
Abstract
Primary neurons were grown on structured silicon (Si) substrates, in the absence of chemotropic factors or synthetic extracellular matrix. The Si substrates used for the study comprise hierarchical structures in the micro- and nanolength scales. The substrates were structured via femtosecond laser irradiation of the Si wafer, in a reactive SF6 environment. Electron microscopy revealed that the neurons formed an elaborate web of cytoplasmic processes in the absence of glial elements. The neuronal cytoplasm autografted the depth of the spikes, and the neurite sprouting took place over the spikes surface. Here we demonstrate how microfabrication of a Si surface provides an excellent platform for multifaceted studies of neuronal specimens.
Introduction
Neurons depend on surface support more than other cells, but readily adhere to surfaces coated with ECM proteins. Moreover, brain development and neuronal networks formation depend on an intricate blueprint of axon guidance, dendritic arborization, and synapse formation cues. The challenge to comprehend the multitude of cues neurons receive, process, and direct, in vivo, and to simulate 3D cultures in vitro, has been addressed; however, the results are limited.10,11 Recently, Pautot et al. addressed this subject developing the first 3D neuronal network on silica beads. 11 The importance of 3D neuronal cultures can be realized if one thinks that the dimensionality of the network has a strong impact on its connectivity, and therefore plays an important role for its possible behavior. 12 In our study, we have utilized silicon (Si) microstructured substrates to enable neurons to grow in 3D, without coating, synthetic ECM, or chemotropic factors.
Materials and Methods
Substrate preparation
The preparation of the substrates involved the microstructuring of the flat Si surface using ultrafast laser, under a reactive gas atmosphere (SF6). A single crystal n-type Si (100) wafer was mounted on a high-precision X–Y translation stage normal to the incident laser beam, in a vacuum chamber evacuated at a base pressure of 10−2 Torr. The pressure of the reactive gas was kept constant during irradiation (500 Torr). The irradiating source was a regenerative amplified Ti:Sapphire laser (λ = 800 nm, τ = 180 fs pulse duration) delivering pulses at a repetition rate of 1 kHz. The laser pulsed fluence used was 0.21 J/cm2. A mechanical shutter was synchronized with the stage motion to provide a uniform exposure of a 2 × 2 mm2 area to an average of 500 pulses. Following the irradiation process, the samples were first cleaned in ultrasonic baths of trichloroethylene, acetone, and methanol followed by a 10% hydrofluoric acid aqueous treatment to remove the oxide grown on the surface.
The thermal oxidation of Si is performed in air at 1000°C for 1 h, resulting in the formation of 110 nm silicon oxide layer.
The morphology of the samples' surface, prior as well as after the cells culture, was characterized by field emission scanning electron microscopy (SEM) (Jeol 7000 Tokyo, Japan).
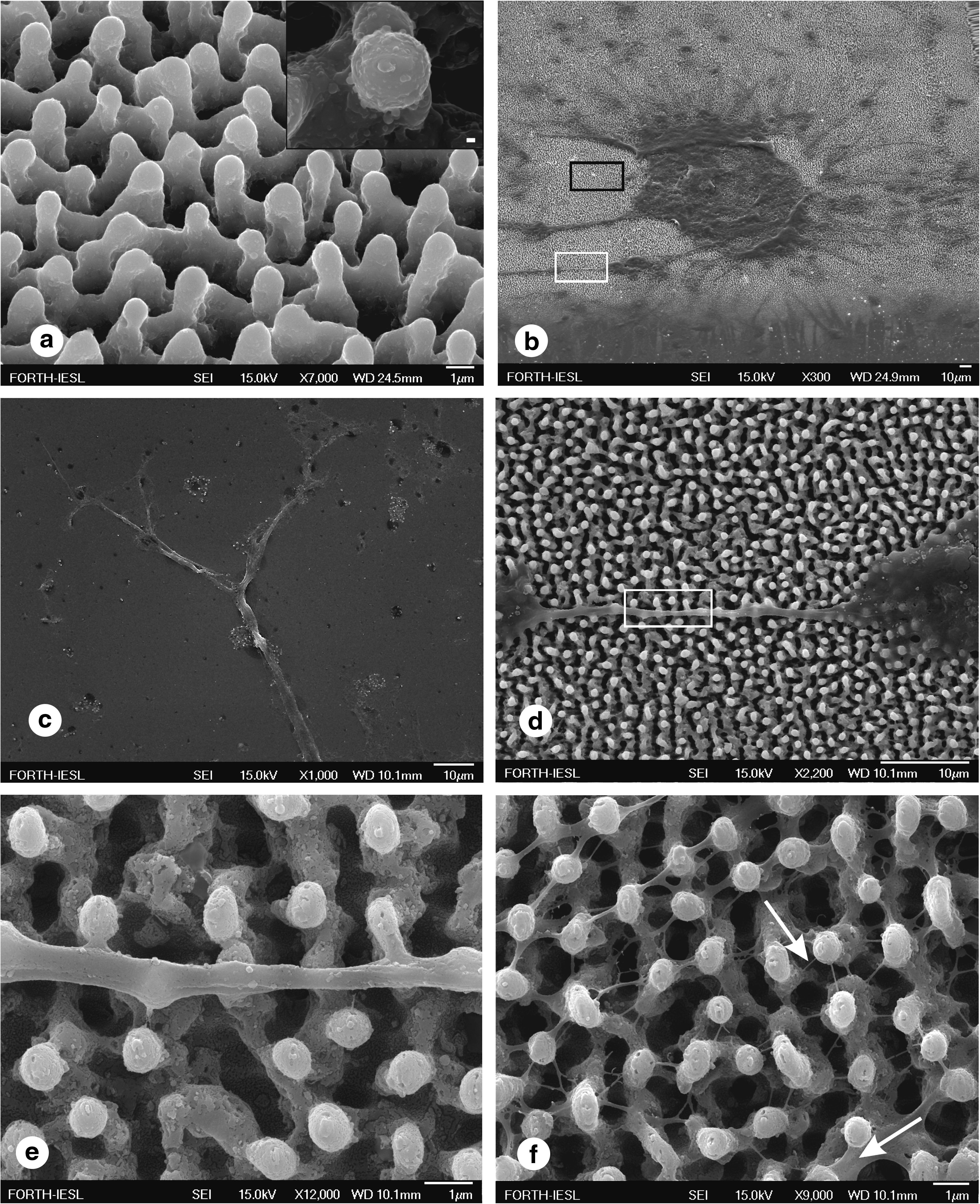
As clearly seen in Figure 1a, the surfaces possess hierarchical micro- and nanostructures. 13
(

Preparation of primary neuronal cultures
