34+ progenitor cells by TAT-HOXB4 in secondary transplantation
To further evaluate the long-term repopulating potential of expanded SRCs, BM cells obtained from each primary recipient mouse were assessed for their SRC activity by secondary expansion and transplantation (see Materials and Methods section). Fourteen weeks after primary transplantation, the transplanted marrow contained CD34+ cells (∼0.87%; Fig. 4A). Our study purified these CD34+ BM cells and incubated them with 15 nM TAT-HOXB4. The number of CD34+ cells increased approximately sixfold (Fig. 4B). In addition, the total CFCs and total LTC-ICs were expanded ∼7.2-fold and ∼12.5-fold, respectively, by TAT-HOXB4 (Fig. 4C, D). The SRC frequency increased from 1 in 21,000 (range defined by ±1 s.e.: 17,500–26,000; BSA) to 1 in 2500 (range defined by ±1 s.e.: 2100–2900; TAT-HOXB4) CD34+ cells in secondary-transplanted UCB (Fig. 4E). The successful rate of engraftment in the secondary transplantation was 80% for TAT-HOXB4 versus 8% for BSA (Table 2). The ability of TAT-HOXB4 to increase the repopulating capacity of primary-transplanted SRCs was shown by the significant enhancement of the level of chimerism in mice transplanted with CD34+ cells treated with TAT-HOX4 versus BSA (6.35% for TAT-HOXB4 vs. 0.43% for BSA) (Fig. 4F). Phenotypic analysis also showed that TAT-HOXB4 did not alter the differentiation program of secondary-transplanted hematopoietic progenitors in vivo. The expanded progenitor cells from the marrow of primary-transplanted mice were able to differentiate into cells containing all the expected CD markers such as myeloid progenitor (CD14/CD15), B-lymphoid (CD19), and mature myeloid (CD45) cells (Fig. 4G and data not shown). All these results demonstrated that TAT-HOXB4 was able to expand the primary-transplanted stem cells to facilitate efficient secondary engraftment without affecting their differentiation potential.
