
Abstract
Stem cells offer an interesting tool for tissue engineering, but the clinical applications are limited by donor-site morbidity and low cell number upon harvest. Recent studies have identified an abundant source of stem cells in subcutaneous adipose tissue. Adipose stem cells (ASCs) present in adipose tissue are able to differentiate to several lineages and express multiple growth factors, which makes them suitable for clinical application. Buccal fat pad (BFP), an adipose-encapsulated mass found in the oral cavity, could represent an easy access source for dentists and oral surgeons. The stromal vascular fraction obtained from fresh BFP-derived adipose tissue and passaged ASCs were analyzed to detect and quantify the percentage of ASCs in this tissue. Here we show that BFP contains a population of stem cells that share a similar phenotype with ASCs from abdominal subcutaneous fat tissue, and are also able to differentiate into the chondrogenic, adipogenic, and osteogenic lineage. These results define BFP as a new, rich, and accessible source of ASCs for tissue engineering purposes.
Introduction
In the late 1960s Friedenstein et al. demonstrated that the mesenchymal stroma from human bone marrow contains a population of cells that proliferate when cultured on plastic and differentiate to cell lineages derived from the mesoderm, such as chondrocytes and osteoblasts. 1 Later, these precursor spindle-shaped cells are referred to as mesenchymal stem cells (MSCs), 2 and shown to differentiate to several lineages in vitro 3 and in vivo,4–6 making these cells promising candidates for mesodermal defect repair. However, the clinical use of MSCs provides several problems, such as pain associated with the harvest procedure, complexity of the technique, and low cell number upon purification, especially in old donors. 7 This makes an ex vivo expansion step necessary to obtain therapeutic cell doses, which is time consuming and expensive, and contains the risk of cell contamination and cell loss.
Adipose tissue represents a promising source of MSCs available in large quantities, which does not require the use of general anesthesia and results in minimal patient discomfort. Further, adipose tissue yields higher numbers of MSCs than bone marrow, which could avoid cell expansion. 8 The nomenclature of MSCs from adipose tissue varies widely, but the consensus reached at the Second Annual International Fat Applied Technology Society Meeting (2004, Pittsburgh, PA) was to use the term “adipose stem cells” (ASCs). ASCs are able to differentiate to multiple mesenchymal tissue cell types, such as osteoblasts, chondrocytes, adipocytes, myocytes, cardiomyocytes, and endothelial cells, and there is growing evidence suggesting that they can also give rise to cells from other lineages, such as ectoderm and endoderm.9–12 In addition, adipose tissue secretes a variety of angiogenic and antiapoptotic growth factors that makes fat a promising source for reconstructive surgery. 13 ASCs have been used successfully in several experimental studies in the bone tissue engineering field.14–17 Therefore, adipose tissue is an interesting source for cell-based therapy.
The oral cavity contains a mass of specialized fatty tissue, buccal fat pad (BFP) or Bichat's fat pad, that is distinct from subcutaneous fat. 18 The easy accessibility and rich vascularization make this fat mass an attractive graft, which has been used widely in oral surgery for the repair of bone and periodontal defects.19–22 The harvesting of BFP is a noncomplicated procedure, which requires a minimal incision with local anesthesia, and causes minimal donor-site morbidity.
This study demonstrates that BFP is a rich and accessible source of stem cells that express the characteristic markers of ASCs and that can differentiate to chondrocytes, osteoblasts, and adipocytes.
Materials and Methods
Tissue preparation and cell isolation/expansion
Subcutaneous abdominal adipose tissue (SC) and BFP were obtained from healthy individuals undergoing elective abdominal and orthognathic surgery procedures in different Hospitals (Hospital Clínic and Hospital Sant Joan de Déu, Barcelona, Spain). The age range of the patients submitted to abdominal surgery was 19–57 years (n = 4), and the age range of the patients undergoing oral surgery was 17–40 years (n = 10). All patients were in good health, and no diabetes or other systemic complications were reported. The informed consent and experimental protocols in this study were reviewed and approved by the Ethics Committee of the Universitat Internacional de Catalunya in Sant Cugat (Barcelona, Spain). Sample tissues were processed according to a modification of a previously reported procedure. 23 Raw oral fat tissue (16 ± 4.1 mL) and subcutaneous fat tissue (22 ± 12.2 mL) (mean ± standard deviation [SD]) were washed several times with sterile phosphate-buffered saline (PBS), minced into small pieces, and treated with 0.075% collagenase I (Sigma, St. Louis, MO) for 60 min at 37°C. After incubation, adipose tissue was centrifuged at 400 g for 10 min to separate the adipocytes and lipid droplets from the stromal vascular fraction (SVF).
