
Abstract
There are only a few reports that describe the hepatocytic differentiation potential of human adipose tissue-derived mesenchymal stem cells (hADMSCs) and no reports that describe the in vivo functions of hepatocyte-like cells differentiated from somatic stem cells including hADMSCs. In this study, we established a new method for generation of functional hepatocyte-like cell clusters using floating culture method and induced functional hepatocyte-like cell clusters, which functioned effectively not only in vitro but also in vivo. The generated hepatocyte-like cell clusters were characterized by gene expression analysis, functional assays, and transplantation into non-obese diabetic severe combined immunodeficiency (NOD-SCID) mouse with chronic liver injury. The generated hepatocyte-like cell clusters expressed various genes normally found on mature hepatocytes. The cell clusters exhibited functional characteristics of hepatocytes: they expressed albumin, secreted urea, had cytochrome P450 activity, could take up low-density lipoprotein, and stored glycogen. Transplantation of these cell clusters into NOD-SCID mouse with chronic liver injury resulted in a significant improvement of serum albumin and total bilirubin levels. In summary, we established a new protocol for efficient induction of hADMSCs into functional hepatocyte-like cell clusters.
Introduction
Tissue engineering holds promise for the development of stem cell-based therapy of the liver. Many types of stem cells from different sources have been investigated for their ability to differentiate into hepatocytes. Recently, adult somatic stem cells, which can give rise to tissues of more than one germ layer, have been identified. 3 The current research focus is on mesenchymal stem cells (MSCs), which are found in human bone marrow (BM), 4 scalp tissue, 5 placenta, 6 and umbilical cord blood. 7 These stem cells can be induced into multiple-type lineages.8–12 MSCs from the BM and umbilical cord blood have been induced into a hepatic lineage13–17 ; however, it may also prove difficult to obtain sufficient amounts of autologous adult stem cells from these origins. Recently, MSCs have been isolated from adipose tissue.18,19 Adipose tissue is an attractive source of human MSCs because it is a safe and abundant source of large amounts of MSCs. Adipose tissue-derived MSCs (ADMSCs) exhibit differentiation potential similar to BM-derived MSCs. Nevertheless, there are a few reports describing the hepatocytic differentiation potential of ADMSCs20,21 and the effective functions of these differentiated hepatocyte-like cells not only in vitro but also in vivo.
In this study, we established a new method for generation of hepatocyte-like cell clusters, which functioned effectively not only in vitro but also in vivo.
Materials and Methods
Adipose tissues from human subjects
Excess adipose tissues were resected from the gastro-omental artery during coronary artery bypass graft surgery and gastrectomy in 10 subjects (four men and six women; age, 55 ± 5 years, mean ± standard error of the mean; range, 40–60 years). All subjects provided informed consent. The protocol was approved by the Review Board for Human Research of the Osaka University Graduate School of Medicine. All subjects fasted for at least 10 h before surgery and none was being treated with steroids. Ten to 50 g of adipose tissues was obtained from each subject.
Isolation of human ADMSCs
Human ADMSCs (hADMSCs) were obtained using the method described by Björntorp et al. 18 and Zuk et al., 19 with some modifications as described below.22,23 Resected excess adipose tissue was minced and then digested in Hank's balanced salt solution (HBSS; Gibco Invitrogen, Grand Island, NY) containing 0.075% collagenase type II (Sigma-Aldrich, St. Louis, MO) at 37°C for 1 h. Digests were filtered with a cell strainer (BD Bioscience, San Jose, CA) and centrifuged at 800 g for 10 min. Erythrocytes were excluded using density gradient centrifugation with Lymphoprep (d = 1.077; Nycomed, Oslo, Norway). The cells were then plated using Dulbecco's modified Eagle's medium (DMEM; Gibco Invitrogen) with 10% defined fetal bovine serum (FBS; Hyclone, Northumberland, United Kingdom) and incubated for 24 h at 37°C. Following incubation, the adherent cells were washed extensively and then treated with 0.2 g/L ethylenediaminetetraacetate (EDTA) solution (Nacalai Tesque, Kyoto, Japan). The resultant suspended cells were replated at a density of 10,000 cells/cm2 on human fibronectin (FN)-coated dishes (BD BioCoat) in medium I (60% DMEM-low glucose, 40% MCDB-201 medium [Sigma-Aldrich], 1 × insulin–transferrin–selenium [Gibco Invitrogen], 1 nM dexamethasone [Sigma-Aldrich], 100 μM ascorbic acid 2-phosphate [Sigma-Aldrich], 10 ng/mL epidermal growth factor [PeproTec, Rocky Hill, NJ], and 5% FBS).