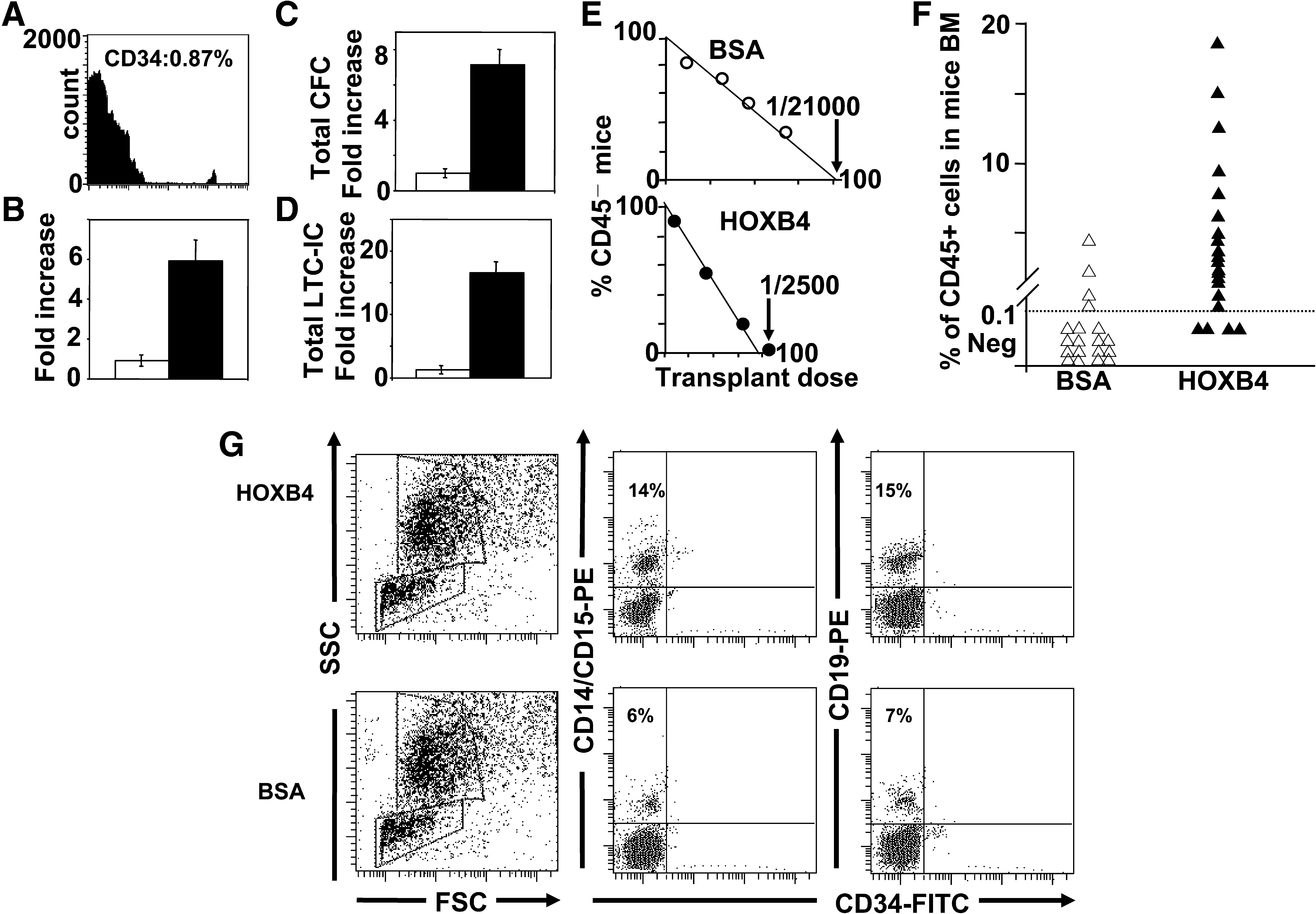
TAT-HOXB4 expanded CD34+ cells from the BM of primary-transplanted mice, which were secondary-transplanted into NOD-SCID mice. (
Initial cell number was 1500.
Expansion by TAT-HOXB4 using a serum-free medium
To facilitate clinical application, a new serum-free medium containing TAT-HOXB4 to support in vitro expansion of HSCs was developed. The recipe of the serum-free medium has been described earlier (see Materials and Methods section). To evaluate the feasibility of expanding HSCs using a serum-free medium that contains TAT-HOXB4, UCB cells were incubated with 15 nM TAT-HOXB4 in a serum-free medium. The number of total MNCs increased approximately threefold and CD34+ cells increased approximately sixfold (Fig. 5A, B). The total CFCs and total LTC-ICs were expanded ∼7-fold and ∼15.1-fold (Fig. 5C, D), respectively. The SRC frequency increased from 1 in 22,000 (range defined by ±s.e.: 17,500–26,000; BSA) to 1 in 1800 (range defined by ±1 s.e.: 2100–2900; TAT-HOXB4) (Fig. 5E). Phenotypic analysis also showed that the HSCs expanded by serum-free medium retained multipotency ex vivo. The expanded progenitor cells from the marrow of transplanted mice were able to differentiate into cells containing all the CD markers analyzed in our study, such as myeloid progenitor (CD14/CD15), B-lymphoid (CD19), and mature myeloid (CD45) (Fig. 5F and data not shown). All these results demonstrated that TAT-HOXB4 was able to expand the HSCs in a serum-free medium.