Cell pellets were re-suspended in red blood cell lysis buffer (8.2 g/L NH4Cl, 0.84 g/L NaHCO3, and 0.37 g/L disodium ethylenediaminetetraacetic acid, pH 7.4) and incubated for 10 min at room temperature. SVF cells were resuspended in low-glucose Dulbecco's modified Eagle's medium containing 10% fetal bovine serum (FBS) and 100 units/mL antibiotics/antimycotics solution. Suspended cells were passed through a 100 μm cell strainer (BD Biosciences, Palo Alto, CA), cells were counted, and their viability was assessed with Trypan Blue exclusion. Cells were seeded at 5 × 103 cells/cm2 in 100 mm tissue culture dishes and maintained in a humidified incubator at 37°C and 5% CO2. After 3–4 days individual cell colonies were visible upon microscopic examination. The initial adherent cell population, referred to as passage 0 (P0), as well as after several passages (up to 1 month of culture), was analyzed by flow cytometry. After 1 week of culture adherence, cells from BFP (n = 5) and abdominal fat (n = 4) were counted, and cell number per gram of adipose tissue was calculated. Cells from BFP in P2 (n = 3) were induced to differentiate in the adipogenic medium (AM), osteogenic medium (OM), or chondrogenic medium (CM) for several weeks. Differentiation media consisted of high-glucose Dulbecco's modified Eagle's medium and lineage-specific medium supplementation as described
23
(Supplemental Table S1, available online at
Human osteogenic sarcoma cells (SAOS-2) were obtained from the American Type Culture Collection (ATCC HTB85, Manassas, VA) and cultured in McCoy's 5a medium (Gibco BRL, Grand Island, NY) containing 15% FBS and used as osteogenic control. Fresh adipose tissue was obtained from BFP as well as from excised abdominal adipose tissue from patients and used as adipogenic control. Chondrocytes derived from human nucleus pulposus were used as chondrogenic control.
Flow cytometry
Flow cytometry was performed with fresh adipose tissue from BFP (n = 5), and passaged ASCs were obtained from BFP (n = 4) and SC (n = 4) from patients undergoing elective abdominal surgery. For flow cytometry analysis, cells were resuspended at 106 cells/mL in the control medium. Fifty microliters of the cell suspension containing 3–5 × 105 cells was incubated with 5 μL primary monoclonal antibodies (MAbs) directed against the following antigens and coupled to phycoerythrin (PE), fluorescein isothiocyanate (FITC), or peridinin–chlorophyll–protein complex (PerCP) for 30 min at 4°C: CD14[PE], CD34[PerCP], CD45[PerCP], CD29[PerCP], CD73[PE], SSEA[PE], CD90[FITC], CD146[FITC], HLA-DR[FITC], and CD105[FITC]. All MAbs were purchased from BD Biosciences, except CD105 and CD146, which were purchased from R&D Systems (Minneapolis, MN) and eBioscience (San Diego, CA), respectively.
All samples were washed in PBS containing 2% FBS, fixed with Cell-Fix (BD Biosciences), and incubated on ice for 30 min. Samples were then analyzed by FACScan flow cytometry (Becton Dickinson Immunocytometry Systems, San Jose, CA). Aliquots of cells that were incubated with isotype-matched mouse anti-human IgGs served as negative control. Data were analyzed using CellQuest software and FACSDIVA for SVF (Becton Dickinson Immunocytometry Systems).
Immunohistochemistry
Cells were rinsed twice with PBS and fixed with 3% paraformaldehyde for 20 min at room temperature. Thereafter, cells were incubated in 100 mM NH4Cl in PBS for 10 min to quench autofluorescence. Nonspecific binding was blocked by incubating the cells in 1% bovine serum albumin (BSA) in PBS containing 0.2% Triton X-100 for 15 min at room temperature. Osteogenically stimulated cells and undifferentiated ASCs were incubated for 1 h with a mouse anti-human osteocalcin MAb (R&D Systems) diluted 1:100 in PBS with 1% BSA, and anti-mouse Alexa 546 secondary antibody was used at a concentration of 1:500. Chondrogenically stimulated and undifferentiated ASCs were pretreated with pepsin solution (Zymed Laboratories, San Francisco, CA) for 10 min at 37°C and incubated for 1 h with a mouse anti-human type II collagen antibody (Acris antibodies, Hiddenhausen, Germany) diluted 1:5 in PBS with 1% BSA. Cells were conjugated with anti-mouse fluorescein secondary antibody diluted 1:500 (Chemicon International, Temecula, CA).
Histology
Differentiated and undifferentiated ASCs were processed using the following histological assays: for adipogenesis, Oil Red O stain (Sigma) was used to detect intracellular lipid accumulation according to standard protocols. Cells were counterstained with Mayer hematoxylin and eosin solution (Sigma). For osteogenesis, Alizarin Red solution (Millipore, Billerica, MA) was used to detect mineral deposition according to the manufacturer's instructions. For chondrogenesis, Toluidine Blue (TB) (Panreac, Barcelona, Spain) was used to assess proteoglycan extracellular matrix according to standard protocols. Stained slides were examined by microscopy.
Alkaline phosphatase activity assay
Triplicate ASC cultures were induced to differentiate in OM for up to 3 weeks. At 7 and 21 days, cells were washed twice with PBS, and lysed in 10 mM Tris-HCl (pH 8.2) containing 2 mM MgCl2 and 0.05% Triton X-100 at 4°C for 1 h. The lysates were centrifuged at 200 g for 10 min at 4°C, and submitted to two freeze–thaw cycles. The lysates were cleared by centrifugation at 11,300 g for 30 min (4°C) and incubated for 1 min in prewarmed alkaline phosphatase (ALP) solution provided by Linear Chemicals (Barcelona, Spain). p-Nitrophenolate release was determined spectrophotometrically at 405 nm. Protein concentration in the supernatant was analyzed using the BioRad Protein Assay (Bio-Rad Laboratories, München, Germany) with BSA as standard. Undifferentiated ASCs served as negative control and SAOS-2 cells as positive control.