Cultures of HepG2, normal skin fibroblasts, and primary hepatocytes
Both HepG2 (a hepatoblastoma cell line) and normal skin fibroblasts were cultured with DMEM containing 10% FBS and antibiotic/antimycotic solution. Primary hepatocytes (lot no. ACBRI3716) were purchased from Applied Cell Biology Research Institute (Kirkland, WA) and cultured using the method recommended by the manufacturer. Cells were maintained at 37°C.
Flow cytometric analysis
hADMSCs isolated from adipose tissue were characterized by flow cytometry. Cells were detached and stained with fluorescein isothiocyanate-conjugated mouse monoclonal antibodies to human CD31 (BD Pharmingen, San Diego, CA), CD105 (Ancell, Minneapolis, MN), CD133 (R&D Systems, Minneapolis, MN), and phycoerythrin-conjugated mouse monoclonal antibodies to human CD29, CD34, CD45, CD56, CD73, CD166 (BD Pharmingen), CD44, or CD166 (Ancell). Isotype-identical antibodies served as controls. Further, we incubated cells with mouse monoclonal antibodies against human stage-specific embryonic antigen-4 (SSEA-4), tumor rejection antigen (TRA)-1-60, tumor rejection antigen (TRA)-1-81 (Chemicon International, Temecula, CA), multidrug-resistance transport protein (ABCG-2; BD Pharmingen), or CD117 (BD Pharmingen), with nonspecific mouse antibody used as a negative control. After washing with Dulbecco's phosphate-buffered saline (PBS; Nacalai Tesque), cells were incubated with phycoerythrin-labeled goat anti-mouse immunoglobulin (Ig) antibody (BD Pharmingen) for 30 min at 4°C. After three washes, cells were resuspended in PBS and analyzed by flow cytometry using a FACSCalibur flow cytometer and CellQuest Pro software (BD Bioscience).
Adipogenic, osteogenic, and chondrogenic differentiation procedures
For adipogenic differentiation, cells were cultured in differentiation medium (Zen-Bio, Durham, NC). After 3 days, half of the medium was changed with adipocyte medium (Zen-Bio) and this was repeated every 2 days. Five days after differentiation, characterization of adipocytes was confirmed by microscopic observation of intracellular lipid droplets by oil red O staining. Osteogenic differentiation was induced by culturing the cells in DMEM containing 10 nM dexamethasone, 50 mg/dL ascorbic acid 2-phosphate, 10 mM β-glycerophosphate (Sigma, St. Louis, MO), and 10% FBS. Differentiation was examined by alizarin red staining. For chondrogenic differentiation, 2 × 105 cells of hADMSCs were centrifuged at 400 g for 10 min. The resulting pellets were cultured in chondrogenic medium (α-MEM supplemented with 10 ng/mL transforming growth factor-β, 10 nM dexamethasone, 100 μM ascorbate, and 10 μL/mL of 100 × insulin–transferrin–selenium solution) for 14 days as described previously. 24 For alcian blue staining, nuclear counterstaining with Weigert's hematoxylin was followed by 0.5% alcian blue 8GX for proteoglycan-rich cartilage matrix.
Differentiation procedure of hepatocyte-like cell clusters
Our differentiation procedure consisted of three stages. In stage I, hADMSCs were cultured and expanded in medium I for three to four passages. In stage II, the cells were dissociated with trypsin-EDTA and the resulting single cells were suspended in medium II (80% knockout-DMEM [Gibco Invitrogen], 20% defined FBS, 1 mM glutamine, and 1% nonessential amino acids [both from Gibco Invitrogen]). The suspension was placed in an ultralow-attachment culture dish (Hydrocell; purchased from CellSeed, Tokyo, Japan), and the cells self-aggregated into cell clusters within 1 day. The cell clusters were then cultured for additional 2 days. In stage III, after washing extensively with PBS, 2-day-old cell clusters (consisting of 1000 cells on average) were cultured on a Hydrocell dish for 4 weeks in medium III (60% DMEM-low glucose, 40% MCDB-201, 1 nM dexamethasone, 100 μM ascorbic acid, 10 ng/mL epidermal growth factor, basic fibroblast growth factor [Peprotech, Rocky Hill, NJ], hepatocyte growth factor [Peprotech], and oncostatin M). Finally, 0.1% dimethyl sulfoxide (DMSO; Nacalai Tesque) was added on the 10th day after induction of differentiation.
Hepatic differentiation procedure on extracellular matrix
After the third passage, hADMSCs were plated on human FN, collagen type I, or Matrigel-coated dishes in medium III. Further, 0.1% DMSO was also added on the 10th day after induction of differentiation.