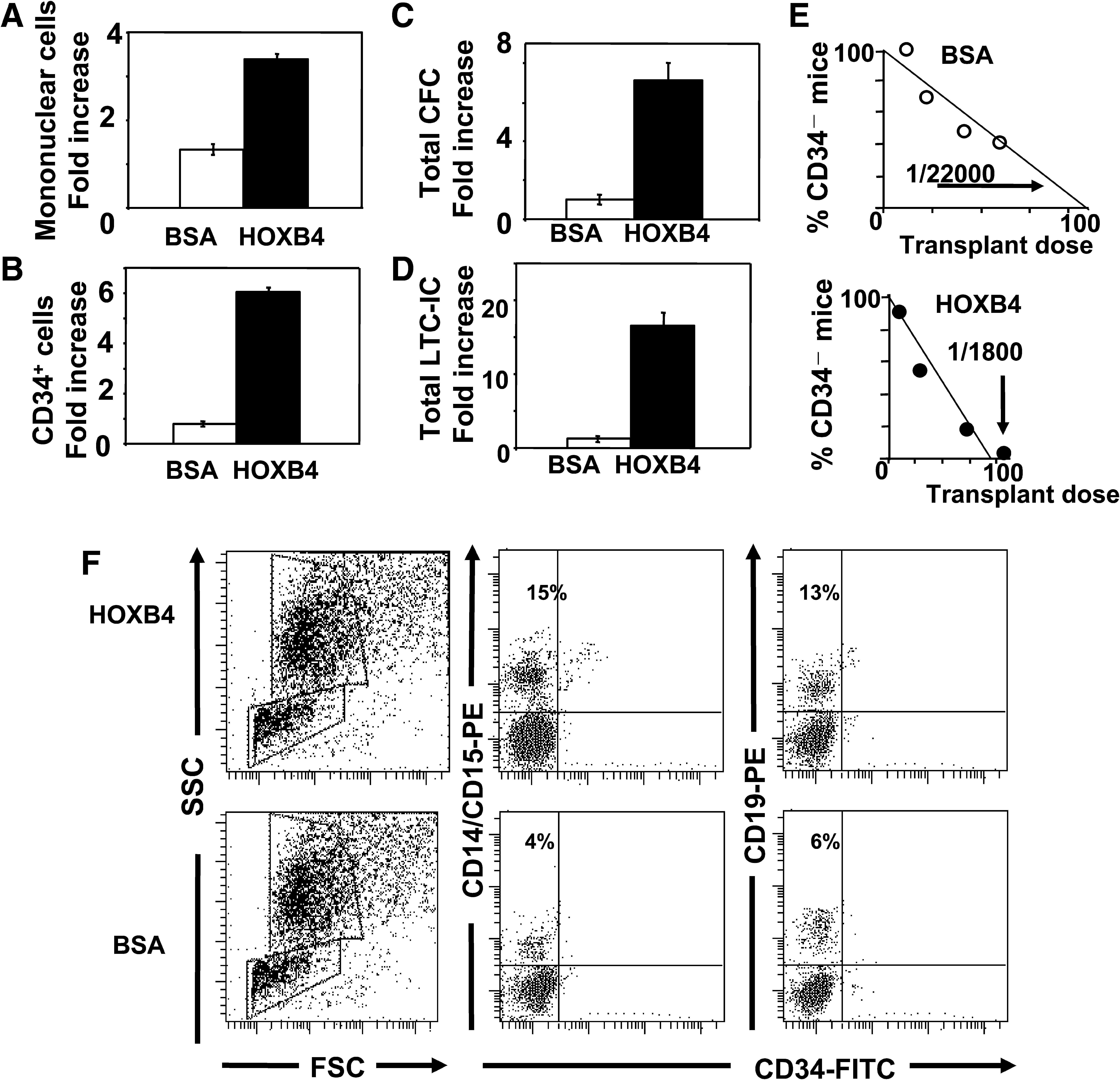
TAT-HOXB4 in serum-free medium expanded CD34+ cells from UCB, which were transplanted into NOD-SCID mice. (
Discussion
The technical hurdles involved in stem cell expansion are high, and efficient methods of expansion have not yet been established.4–6 HOXB4 was recently shown to be one of the promising molecules capable of expanding stem cells in vitro and ex vivo.7,16,24 However, clinical trials have not been performed because of the unreliability of delivery methods (e.g., retroviral vectors) and limitation of the amount of recombinant proteins to be used.15,16 This report demonstrated a technical advancement designed to overcome the difficulties encountered for the application of HOXB4 in a clinical setting using a one-step desalting/renaturation method to rapidly purify native proteins. The routine usage of the CliniMacs/StemSep procedures to purify large numbers of hematopoietic progenitors from UCB and PB was also applied. In addition, using these mass-produced TAT-HOXB4 proteins, our study demonstrated that (1) TAT-HOXB4 protein was capable of expanding stem cells from UCB and PB as measured by the fold of increase in CD34+ counts, total number of CFCs, total number of LTC-ICs, and significantly increased engraftment ratio in NOD-SCID mice; (2) the transplanted human stem cells could be reharvested from mouse marrow and re-expanded by TAT-HOXB4 to achieve successful secondary engraftment. All these results demonstrated that our practical approaches of using TAT-HOXB4 to expand stem cells have set up the stage for clinical trial in the immediate future.