Quantitative real-time polymerase chain reaction
Gene expression of peroxisome proliferating receptor gamma (PPARγ) was quantified in adipogenically stimulated cell cultures. Total cellular RNA was extracted from SC, BFP, and ASCs cultured in AM for 3, 9, and 21 days, using Trizol reagent (Invitrogen, Carlsbad, CA), and 2 μg of RNA was reverse-transcribed using M-MLV Reverse transcriptase (Invitrogen). Undifferentiated ASCs and human adipose tissue from SC and BFP served as control. Expression of PPARγ and the osteogenic genes core-binding factor alpha subunit 1 (CBFA1) and osteonectin (SPARC) was quantified in ASCs cultured in OM for 7, 14 (CBFA1, SPARC, and PPARγ), and 21 days (PPARγ). Undifferentiated ASCs and SAOS-2 cells were used as control. Expression of PPARγ and SOX9 was analyzed in ASCs cultured for 5 days (PPARγ) and 14 days (SOX9 and PPARγ) and in CM. Undifferentiated ASCs and chondrocytes derived from human nucleus pulposus were used as chondrogenic control. Real-time polymerase chain reactions (PCRs) were performed using 2.5 μL cDNA and SYBR Green Supermix (Bio-Rad Laboratories). Triplicate reactions were performed using primer pairs for PPARγ,
24
CBFA1, osteonectin, and SOX9. The oligonucleotide sequence of the primers is listed in Supplemental Table S2 (available online at
Statistical analysis
Average values were expressed as mean ± SD. ALP and real-time PCR was expressed as mean ± standard error of the mean. Marker expression differences between two different sources of ASCs were analyzed statistically using a nonparametric Mann–Whitney U-test. Differences were considered significant if p < 0.05.
Results
BFP contains progenitor cells with ASC characteristics
To study if BFP contains progenitor cells with ASC characteristics, BFPs from healthy patients were extirpated and SVF was isolated. Freshly isolated and cultured SVF was analyzed for cell morphology and membrane marker profile to observe whether the cells derived from BFP share characteristics with ASCs from another fat depot. After 1 week of culture, adherent cells from abdominal fat (n = 4) and BFP (n = 5) were counted, and cell number per gram of adipose tissue was calculated.
Morphology
Some adherent cells were observed 48 h after seeding. Nonadherent cells disappeared over time in culture (Fig. 1A). Cells from BFP and SC remained in a quiescent phase (dormant) during 2–4 days; afterward, they began to multiply rapidly, approaching confluence as a monolayer of large flat cells. Both BFP and SC cells showed similar morphology; that is, they were spindle shaped. After 7 days of culture, both BFP and SC cells exhibited a more fibroblast-like morphology characteristic of ASCs. At this time point, cells reached 90% confluence and were passaged.
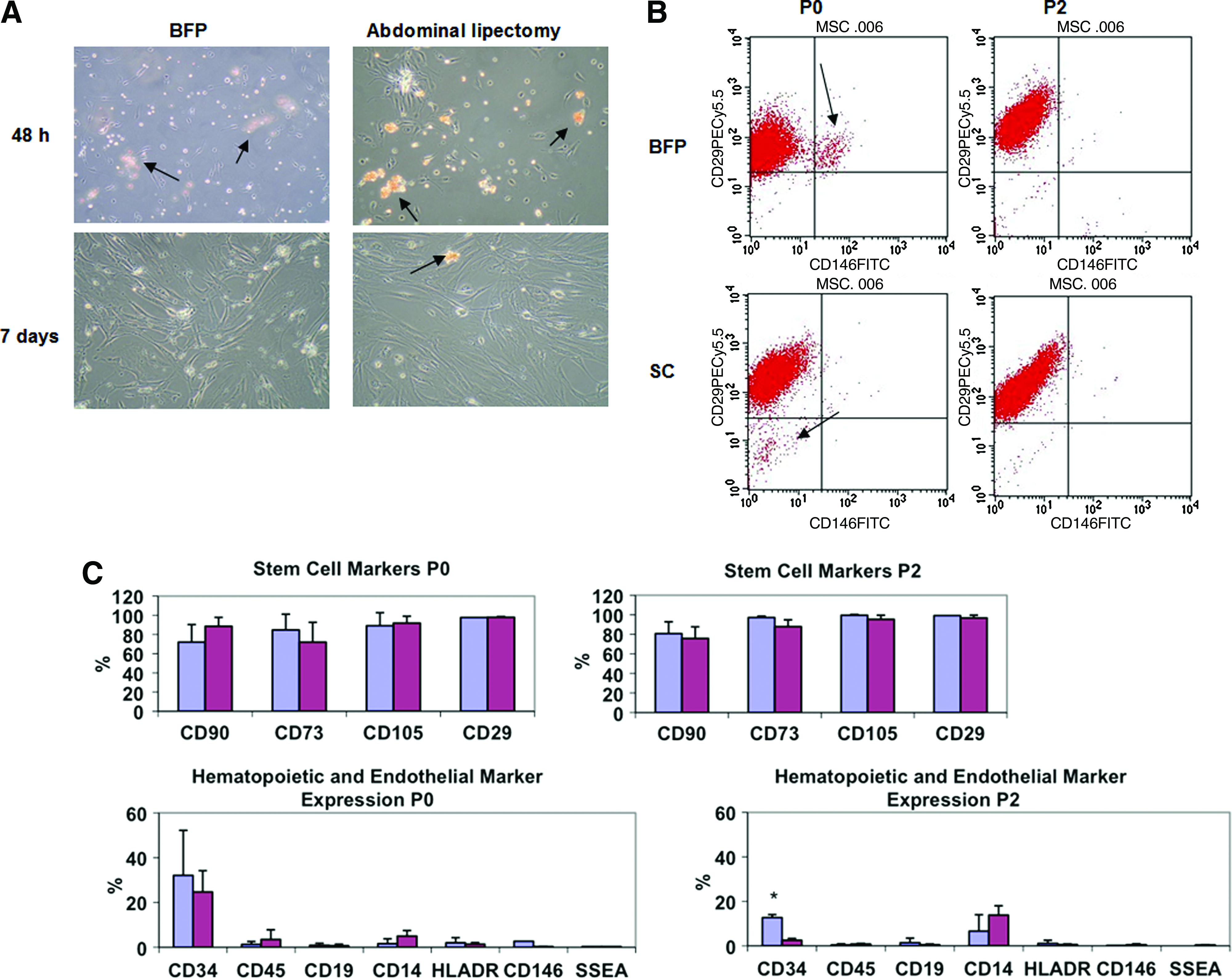
Comparison of morphology and phenotype of ASCs derived from BFP or SC. (
Cell surface marker profile
To evaluate the surface marker profile, cells were stained with a panel of antibodies for marker expression to define the percentage of ASCs in BFP and SC, following the minimal criteria of marker expression to define MSCs. 25 Cultured ASCs from BFP and SC were positive for CD73, CD90, and CD105, and negative for the hematopoietic markers CD45, CD19, CD14, and HLADR (Table 1). Cells were also positive for CD29, a characteristic marker of ASCs, 26 and negative for CD146, a marker expressed in endothelial and vascular smooth muscle cells. 27 Some cross-contaminating populations were found in initial passaged ASCs from BFP and SC, but they disappeared with further passaging (Fig. 1B). A CD146+CD29+ population was observed in initial passaged ASCs from BFP, but not SC. CD146 is characteristic of endothelial progenitor cells, which are likely found in the rich microvasculature within this adipose tissue.
