
Abstract
The ideal bioartificial liver should be designed to reproduce as nearly as possible in vitro the habitat that hepatic cells find in vivo. In the present work, we investigated the in vitro perfusion condition with a view to improving the hepatic differentiation of pluripotent human liver stem cells (HLSCs) from adult liver. Tissue engineering strategies based on the cocultivation of HLSCs with hepatic stellate cells (ITO) and with several combinations of medium were applied to improve viability and differentiation. A mathematical model estimated the best flow rate for perfused cultures lasting up to 7 days. Morphological and functional assays were performed. Morphological analyses confirmed that a flow of perfusion medium (assured by the bioreactor system) enabled the in vitro organization of the cells into liver clusters even in the deeper levels of the sponge. Our results showed that, when cocultured with ITO using stem cell medium, HLSCs synthesized a large amount of albumin and the MTT test confirmed an improvement in cell proliferation. In conclusion, this study shows that our in vitro cell conditions promote the formation of clusters of HLSCs and enhance the functional differentiation into a mature hepatic population.
Introduction
For the time being, however, such supports can only temporarily replace a subset of essential liver functions; the most important challenge concerns how to maintain viable and functional hepatocytes outside the native liver environment. 6 The use of bioreactors is a fundamental requirement because of the high metabolic activity of liver cells. Several hepatocyte bioreactors, based on different configurations, have been developed and most attempts were focused on adapting bioreactor technologies to the development of an extracorporeal hybrid liver support device.2,7 Despite recent advances, isolated hepatocytes usually begin to proliferate under suboptimal culture conditions, but they have the tendency to lose most of the characteristic differentiated hepatic functions progressively, dying within 3–4 days.8,9
In an effort to overcome this limitation, which relates to the marked specialization of mature hepatocytes, several authors turned their attention to stem cell biology. Herrera et al. were recently successful in isolating a population of adult human liver stem cells (HLSCs). 10 These HLSCs differentiated into mature hepatocytes and in vivo they contributed to regenerating the liver parenchyma in severe-combined immunodeficient mice. In light of these results, obtained with classical cell biology techniques (i.e., monolayer and static growth conditions), we applied advanced tissue engineering strategies based on the use of a bioreactor to create flow of perfusion medium through the scaffolds and on the presence of supporting cells (such as hepatic stellate cells [ITO] cells) in HLSC cultures in an effort to create a more natural hepatic milieu in vitro.
Material and Methods
Perfusion bioreactor: Mathematical model
We used a perfusion bioreactor in which eight porous scaffolds can be cultured in eight independent chambers. In each chamber the medium flows from top to bottom through a three-dimensional (3D) scaffold. HLSC growth in each scaffold was described using a mathematical model. The model applied in this work was derived from our previous work. 11 Briefly, the geometry of a highly porous scaffold, as the collagen sponge (Avitene Ultrafoam Collagen Hemostat; Davol Inc., Cranston, RI) used in the experiment (Fig. 1a), was approximated to a cylinder with parallel cylindrical channels and no interconnections (Fig. 1b).
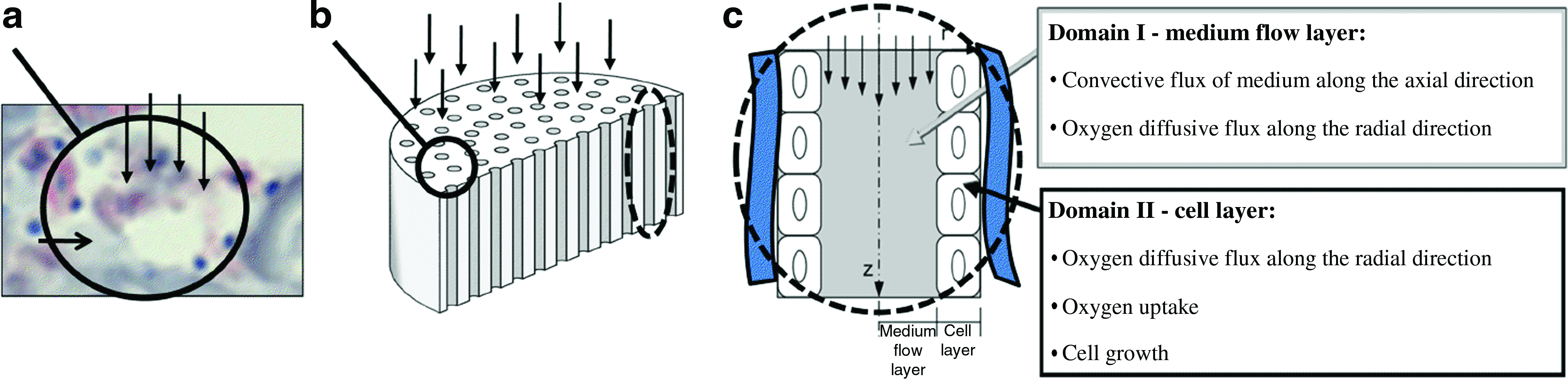
Bioreactor model. (
A channel diameter of 200 μm was the mean value used for the distribution, based on microscopic evidence and image analysis of the collagen scaffolds, whereas the total scaffold porosity was the maximum available for this geometry.