Quantitative reverse transcription–polymerase chain reaction analysis and western blot analysis
Total RNA was isolated from cells using an RNAeasy kit (Qiagen, Valencia, CA). After treatment with DNase, cDNA was synthesized using Superscript III RNase H-minus Reverse Transcriptase (Invitrogen, Carlsbad, CA). Real-time polymerase chain reaction (PCR) was performed using the ABI Prism 7900 Sequence Detection System (Applied Biosystems, Foster City, CA). 20 × Assays-on-Demand™ Gene Expression Assay Mix for Albumin (ALB) (Hs00609411_m1), glutamine synthetase (Hs00374213_m1), cytochrome P450 (CYP)1B1 (Hs00164383_m1), cytokeratin 18 (CK18) (Hs01941416_g1), alpha fetoprotein (AFP) (Hs00173490_m1), CYP3A4 (Hs01546612_m1), CYP3A7 (Hs00426361_m1), alpha-1-antitrypsin (Hs01097800_m1), glucose-6 phosphatase (Hs00609178_m1), and glyceraldehyde-3-phosphate dehydrogenase (Hs99999905_m1) were obtained from Applied Biosystems. TaqMan® Universal PCR Master Mix, No AmpErase® UNG (2 ×) was also purchased from Applied Biosystems. Reactions were performed in triplicate and the mRNA levels were normalized relative to glyceraldehyde-3-phosphate dehydrogenase expression. The ratios were indicated as relative to hepatocytes.
Western blot analysis was performed to detect ALB in undifferentiated and differentiated hADMSC clusters using anti-human ALB antibody (goat polyclonal, bovine, mouse, and pig ALB-adsorbed, affinity purified; Bethyl Laboratories, Montgomery, TX) that does not react with bovine or mouse ALB.
Evaluation of urea synthesis
After washing extensively with PBS, undifferentiated hADMSCs, undifferentiated hADMSC clusters (during the same period of culture with medium stage II), differentiated hADMSCs clusters, HepG2, primary hepatocytes, and normal human fibroblasts were incubated in HBSS containing 5 mM NH4Cl for 2 h at 37°C to evaluate their capacity for urea synthesis. Each supernatant was collected and the amount of urea was determined using a QuantiChrom Urea Assay Kit (BioAssay Systems, Hayward, CA). Urea synthesis per cell was calculated based on the amount of DNA, which was determined with a DNA Quantification Kit (Primary Cell, Hokkaido, Japan) according to the instructions provided by the manufacturer.
Measurement of CYP activity
Undifferentiated hADMSCs, undifferentiated hADMSC clusters (during the same period of culture with medium stage I), differentiated hADMSC clusters, HepG2, primary hepatocytes, and normal human fibroblasts were cultured for 3 days in the absence or presence of 10 μM rifampicin (Sigma-Aldrich) based on a previous report. 25 The medium was removed and the cells were incubated for 1 h in HBSS supplemented with 100 μM 7-benzyloxy-4-trifluoromethyo-coumarin (BFC; Ultrafine, Manchester, United Kingdom). The supernatant was collected from the cultures and absorbance was measured with a spectrofluorometer at excitation and emission wavelengths of 490 and 505 nm, respectively. A standard curve was prepared from serial dilutions of 7-hydroxy-4-trifluoromethyl-coumarin (HFC; Ultrafine), and the HFC concentration in each culture medium was calculated based on the standard curve. The CYP activity per cell was calculated relative to the amount of DNA.
Uptake of low-density lipoprotein
Human low-density lipoprotein (LDL; density: 1.019–1.063 g/mL) was isolated by sequential ultracentrifugation from normolipidemic healthy donors, dialyzed against saline-EDTA, and then sterilized by filtration through a 0.2-μm filter. Lipoproteins were labeled with 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine (DiI; Molecular Probes, Eugene, OR) by incubating the LDL in 0.5% bovine serum albumin/PBS with 100 mL DiI in DMSO (3 mg/mL) for 8 h at 37°C. The lipoproteins were then dialyzed against PBS and filtered before use. To evaluate the uptake of DiI-LDL, differentiated hADMSC clusters were incubated with serum-free DMEM containing 10 μg/mL DiI-LDL for 3 h at 37°C. Finally, the cells were rinsed three times and mounted. Counter staining was performed with 4′,6-diamidino-2-phenylindole (DAPI). The slides were examined using a fluorescent microscope (Bioview, Osaka, Japan).
Periodic acid-Schiff staining for glycogen
Differentiated hADMSC clusters were fixed in 4% paraformaldehyde and embedded in paraffin. Samples were cut into 5-μm-thick sections and then oxidized in 1% periodic acid for 5 min, rinsed three times in de-ionized water (dH2O), treated with Schiff reagent for 15 min, and rinsed in dH2O for 5 to 10 min. Samples were counterstained with Mayer's hematoxylin for 1 min, rinsed in dH2O, and examined by light microscopy.