Results are shown as mean ± standard deviation of the percentage of cells expressing a marker. Cells were analyzed at 7 days of culture (P0), passage 2 (P2), and passage 5 (P5) of culture.
For CD29, CD146, and SSEA, marker expression at P5 cells of one patient were analyzed.
After 1 month of culture of ASCs from BFP (P5), more than 98% of the cells expressed the characteristic MSC markers CD105, CD73, and CD29, and less than 2% cells expressed the hematopoietic markers CD45, CD19, CD14, and HLADR, according to previous studies. 28 ASCs from BFP showed some expression of CD34 (32%), but this declined with time in culture to 7.5% at 1 month of culture. Table 1 shows the marker expression percentage of ASCs from BFP, as mean ± SD measured at several passages. Figure 1C shows the comparison between marker expression of ASCs from BFP and SC at P0 and P2. Marker expression was similar for BFP and SC, except CD34 expression, which was much higher in ASCs from BFP at P2 (p < 0.05) than in ASCs from SC.
Cell yield
Cell yield from adipose tissue obtained from patients undergoing elective oral or abdominal surgery was analyzed (Table 2). The mean age was 19.6 years for patients undergoing oral surgery (n = 5, range 18–22 years), and 43.3 years for patients undergoing abdominal surgery (n = 4, range 19–57 years). The mean yield of ASCs obtained from BFP after 1 week of culture was 513 ± 227 × 103 cells per gram of tissue, and the mean yield of ASCs from abdominal adipose tissue was 253 ± 56 × 103 cells per gram of adipose tissue. This result shows more ASCs in BFP than in SC; however, it did not reach significance due to high patient variability. The number of ASCs obtained from SC was similar as observed in another study. 29
Cell number is expressed per gram of adipose tissue after 1 week of culture.
SVF from BFP is a rich source of ASCs
Adipose tissue can be split in two different fractions, that is, one fraction containing mature adipocytes and another fraction, SVF, containing a heterogeneous cell population. To our knowledge, the SVF from BFP has not yet characterized. To determine the percentage of ASCs present in this fraction, we characterized the SVF from five patients. Coexpression of different markers was determined by flow cytometry to quantify the cell population representing fresh ASCs. Patient age was between 19 and 29 years, and 60% were women. SVF from BFP contained a mixture of cells, as shown by representative dot plots from the different patients and percentages of marker coexpression (Fig. 2B, C). The SVF contains a high percentage of cells (21%–38%) that express CD90, CD73, CD29, and CD34, and which do not express the hematopoietic and endothelial lineage markers (CD45, CD19, CD14, HLADR, and CD146). In addition, this population of cells does express CD34, but not CD105, as reported previously.30,31 This marker profile is characteristic of fresh ASCs, whereas culture of ASCs on plastic plates increases CD105 expression and reduces CD34 expression.30,31 Taken together, these results demonstrate that SVF from BFP is a rich source of ASCs (∼30%), similar to other sources of adipose tissue.