The model contains a uniform cell layer (Domain II) adhering to the channel wall while the medium flows through the central portion of the channel (Domain I) as exemplified in Figure 1c. The thickness of the cell layer increases as the cells grow, while the axial velocity of the medium, vz, depends on the total flow rate and the diameter of Domain I.
Oxygen was chosen as the reference species because its concentration strongly affects cell growth. 12
The phenomena governing oxygen concentration for each layer, and consequently also cell growth, are summarized in Figure 1c. Assuming that none of the phenomena depend on angular coordinates, the governing equations were derived for cylindrical geometry in term of axial and radial coordinates as follows:
Domain I: medium flow layer
where
Domain II: cell layer
where
F is a inhibitory function that stops the cell growth when oxygen concentration is below the physiological limit
The oxygen concentration at each channel inlet (
The system of equations (1), (4), and (7) was solved using MATLAB (The MathWorks, Natick, MA) by means of the finite volume method. Every channel was discretized in the axial and radial directions.
Cell culture and differentiation protocol
Human liver stem cells
These cells were isolated, purified, and characterized as described by Herrera et al. 10 Briefly, HLSCs were obtained by culturing cryopreserved hepatocytes obtained from Lonza Group Ltd., Basel, Switzerland (IRB approval is not required).
The cryopreserved cell population was quickly thawed in a 37.1°C water bath with gentle shaking, and then it was seeded in a flask for cell expansion. Cell viability was determined using Trypan blue exclusion staining. Characterization was done as described by Herrera et al. 10 Briefly, immunofluorescence was used to confirm HLSCs purity in cell culture on chamber slides. The absence of CD34, c-kit, and CK19 or markers for nonparenchymal liver cells (alpha-smooth muscle actin [SMA]) confirmed their purity. These assays were randomly performed in triplicate for the cryopreserved vials supplied. HLSCs purity was 98%.
HLSCs were expanded in a flask in the presence of hepatic cell medium (HCm): α-minimum essential medium/endothelial cell basal medium-1 (3:1) (from Gibco, Carlsbad, CA/Cambrex, Charles City, IA) supplemented with
ITO cells
These cells were isolated and characterized using a two-step collagenase perfusion as described by Paik and colleagues. 13 Briefly, ITO cells were obtained by culturing cryopreserved cells obtained from Dr. M. Pinzani's Lab, University of Florence, Florence, Italy (IRB approval is not required).
Collected ITO cells were suspended at a concentration of 1 × 105 cells/mL in Dulbecco's modified Eagle's medium supplemented with fetal calf serum (20%), penicillin (50 IU/mL), and streptomycin (50 μg/mL) (all from Sigma) and cultured on well plates coated with collagen type I (10 μg/mL) (from Sigma) in a 5% CO2 humidified atmosphere.
3D cultures
For all the differentiation experiments, 3D cultures were conducted in a perfusion bioreactor according to the protocol published elsewhere. 14 Briefly, cells between passages 3 and 8 were detached from the flask surface with trypsin-EDTA, washed, and counted; then 5 × 105 cells were seeded onto a collagen sponge15,16 (Avitene® Ultrafoam™Collagen Hemostat; Davol, Inc.) with diameter 5 mm and thickness 3 mm. Up to eight sponges were seeded for each experiment.
On the day after seeding, the sponges were placed, one in each separate chamber, in the bioreactor, which assured a perfusion medium flow rate of 0.5 mL/min through each scaffold. 14
The differentiated conditions used in the bioreactor are shown in Table 2.
HLSCs, human liver stem cells; ITO, hepatic stellate cells.
All cultures were performed in triplicate; the scaffolds were analyzed at fixed time points, in terms of their biochemical parameters on days 1, 3, 5 and 7, and their morphological features on days 3 and 7.
MTT assay
Cell proliferation rates were analyzed by the MTT (3-4,5-dimethylthiazol-2yl-2,5-diphenyltetrazolium bromide)-based cytotoxicity test using the Denizot and Lang method with minor modifications. 17 3D cell cultures removed from the bioreactor were placed on a culture plate with 1 mL of MTT solution (0.8 mg/mL in phosphate-buffered saline). Cultures were returned to the incubator and the supernatant was discarded after 3 h. Each scaffold was then transferred to an Eppendorf microtube, and 1 mL of extraction solution (0.01 N of HCl in isopropanol) was added. The Eppendorf microtubes were vortexed for 5 min to enable total color release from the scaffolds and then centrifuged at 15,000 g for 5 min, and the optical density of the supernatants was read at 540 nm.