Induction of chronic liver injury in mice and transplantation of differentiated hADMSCs clusters
Six-week-old non-obese diabetic severe combined immunodeficiency (NOD-SCID) mice (Clea Japan, Osaka, Japan) were injected intraperitoneally with carbon tetrachloride (CCl4; 300 μL/kg body weight) twice a week for 12 weeks to induce chronic liver injury. Differentiated hADMSCs clusters (1.5 × 106 cells equivalent cell clusters [for differentiated cell clusters] and 5 × 106 cells for hADMSCs) were transplanted under the left kidney capsule of CCl4-treated NOD-SCID mice. Seven days after transplantation, the mice were sacrificed and blood samples were collected for determination of serum total bilirubin levels. The left kidneys were removed for histological analysis.
Histological analysis
Native liver and graft-bearing left kidneys obtained from CCl4-treated mice were fixed immediately with 10% formalin, embedded in paraffin, and processed for staining with hematoxylin and eosin, or Sirius Red. The left kidneys were also processed for Periodic acid-Schiff (PAS) staining. Differentiated hADMSC clusters and graft-bearing left kidneys were subjected to immunofluorescence staining for ALB and alpha-1-antitrypsin. They were placed into optimal cutting temperature compound (Sakura Finetechnical, Tokyo, Japan) and immediately frozen. Next, 7 μm-thick sections were fixed in 4% paraformaldehyde/PBS for 30 min. The fixed sections were then incubated with blocking solution (Blocking One; Nacalai Tesque) for 1 h. The samples were incubated with anti-human ALB antibody (Bethyl Laboratories) followed by Alexa Fluor 546-labeled donkey anti-goat IgG (Molecular Probes) and anti-human alpha-1-antitrypsin antibody (ScyTek Laboratories, Logan, Utah), followed by Alexa Fluor 546-labeled donkey anti-rabbit IgG. Counter staining was performed with DAPI. The slides were examined using a fluorescent microscope (Bioview, Tokyo, Japan).
Measurement of serum levels of ALB, alpha-1-antitrypsin, and total bilirubin
Serum levels of mouse ALB and human alpha-1-antitrypsin were determined using an ALB enzyme-linked immunosorbent assay system and human specific alpha-1-antitrypsin enzyme-linked immunosorbent assay system (both from Bethyl Laboratories) according to the instructions supplied by the manufacturer. Serum total bilirubin level was measured using QuantiChrom™ Bilirubin Assay Kit (BioAssay Systems).
Statistical analysis
All data were expressed as mean ± standard error of the mean. Differences between groups were examined for statistical significance using the Student's t-test (SPSS, Chicago, IL). A p-value less than 0.05 denoted the presence of a statistically significant difference.
Results
Characterization of hADMSCs
Flow cytometry was used to assess markers expressed by hADMSCs (Supplemental Fig. 1, available online at
Next, we examined the adipogenic, osteogenic, and chondrogenic differentiation potentials of hADMSCs. Adipogenic differentiation was confirmed by the accumulation of intracellular lipid droplets that stained positively with oil red O and the amount of oil red O was augmented by induction (Supplemental Fig. 2A, available online at
Quantitative reverse transcription–PCR analysis of immature and mature hepatocyte in clusters of differentiated hADMSCs
To promote the differentiation of hADMSCs to hepatocyte-like cells, we applied the procedure outlined in Figure 1A. In the differentiation protocol, to identify the expression of genes in other endoderm-derived cell types, or mesoderm- or ectoderm-derived cells during differentiation of hADMSCs process, we examined the expression levels of AFP, pdx-1 (pancreas), alpha-cardiac actin (mesoderm), and Sox1 (ectoderm) (Fig. 1B). Next, we used quantitative reverse transcription–PCR to determine whether our procedure resulted in differentiation of hADMSCs into hepatocyte-like cells by examining the expression levels of various markers of immature and mature hepatocytes such as AFP and CYP3A7 (as markers of immature hepatocytes) and CYP3A4, G6P, alpha-1-antitrypsin, ALB, glutamine synthetase, CYP1B1, and CK18 (as markers of mature hepatocytes) (Fig. 2). The mRNA expression levels were also assessed for differentiated hADMSCs on FN, collagen type I, or Matrigel-coated dishes. Differentiated hADMSC clusters by our procedure expressed higher levels of ALB mRNA than other differentiated hADMSCs. The expression levels of ALB, AFP, G6Pase, CYP3A7, and CYP3A4 were same as those of primary hepatocytes. AFP expression levels of primary hepatocytes and the cell clusters were same, but less than that of HepG2. The expression levels of glutamine synthetase, CYP1B1, and CK18 were higher in differentiated hADMSC clusters than other differentiated hADMSCs and HepG2. The mRNA expression level of alpha-1-antitrypsin in differentiated hADMSCs was lower than that of HepG2 and hepatocytes.