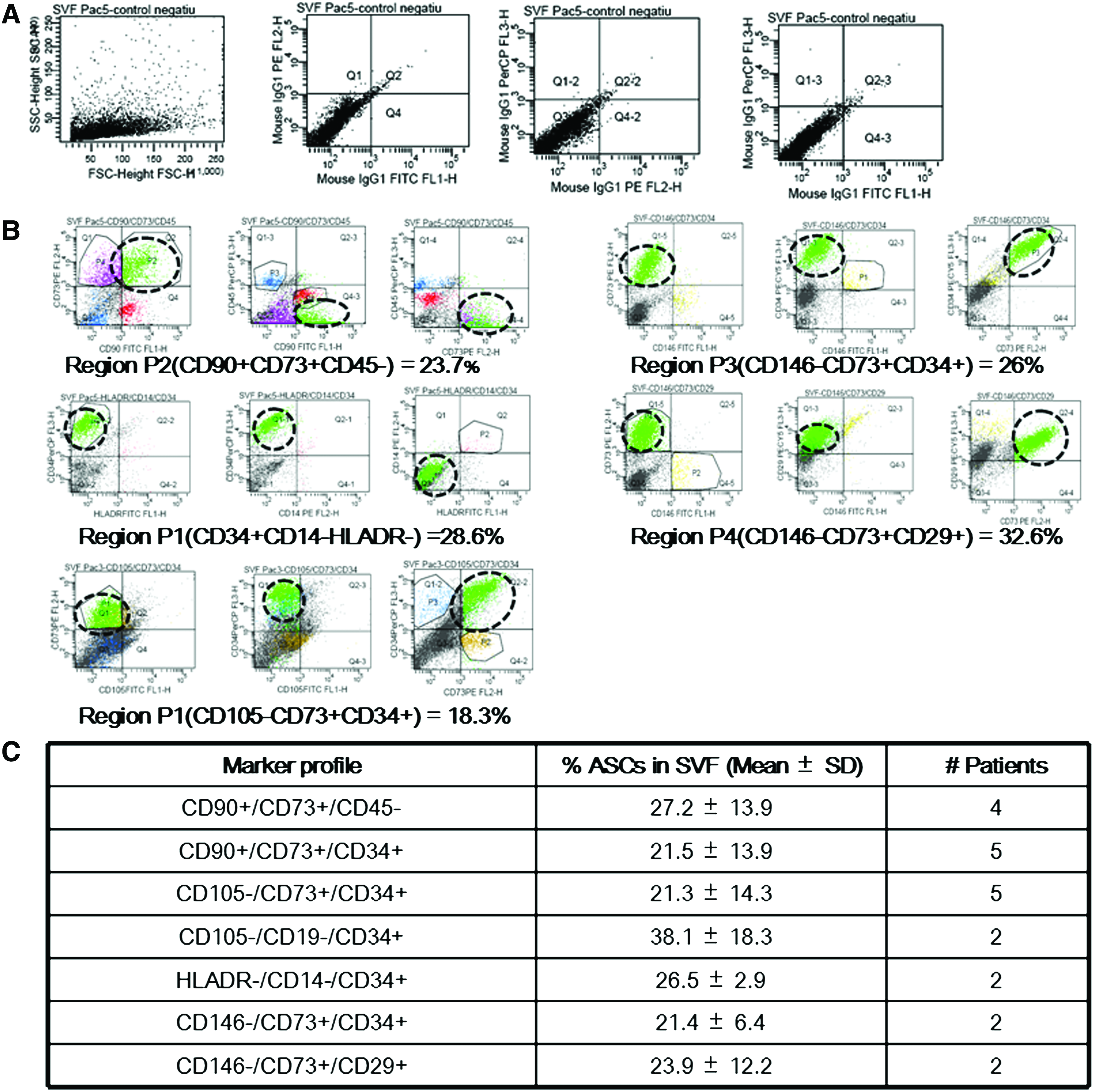
Flow cytometry of fresh ASCs obtained from BFP, analyzed by FACS DIVA software. (
Multilineage differentiation potential of ASCs derived from BFP in vitro
Osteogenesis
To assess the osteogenic potential of ASCs, ALP, an early marker of osteoblastic differentiation, was determined after 0, 7, and 21 days of culture in OM. SAOS-2 cells were used as a positive control. After 1 week of culture, ASCs changed their morphology from spindle shaped to more polygonal shaped, which was accompanied by an increase in ALP activity up to day 21 (Fig. 3A). ASCs cultured in OM showed a 2.5-fold increase in ALP activity at day 7, and a 16.5-fold increase at day 21. Areas of high granular density appeared and multiple layers of ASCs were formed after 2 weeks. These areas were stained intensely with Alizarin Red, indicating calcification of the extracellular matrix (Fig. 3BII). ASCs grown in the control medium did not stain with Alizarin Red (Fig. 3BI).

Osteogenic differentiation of ASCs from BFP. (
Protein expression of osteocalcin, a marker characteristic of mature osteoblasts and osteocytes, was determined by immunofluorescence. ASCs cultured in OM, but not undifferentiated ASCs, showed osteocalcin expression (Fig. 3CI, CII), as did SAOS-2 cells (positive control) (Fig. 3CIII). During the differentiation process, expression of the osteogenic genes CBFA1 and osteonectin (SPARC) increased by eight- and twofold, respectively, at day 14 of culture. On the other hand, we did not observe a decrease in gene expression of the adipocyte marker PPARγ until day 21 (Fig. 3D). SAOS-2 cells were used as positive control. These data demonstrate that ASCs from BFP are capable of osteogenic differentiation in vitro.
Adipogenesis
After adipogenic induction of ASCs, we observed intracellular lipid vacuoles, which increased in size and number during culture (Fig. 4A). Oil Red staining revealed multiple intracellular lipid filled droplets in ASCs cultured in AM, but not in undifferentiated ASCs. The expression levels of the specific adipocyte marker PPARγ and GAPDH were measured by quantitative real-time (RT)-PCR using the comparative Ct method (Fig. 4B). PPARγ expression increased during culture, reaching approximately fourfold induction compared to undifferentiated ASCs. Whole adipose tissue samples served as positive control.