Albumin ELISA assay
Albumin secretion from the HLSCs was determined using ELISA. Samples were harvested daily from the circulating medium and frozen at −20°C. The albumin concentration was measured using the human albumin AssayMax Human Albumin ELISA (Gentaur, Brussels, Belgium). The minimum detectable dose of albumin with this kit is typically 100 ng/mL.
Histological and morphological analyses
Specimens were fixed in formalin, embedded in paraffin, and stained with hematoxylin and eosin. Cryostatic sections of 7 μm were used for morphological and immunohistochemical analyses. They were layered over gelatine-coated glass slides, fixed with absolute acetone for 10 min at room temperature, and cryopreserved at −20°C until use. Ck19 on HLSCs was identified using the acid phosphatase anti-acid phosphatase procedure. Reactions were all conducted in humid chambers at room temperature. Briefly, after saturating nonspecific antigen sites with 1:20 rabbit serum in 0.05 M Maleate TRIZMA (pH 7.6; Sigma) for 20 min, the first antibody was added to samples (1:20 mouse anti-human CK19; Dako, Milano, Italy). After incubation for 2 h, the samples were rinsed with buffer solution, and the second antibody was added for 30 min (Link Ab [Dako], rabbit anti-mouse). After rinsing again, the sections were incubated for 30 min with 1:50 mouse acid phosphatase anti-acid phosphatase Ab (Dako), then rinsed once more, and finally, reacted for 20 min with Fast Red Substrate (Sigma). Counter staining was done with hematoxylin (Sigma).
Statistical analyses
Values are given as means ± standard deviation. The one-way analysis of variance in the Excel software package was used for data analyses. Statistical significance was assumed when p < 0.05.
Results
Bioreactor model
The bioreactor model was developed as reported in the Material and Methods section (Fig. 1). We considered the time-dependent profile of cell volume and oxygen concentration along each channel as the main output of the system. Different flow rates have been used in the model: Figures 2 and 3 show the results for three of them, that is, 0.1, 0.5, and 2.5 mL/min. The results obtained with the model confirmed that cell growth depends on the flow rate of the medium. Using the lowest flow rate (0.1 mL/min) would enable a rapid cell growth at the channel inlet (Fig. 2, continuous line) where the oxygen concentration remains higher than 80% of the saturation value. On the other hand, a flow rate of 0.1 mL/min would not be able to avoid an oxygen gradient forming along the length of the channel: the O2 concentration would decrease moving from the inlet to the outlet of the channel. The concentration at the outlet of the channel (dotted and dashed line in Fig. 2) would drop to the near-critical level of 5.26e−2 mol/m3 after 7 days of culture. This oxygen gradient would lead to a slower cell growth rate in the central and bottom portions of the scaffold (dashed lines, and dotted and dashed lines) and consequently to an uneven distribution of the cell volume in the z-direction.

Time variation of the percentage of volume occupied by the cell (upper line) and the O2 concentration related to the saturation value (bottom line). Data are given for three flow rates, that is, 0.1 mL/min (left), 0.5 mL/min (center), and 2.5 mL/min (right). Each graph shows the data for three z-coordinates, that is, at each channel's inlet (continuous line), midline (dashed line), and outlet (dashed and dotted line). The horizontal dotted line in the O2 graphs corresponds to the threshold oxygen concentration of 5.26e−2 mol/m3.