Fetal brains were obtained from timed pregnant mice at embryonic day 14. After removing dura-arachnoid membranes, whole brains were isolated and incubated in Hank's balanced salt solution medium (Biochrom, Berlin, Germany). After series of washings at low centrifugation speed, with the supernatant discarded, the pellets were resuspended in a small volume of Dulbecco's modified Eagle's medium (DMEM/F12:1/1) (Biochrom) medium. After vigorous trituration the cells were resuspended to a final volume, in DMEM/F12 supplemented with penicillin (100 U/mL), streptomycin (100 μg/mL), and B-27 (Gibco) (DMEM/F12: B-27 = 50:1). Finally, the neurons were plated on polyornithine-precoated dishes. The primary neuronal cultures were kept at 37°C, in a 5% CO2 humidified incubator for 3 days and then were enzymatically (Trypsin–Gibco, 0.05% Trypsin–ethylenediaminetetraacetic acid) detached and plated on the substrates under study. The density with which cells were plated on the substrates was 2.5 × 106 cells/well.
Immunofluorescence
Cells grown on the substrates under study were fixed at 5 days in vitro in 4% paraformaldehyde in 0.1 M phosphate buffer for 5 min and permeabilized in phosphate-buffered saline containing 0.1% Triton X-100 (PBST). The cells were blocked in PBST with 10% serum for 10 min, and all subsequent reactions were carried out in PBST with 2% serum. A monoclonal anti-beta III tubulin primary antibody (Promega G712A; 1:1000, Madison, WI), a rabbit polyclonal anti-glial fibrillary acidic protein (GFAP) (Abcam-AB7260; 1:3000, Cambridge, United Kingdom), Cy5-conjugated anti-mouse secondary antibody (1:500), and a 7-amino-4-methylcoumarin-3-acetic acid (AMCA)-conjugated anti-rabbit secondary antibody (1:500) obtained from Jackson Immunoresearch Laboratory, West Grove, PA, were used for the immunostaining. Beta III tubulin was used as a neuronal marker because its initial expression begins during or immediately after the completion of mitosis in neuroblasts. 14
Confocal microscopy
The digital images were acquired on Leica TCS SP5 confocal microscope using the LAS AF software. The 488 and 633 nm laser lines were used to view spikes (at reflection mode) and cell immunofluorescence, respectively.
Results and Discussion
Microstructured roughness in the form of spikes promoted the adhesion of single or clusters of neurons to the substrate (Fig. 1b). In contrast, only few cells survived on a flat Si substrate used as a control (Fig. 1c) to verify the role of roughness in absence of synthetic ECM. The cells developed into a dynamic cellular aggregate, with long neurites sprouting over the spikes' surface (Fig. 1d). The integrity and uniform sprouting observed suggest the biocompatibility of the neuronal population with the surface. The protrusions of the neurolemma grew over the spikes, engulfed their top (Fig. 1e), and incorporated them to form a web. Moreover, a network of nanoscale thin neuritic extensions was formed along the third dimension (-z axis) utilizing the space toward the base of the spikes (Fig. 1f). A SEM micrograph of the sample's cross section demonstrates the mechanically sheared cell membrane and the engulfed Si spikes (Fig. 1g). Having enveloped the top of the spike, the thicker extensions appeared (∼550 nm) and the processes continued toward the bottom, with thinner nanoextensions (∼35 nm). A Si substrate with nonperiodical spikes exemplifies the ergonomic way of neuritic sprouting, because neuritic extension is dictated by the shortest intraspike distance (Fig. 1i). In addition, multiple protrusions extended along the z-axis from one single neurite.