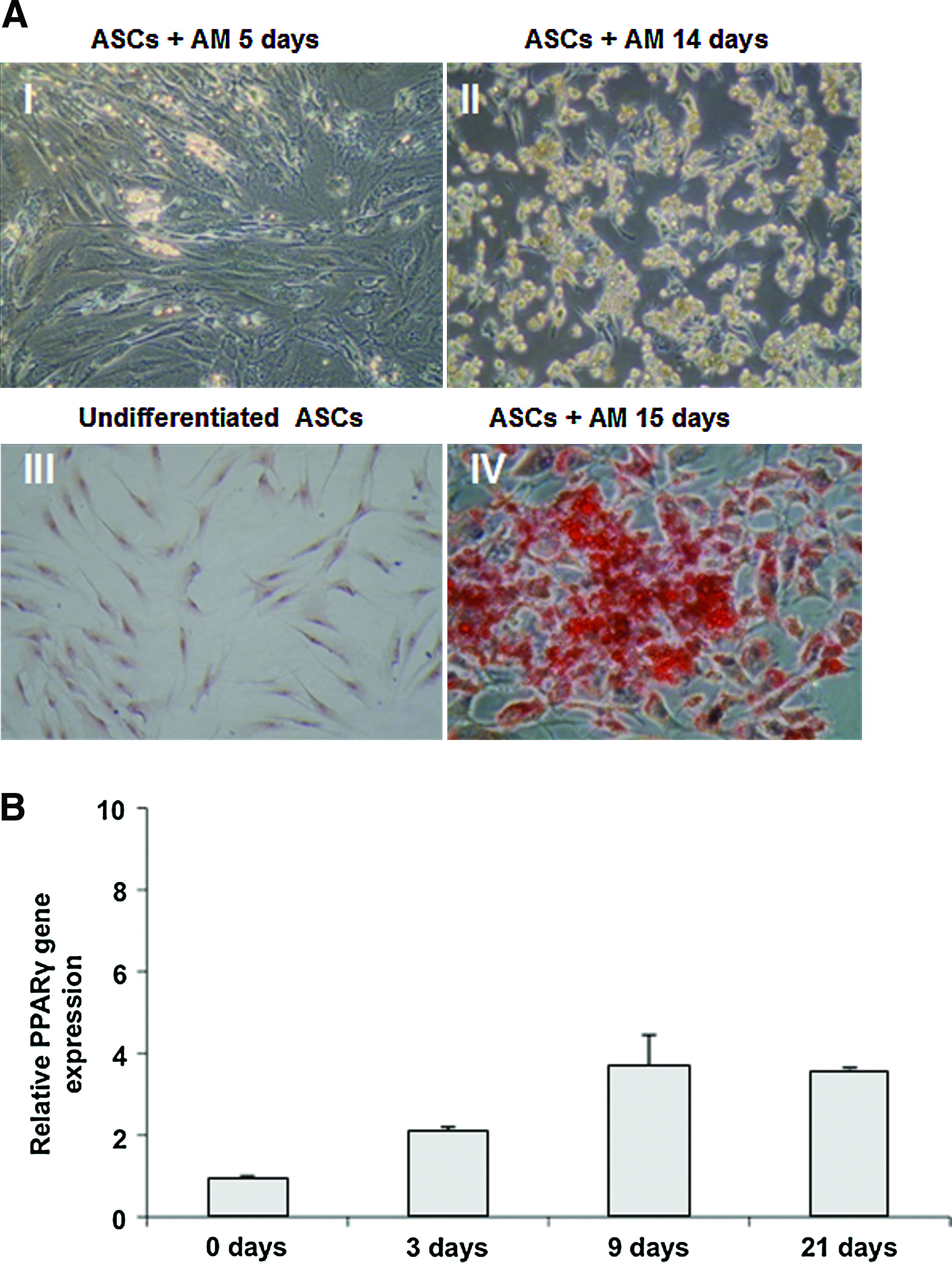
Adipogenic differentiation of ASCs from BFP. (
Chondrogenesis
ASCs from BFP synthesized cartilage matrix molecules, and produced an extracellular matrix characteristic for chondrocytes when grown in CM. After ∼5 days of culture in CM, ASCs changed their morphology to more spheroid shaped (Fig. 5AI). ASCs under control conditions did not show TB staining at pH 2, which is specific for sulfated proteoglycans in cartilage matrix (Fig. 5AII). The number of nodules that stained positive with TB (Fig. 5AIII) increased during 4 weeks of culture in CM (data not shown). TB-stained purple nodules were not seen in undifferentiated ASCs (Fig. 5AIV), but they were clearly visible after culture in CM (Fig. 5AV). Immunohistochemical analysis did not show expression of collagen type II in undifferentiated cells (Fig. 5BI), but it was expressed by cells grown in micromass in CM after 4 weeks (Fig. 5BII). Expression of the master chondrogenic factor SOX9 was increased by 3.4-fold in ASCs grown in CM compared to undifferentiated ASCs at day 14 (Fig. 5C). Expression of the adipocyte gene PPARγ decreased during chondrogenic differentiation (Fig. 5C).