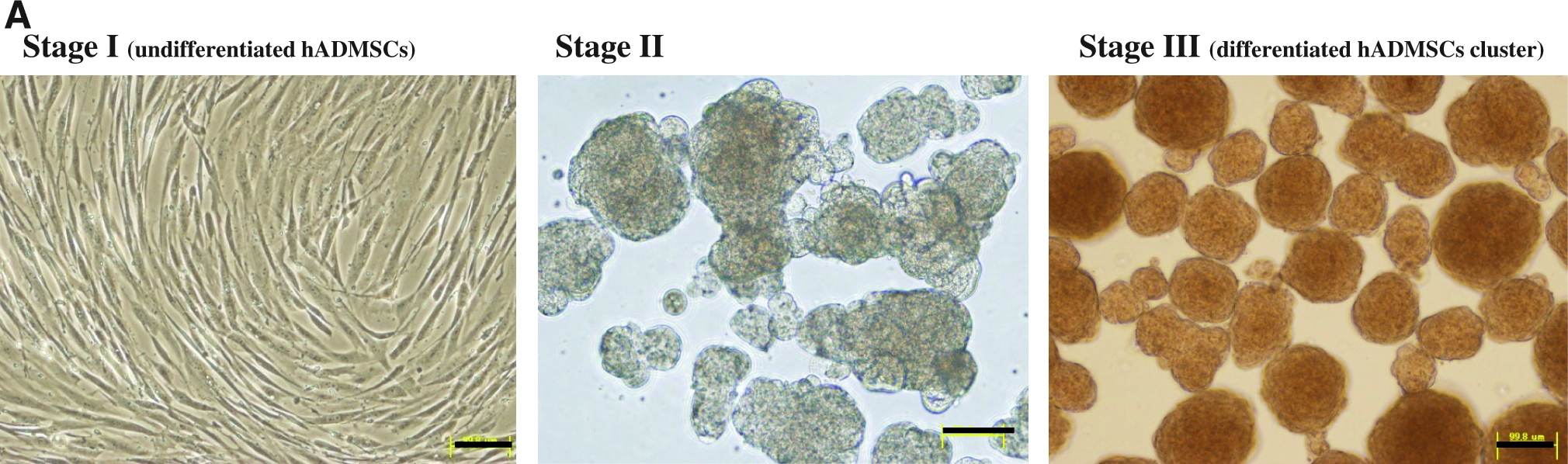
Hepatocytic differentiation of hADMSCs. (
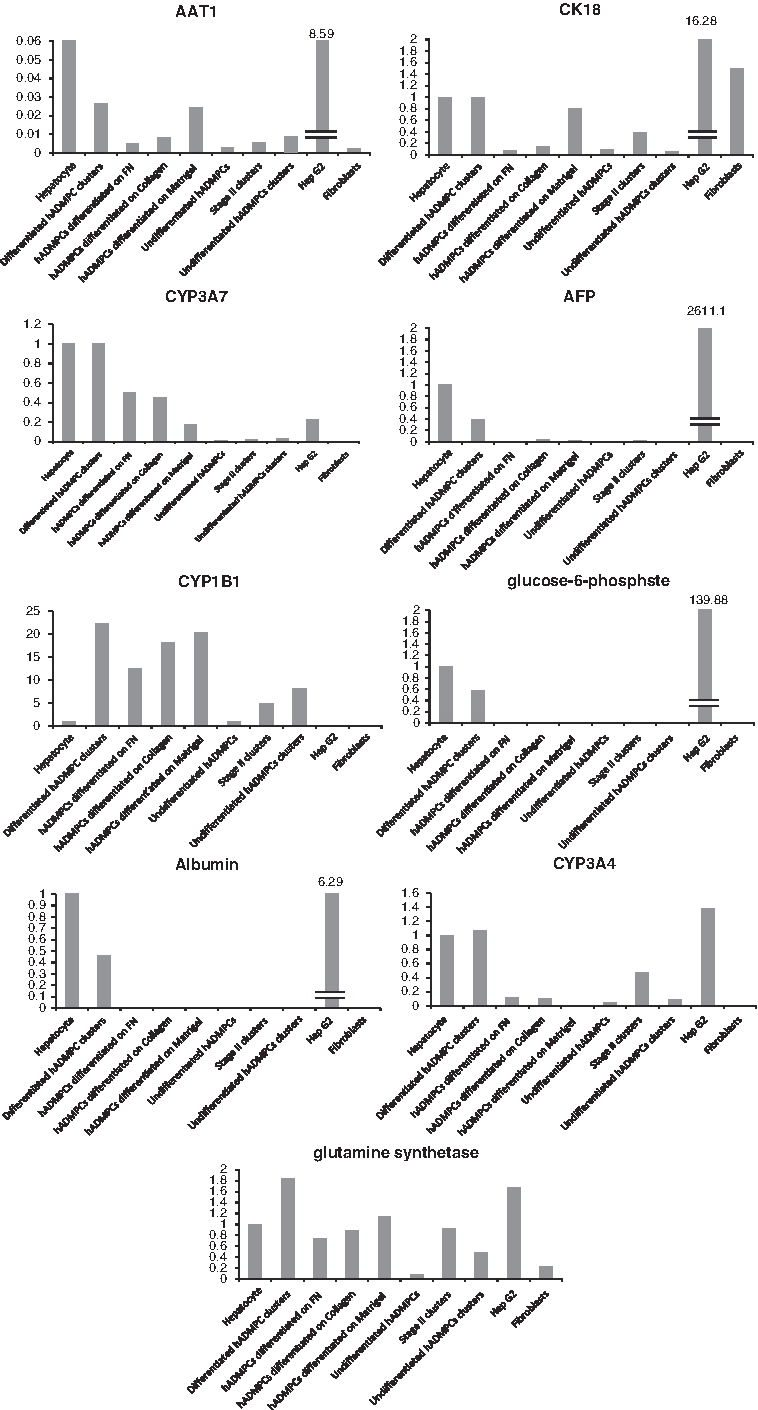
Quantitative RT-PCR analysis of the relative expression levels of mRNAs of immature and mature hepatocyte markers. RNAs were prepared from differentiated hADMSCs clusters (stage III), stage II clusters, undifferentiated hADMSCs clusters, hADMSCs differentiated on FN, hADMSCs differentiated on collagen type I, hADMSCs differentiated on Matrigel, undifferentiated hADMSCs, HepG2, and fibroblasts. Levels of AFP and CYP3A7 as markers of immature hepatocytes, and ALB, glutamin synthase, CYP1B1, CK18, CYP3A4, alpha-1-antitrypsin, and glucose-6-phosphatase mRNAs as markers of mature ones were determined by quantitative RT-PCR using Assays-on-Demand™ Gene Expression Assay Mix. The mRNA levels were normalized relative to glyceraldehyde-3-phosphate dehydrogenase expression and data are mean of triplicate experiments. FN, fibronectin; CYP, cytochrome P450; ALB, albumin; AAT, alpha-1-antitrypsin; RT-PCR, reverse transcription–polymerase chain reaction.
In vitro functional characterization of differentiated hADMSC clusters