A snapshot of the formation of nonsubstrate-constrained growth cone filopodium-like structures is shown in Figure 1h. A blindly winding end (indicated by the arrow) has reached the closest adhesion point and the process was limited by a final adhesion step (asterisk). Such filopodia may grow to physically bridge the distance between the membranes aiming to construct a network. This observation is supported by the Hammarback and Letoumeau guide post-hypothesis that growth cones quickly cross a low-adhesivity substrate gap in vitro, when this gap is spanned by even a single filopodium establishing contact on a distant high-adhesivity substrate. 15
With the cell membrane being amphiphilic, surface chemistry is also an important factor for cell attachment. The importance of surface chemistry on neuronal survival was tested utilizing a geometrically identical Si substrate, following thermal oxidation. Neurons sensed the vertical microstructured roughness and adhered. However, a prominent feature on the spike area was the extended cellular death. Although the oxidized Si spikes maintained the same geometrical microenvironment as the bare Si spikes, they did not allow long-term cell survival. A possible explanation is that Si spikes covered with silicon oxide are super hydrophilic (contact angle water less than 5°), whereas bare Si spikes are hydrophobic, with contact angles close to 130°. Consequently, biomaterial roughness is not a stand-alone prerequisite for neuronal survival and growth, but it rather requires the synergistic effect of surface chemistry. 16 The importance of these findings is multifaceted.
The culture conditions used favored the sole growth of primary neurons, and confocal microscopy was employed to confirm the immunofluorescence of tubulin beta III, which is specific for mature neurons. The cells used here were beta III tubulin immunopositive, and the comparable immunofluorescence (Fig. 2A) and SEM (Fig. 1b) pictures confirm that there was no glial feeder layer on the substrates under study. Hence, the immunostaining enabled us to confirm the neuronal specificity of our cultures, to verify the shape of adhesion on the substrates, and to measure a significant increase in length for some of the neurons 17 (Fig. 2B–D). In addition, primary neuronal cultures were stained with specific antibodies both for beta III tubulin (red) and GFAP (blue) at 5 days in vitro, as shown in Figure 3. The micrograph demonstrates the low numbers of glial cells attached to the Si microstructured substrates. It is worth emphasizing that when the cells were plated on the oxidized Si substrates, adhesion was limited. The wet culture conditions did not compromise the stability of the substrates. In addition, neuronal adhesion was not superficial. This was established by the attachment, growth, and elongation of neurites on the substrates, in all three dimensions. The undeniable proof of outstanding adhesion properties was derived from the electron micrographs demonstrating how the neuronal membranes incorporated the tips of the spikes as their own. Besides promoting neuronal adhesion, the microscale roughness in synergy with the surface chemistry allowed the formation of an exquisitely organized 3D neuritic network. The use of hydrophobic, Si microstructures enabled the “autografting” of these structures by the neurons, meaning that the neurons cloaked the spikes, thus self-grafting the area. The ability of the neurons to adjust their shape via dynamic cytoskeletal remodeling is intrinsic. Nevertheless, these intracellular processes that drive membrane extensions, such as migration or filopodial lengthening, work against the membrane's mechanical properties. In this study, we demonstrated that inherent neuronal guidance cues were sufficient to win over steady-state membrane mechanics. The outcome was to allow the filopodia of growth cones to explore this permissive 3D microenvironment, and to “decide” its direction depending on the shortest distance.

(

Primary neuronal culture immunostained both for beta III tubulin (red) and GFAP (blue) at 5 days in vitro. The micrograph demonstrates the low numbers of GFAP-immunopositive cells and staining colocalization.
Our work demonstrates a novel approach that utilizes Si-microfabricated substrate to support growth, and 3D development of cultured neurons, in the absence of chemotropic factors or synthetic ECM. This approach provides a functional platform allowing the study of nervous system properties, including the ability of repair with potential usefulness in biomedical investigations.
Footnotes
Acknowledgments
Part of the work was performed at the Ultraviolet Laser Facility operating at IESL-FORTH and has been supported in part by the European Commission through the Research Infrastructures activity of FP6 (“Laserlab-Europe” RII3-CT-2003-506350) and in part by Marie Curie Transfer of Knowledge project “NOLIMBA” (MTKD-CT-2005-029194). A.S. acknowledges NARSAD for financial support. We thank Sandra Siakouli for expert help with the preparation of the samples for field emission SEM.
Disclosure Statement
No competing financial interests exist.