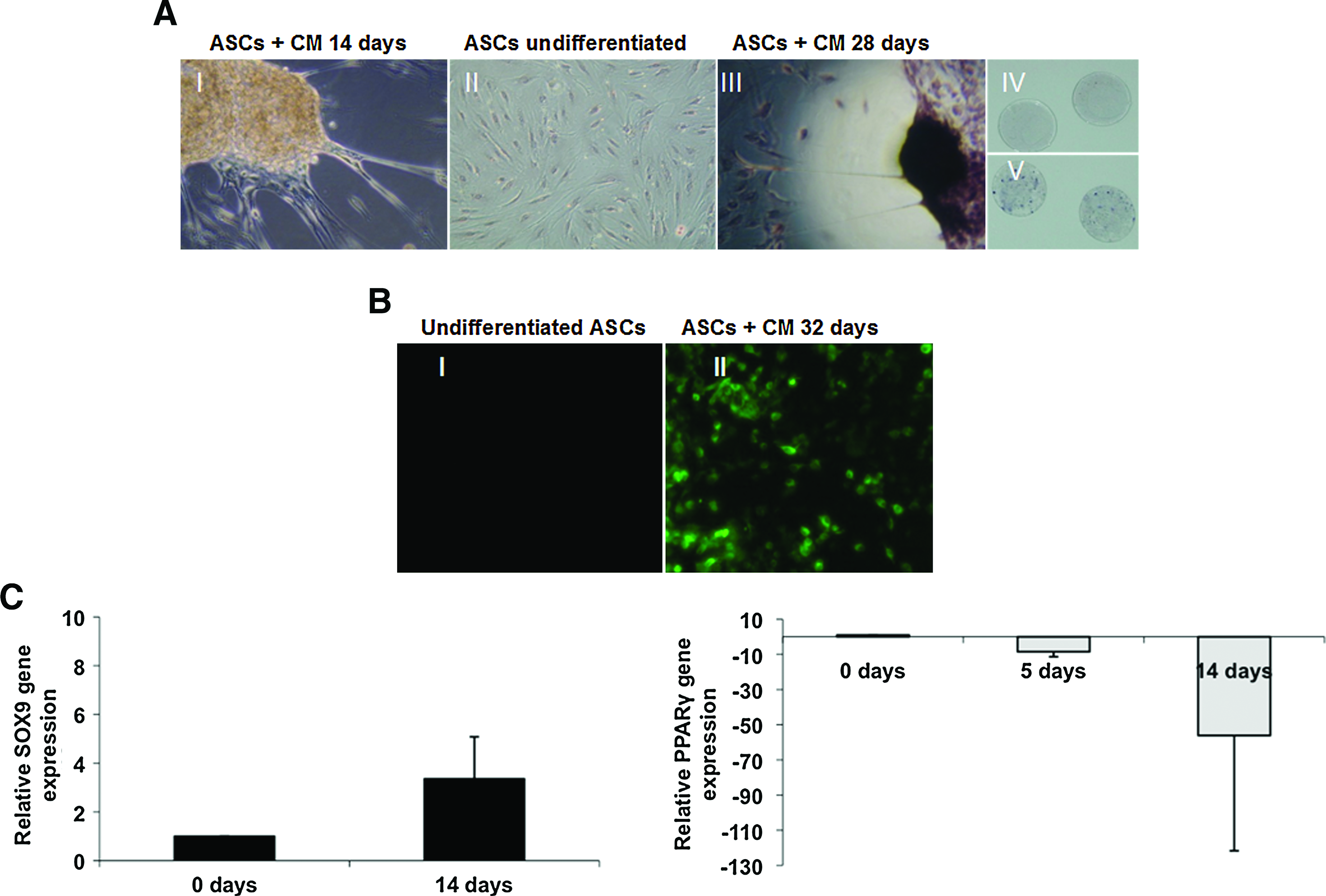
ASCs from BFP cultured in CM. (
Discussion
Adult stem cells are present in various organs and develop important functions in tissue maintenance and homeostasis. 32 One major goal of tissue engineering medicine is to find a source that can provide an adequate number of stem cells for clinical application that implies minimal morbidity. Adipose tissue holds great promise in regenerative medicine; that is, it is available in large quantities as waste material, and it contains more progenitor cells giving rise to different cell populations than bone marrow. 33 Unlike subcutaneous fat, BFP is a specialized mass of adipose tissue considered an ideal flap for oral surgery; since it is easy to harvest and reliable, it contains a rich blood supply, and its harvesting causes minimal donor-site morbidity and low complication.34,35 The size of BFP appears to be similar among different persons and independent of their body weight and fat distribution. 36 Patients with little subcutaneous fat have BFP with normal weight and volume. The results of our study are in agreement with published results showing that the size of BFP is not related to a person's general adiposity. 36
In this study we aimed to assess the presence of ASCs in BFP using flow cytometry, to compare the number of ASCs present in BFP with that in adipose tissue obtained from another fat depot (SC), to compare the cell marker expression in BFP with that in SC, and to determine the multipotent potential of ASCs from BFP in vitro. Our results showed that BFP contains a high proportion of cells with ASC characteristics that can be easily isolated and expanded in vitro. Approximately 30% of SVF from BFP express characteristic markers of fresh ASCs, that is, CD90, CD73, CD29, and CD34.27,37 The percentage of ASCs from BFP obtained in our study (∼30%) is slightly higher than that obtained from abdominal subcutaneous adipose tissue (∼22%). 38 In addition, the occurrence of MSC in bone marrow tissue is <0.01%, 3 which makes BFP an excellent source of fresh ASCs, avoiding the need for in vitro expansion, high costs, and contamination risk.