ALB expression was assessed by western blot analysis and immunofluorescence staining (Fig. 3A, B). The former showed abundant expression of ALB on differentiated hADMSCs, and the latter confirmed these results. ALB- and alpha-1-antitrypsin-expressing cells were clearly detected among the differentiated hADMSCs. The ability to secrete urea was about 12-fold higher for differentiated hADMSCs incubated with NH4Cl, compared with stage I undifferentiated hADMSCs, and as high as that of HepG2 cells (Fig. 3C). Nonfluorescent BFC is metabolized mainly by the CYP3A family of enzymes and converted to fluorescent HFC. The concentration of HFC in the supernatant was measured after incubation with 100 μM BFC. CYP activity in differentiated hADMSC clusters was 40-fold higher than that of undifferentiated hADMSCs (Fig. 3D). In addition, CYP activity in differentiated hADMSC clusters increased 2–2.5-folds following preincubation with rifampicin for 3 days. In contrast, no increase in CYP activity was induced in undifferentiated hADMSCs under this condition.

In vitro evaluation of functional properties of hepatocyte-like cell clusters. (
We also assessed LDL uptake by differentiated hADMSC clusters by incubating differentiated hADMSCs with DiI-LDL (Fig. 3E). DiI-LDL was markedly incorporated into the cytoplasmic area of differentiated hADMSCs. Another function of hepatocytes is glycogen production (glyconeogenesis). PAS staining showed glycogen storage in differentiated hADMSCs (Fig. 3F).