The original TAT-HOXB4 generated from the traditional column purification methods has a lower yield (0.7 mg/L of bacteria culture) than the methods presented in our study. The one-step desalting/renaturation protocol also provided an easier and efficient approach to purify recombinant TAT-HOXB4 en mass. Recently, similar results using recombinant TAT-HOXB4 to expand UCB and PB progenitor cells were reported. 25 However, our study is the first to successfully perform secondary transplantation, suggesting the feasibility of using TAT-HOXB4 in a clinical setting compared with the unreliable secondary transplantation previously reported. 15 Preliminary results also showed that expansion of the secondary-engrafted marrow after harvesting of human CD34+ cells was successful, showing the reasonable rate of increase in CD34+ cells, the total number of CFCs, and the total number of LTC-ICs (data not shown). All these results strongly suggested that TAT-HOXB4 may be the first effective reagent capable of expanding stem cells reproductively.
Retrovirally transduced HOXB4 have been used to expand stem cells with some success. 26 However, it is difficult to adjust the expression levels of HOXB4 because the expression driven by retroviral vectors would be subject to the control of integration sites and the strength of promoters. Stem cells expanded with retrovirally transduced HOXB4 also resulted in high incidence of leukemia. 27 High levels of HOXB4 were also shown to inhibit certain lineage differentiation, 28 which will severely hamper the outcome of transplantation. Using recombinant TAT-HOXB4 would allow us to adjust the dosage of HOXB4 to be delivered inside cells to achieve appropriate lineage differentiation. In addition, the safety of retroviral vectors would preclude the clinical application compared with the usage of recombinant TAT-HOXB4 proteins, whose existence inside cells are transient (within the span of 4 days) and can be regulated by the dosage delivered. Similarly, delivery of recombinant rPTD-HOXB4 into human embryonic stem cells has been shown to promote hematopoietic development of human embryonic stem (hES) cells. 29 However, the CD34+ cells derived from these hES cells did not have increased engraftment capacity when they were exposed to rPTD-HOXB4, in contrast to our direct application of TAT-HOXB4 to UCB and PB progenitor cells which could be readily applied to a clinical setting. Recently, an automated system for delivery of TAT-HOXB4 was developed. 30 This automated system would definitely facilitate our protocol to expand UCB and PB progenitor cells for use in a clinical setting.
In conclusion, our study successfully developed a new purification protocol to mass-produce recombinant TAT-HOXB4 proteins for use in a clinical setting. The experimental results showed that TAT-HOXB4 was capable of expanding progenitor cells from UCB and PB as measured by all the assays employed herein. Our study therefore sets the stage for further clinical trials using TAT-HOXB4 to treat different kinds of hematological diseases.
Footnotes
Acknowledgments
The authors are grateful to Dr. G. Sauvageau for the gift of TAT-HOXB4 plasmid. This research was supported by Taiwan Advance BioPharm, Inc.
Disclaimer
The sponsor of the study had no role in study design, data collection, data analysis, data interpretation, or writing of the report. The authors had full access to all the data in the study and had final responsibility for the decision to submit for publication.
Disclosure Statement
C.H.H., T.C.L., and W.M.K. are employed by Taiwan Advance Biopharm, Inc. K.J.W. is a consultant of Taiwan Advance Biopharm, Inc., and also owns stocks of Taiwan Advance Biopharm, Inc. P.M.C., T.J.C., M.H.Y., and J.Y.K. do not have any relation with the Taiwan Advance Biopharm, Inc. No other competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.