CD34 expression is characteristic of fresh ASCs, 39 and its expression decreases with passaging in ASCs. Expression of CD34 in these cells contrasts with MSCs from bone marrow, which lacks expression of this marker. 40 CD34+ cells are capable of stimulating angiogenesis, and they are involved in neovascularization processes that facilitate healing of ischemic tissues.41,42 The in vitro expansion of cells from SVF causes changes in the phenotype and characteristics of these cells such as loss of CD34 expression. 43 We hypothesize that CD34 expression might be related to differences in cell properties and differentiation potential. Freshly isolated ASCs from BFP lack expression of CD105, but expression of this marker increases rapidly after seeding, as has been described by others using ASCs from other fat depots.30,44 ASCs also lack expression of CD146, a characteristic marker of endothelial cells as well as vascular smooth muscle cells. 27 In our study we found a small population of CD146+ cells in the first passages (up to P2) of ASCs from BFP. The presence of this CD146+ contaminating population, as well as the presence of CD34+ cells, might be due to the highly enriched blood vessel supply in BFP. 45 This could be related to the excellent wound healing properties of BFP as a pedicled graft in oral surgery for treatment of oroantral communications,46,47 maxillary defects, 34 oral submucous fibrosis, 48 and vocal cord defects. 49
It was expected that differences in the site of origin of the adipose tissue is related to different ratios of cell types such as epithelial progenitor cells, fibroblasts, pericytes, myoblasts, and the like. We evaluated differences between SC and BFP, a mass of adipose tissue considered a marker of visceral fat. 18 We observed differences between BFP and subcutaneous adipose tissue regarding CD34 expression. This difference might be attributed to the higher blood supply of visceral tissue, which might contribute to the heterogeneity of the cell population, which in turn might increase the osteogenic potential. 50 The CD34+ cells obtained from dental germ pulp also have the ability to differentiate to preosteoblasts, form bone nodules after implantation in mice, and show angiogenic potential. 51 We found higher CD34 expression in passaged ASCs from BFP compared with passaged ASCs from subcutaneous adipose tissue, which could make them more suitable for bone engineering therapies.
After 1 week of culture, we observed that the number of BFP-derived ASCs was twofold higher than the number of ASCs in SC. This difference might be due to the differences in age of the patients, 52 although this difference might also be attributed to the intrinsic characteristics of the patients or to other particular properties of the adipose source. 53
When ASCs from BFP were cultured in AM for 2 weeks, more than 90% of the cells accumulated lipid vacuoles and expressed the characteristic adipogenic gene PPARγ, which is considered a master regulator of adipogenic differentiation. 54 However, we observed constitutive expression of this adipocyte gene in noninduced ASCs, similar as in other studies. 55 Osteogenesis occurred in the presence of OM within 1 week as shown by increased ALP activity. 1,25-Dihydroxyvitamin D3 plays an important role in bone formation and maturation by increasing Ca2+ concentration at the mineralization site. It promotes early differentiation of progenitor cells to osteoblastic commitment. We, as well as others, 56 found that the combination of 1,25-dihydroxyvitamin D3 with ascorbate-2-phosphate and β-glycerophosphate induces matrix calcification as shown by Alizarin Red staining and osteocalcin expression. Osteocalcin is an osteogenic marker indicative of the final mature stages of osteoblastic differentiation and is expressed when mineralization begins. 57 In addition to the increased ALP activity and matrix calcification, expression of the osteogenic genes CBFA1 and osteonectin confirmed osteogenic differentiation of ASCs from BFP. No increase in PPARγ was observed during culture. After exposing ASCs to CM for 4 weeks, we observed TB-stained nodules indicative of a proteoglycan matrix characteristic for cartilage. Expression of collagen II, a marker believed to be specific for articular cartilage, was observed by immunohistochemistry in differentiated ASCs. Increased expression of the master chondrogenic factor SOX9 was observed in ASCs from BFP at 14 days of culture, followed by decreased expression of the adipogenic marker PPARγ. ASCs could represent a valuable tool for cartilage repair that might overcome the limited intrinsic repair capacity of this avascular connective tissue, which most orthopedic patients complain about. Flow cytometry analysis of marker expression in ASCs before differentiation showed low expression of hematopoietic markers CD45, CD19, CD14, and HLADR, as well as low expression of endothelial markers CD146 and SSEA, which demonstrates that cross-contamination with hematopoietic or endothelial cells did not occur.58,59
The ideal source of stem cells for tissue engineering would be one that can be easily harvested, and contains a high number of expandable stem cells. 60 This study shows that SVF isolated from BFP contains ∼30% ASCs with high expression of the angiogenic marker CD34. Under appropriate conditions, ASCs from BFP differentiate to chondrocytes, osteoblasts, or adipocytes, suggesting that BFP can be a rich alternative source of stem cells, readily available for clinical application in the field of tissue engineering for the repair of bone and cartilage defects.
Footnotes
Acknowledgments
We wish to extend our thanks to the surgical staff of the Hospital Clínic and Hospital Sant Joan de Déu for facilitating the donation of human adipose tissue for this study. We also thank the staff of the Flow Cytometry research core, in Institut de Biotecnologia i Biomedicina, Universitat Autònoma de Barcelona (UAB), for their assistance with the flow cytometry studies, and the Skeletal Tissue Engineering Group Amsterdam (STEGA) at the VU University Amsterdam, The Netherlands, for their excellent support with the real-time PCR analysis. This work was funded by the Faculty of Dentistry, International University of Catalonia. E. Farré Guasch received a fellowship from International University of Catalonia.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.