Transplantation of differentiated hADMSC clusters into NOD-SCID mice with chronic liver injury
NOD-SCID mice were injected with CCl4 (300 μL/kg body weight) twice a week for 12 weeks to induce chronic liver injury (Fig. 4A). Control mice were not treated with CCl4. Histological examination confirmed destruction of acini and fibrosis of the liver in CCl4-injected mice (Fig. 4B). The serum total bilirubin level in CCl4-treated mice was significantly higher than in control mice (p < 0.05; Fig. 4C). To assess the functionality of differentiated hADMSC clusters in vivo, clusters of these cells were transplanted under the kidney capsule of mice with chronic liver injury to exclude possible cell fusion. Total bilirubin level in mice that underwent transplantation decreased significantly and approached the normal level compared with CCl4-treated mice that did not receive transplantation (p < 0.05; Fig. 4C). These data are consistent with the presence of the bilirubin transporter, multidrug resistance-associated protein 2 (MRP2), in differentiated hADMSC clusters (Supplemental Fig. 3). Histological analysis showed engrafted viable cell clusters under the kidney capsule (Fig. 4D), the presence of many human ALB-expressing cells in the graft by immunofluorescence staining, and PAS-stained engrafted differentiated hADMSC clusters.
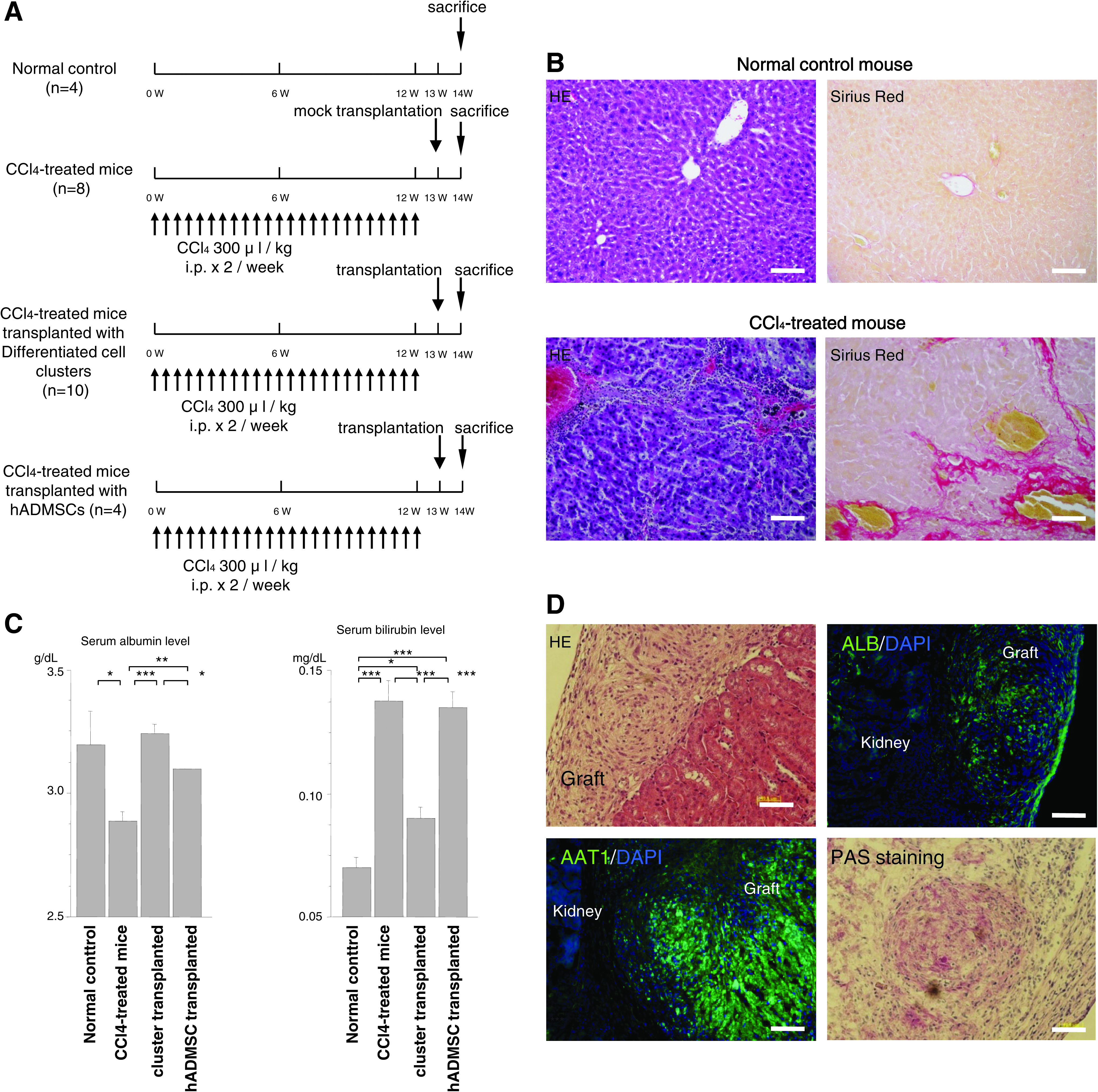
Transplantation of differentiated hepatocyte-like clusters into non-obese diabetic severe combined immuno deficiency (NOD-SCID) mice with chronic liver injury. (
Discussion
There are only few reports describing the hepatocytic transdifferentiation potential of ADMSCs.20,21 These studies described hepatocyte-like cell clusters that differentiate from ADMSCs and continue to display hepatocyte-specific functions. In this study, we generated functional hepatocyte-like cell clusters from hADMSCs that expressed high levels of mature hepatocyte markers such as ALB, glutamine synthetase, CYP1B1, and CK18. These cell clusters exhibited functional characteristics of hepatocytes; they expressed ALB, secreted urea, had CYP activity, took up LDL, and stored glycogen. Alpha-1-antitrypsin expression was markedly lower than those of hepatocytes and HepG2. Alpha-1-antitrypsin is known as an acute phase protein and stimulation by inflammatory cytokines, such as TNF-alpha and IL-6, increased its expression by not more than 100-fold in native liver. 30 It is possible that the low expression of alpha-1-antitrypsin in regenerated hepatocyte-like clusters is due to the experimental condition, that is, incubation in the absence of inflammatory cytokines.
Transplantation of regenerated hepatocyte-like clusters into NOD-SCID mice with experimentally induced chronic liver failure resulted in the secretion of hepatotrophic factors and augmentation of the function of native resident hepatocytes, as well as improvement of serum total bilirubin and ALB levels. It is possible that in transplanted animals, serum unconjugated bilirubin was converted to conjugated bilirubin by the transplanted hepatocyte-like cell clusters and soluble conjugated bilirubin was excreted in the urine because only conjugated bilirubin can be excreted into urine. It is possible that hepatosupportive factors, which were secreted by the clusters, augmented the function of liver in the transplanted animals.
For effective differentiation into hepatocyte-like cells, we introduced several improvements to the differentiation procedure. This included the formation of cell clusters and growth factor stimulation by culturing the intact cell clusters to allow for cell-to-cell interactions. In somatic stem cell differentiation, signals from the microenvironment such as cell-to-cell contacts are probably important factors that dictate the type of mature functional cells. Previous studies indicated that fetal hepatic cells and BM-derived MSCs could be induced into cells with a hepatocyte phenotype only when the cells were seeded at high density. 11 Signals induced at a high cell density, possibly via cell-cell contacts, are suspected to be necessary for hepatocytic differentiation. Further, adult hepatocyte progenitor cells were reported to show high expression levels of mature hepatocyte markers when cultured in spheroids.31–33 In our procedure, we used a floating culture and prepared cell clusters to increase cell–cell contacts. In addition, the differentiated hepatocyte-like cell clusters using our method could be transplanted as intact cell clusters. In two previous reports that described the hepatocytic differentiation potential of ADMSCs,15,16 hepatocyte-like cells were enzymatically digested to single cells and transplanted. Such transplanted cells may not function effectively. Recently, Ohashi et al. developed a cell sheet of primary hepatocytes using temperature-responsive culture dishes to maintain cell-to-cell connections and the presence of adhesion proteins. 34 They reported that transplantation of hepatocyte sheets resulted in efficient engraftment and long survival. We transplanted the intact hepatocyte-like cell clusters into animals to allow for cell-to-cell connections and maintain the presence of adhesion proteins. Detachment from extracellular matrix and single-cell dissociation using enzymes such as trypsin or collagenase were not needed. These methodological modifications allowed efficient induction of hADMSCs into functional hepatocyte-like cell clusters both in vitro and in vivo.
In conclusion, we developed a simple protocol for efficient differentiation of hADMSCs into hepatocyte-like cell clusters. These hepatocyte-like cell clusters exhibited functional characteristics of hepatocytes. These in vitro-introduced hepatocyte-like cells may provide a potentially unlimited source of hepatocyte-like cells without the limitation of immune rejection.
Footnotes
Acknowledgments
This study was supported in part by a grant-in-aid for Akifumi Matsuyama from the Kobe Translational Research Cluster, the Knowledge Cluster Initiative, Ministry of Education, Culture, Sports, Science, and Technology (MEXT) and from the Program for Fundamental Studies in Health Sciences of the National Institute of Biomedical Innovation (NIBIO), and in part by a grant-in-aid for Yoshiki Sawa from the New Energy and Industrial Technology Development Organization (NEDO) of Japan.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.