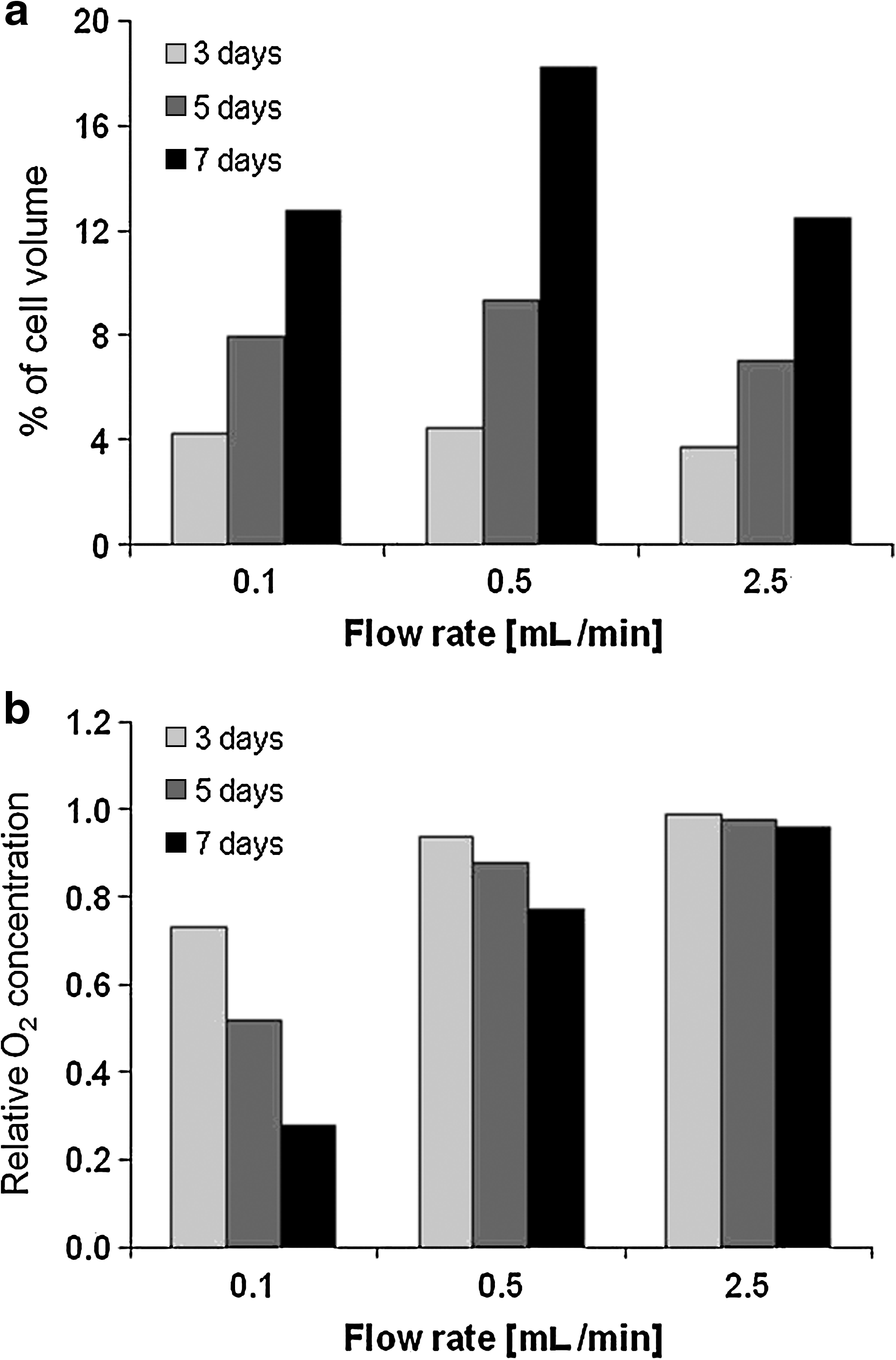
Percentage of mean cell volume (
A flow rate of 0.5 mL/min would be able to maintain the oxygen concentration above 80% of the saturation value through the channel. However, the oxygen gradient would be still present since few days of culture and it would correlate closely with a gradient in the percentage cell volume.
Increasing the flow rate would lead to a more homogeneous cell volume and oxygen concentration along the z-direction (Fig. 2). However, it also induced a high shear stress on the cell surface, which could reduce the cell growth rate (Fig. 3).
Figure 3 shows the prediction model for the mean percentage of volume occupied by the cells in the whole scaffold for three time points and the three flow rates. For short-term hepatocyte culture, a low flow rate (around 0.5 mL/min and below) could give rise to the highest percentage cell volume because the mean oxygen concentration does not strongly depend on the flow rate when the cell volume is low. For longer-term culture (5 days or more), an optimal flow rate (around 0.5 mL/min) achieved the highest mean percentage cell volume. Still, as shown in Figure 2, a uniform cell growth in the z-direction can only be obtained with a flow rate of 2.5 mL/min or more.
We considered a flow rate of 0.5 mL/min as the best choice for maintaining high cell growth rate with moderate heterogeneity in the cell distribution.
Morphological analyses
Figure 4 (left column) shows the morphological features of the HLSCs cultivated in the presence of HCm on day 3, in the previously described dynamic cell culture conditions (0.5 mL/min). In these conditions, the cells have clearly become organized into clusters inside the small compartments of the sponge. The cells are able to retain their phenotype, as confirmed by the red staining (showing positivity) for the biliary marker (CK19) and by the presence of double-nucleated cells (cells inside the circle). The direction of the flow of medium is indicated by white arrows, while the presence of biomaterial fibers (horizontal black arrows) indicates the organization of the small compartments in which the HLSCs could become organized into liver-like tissue.

Morphological analyses of HLSCs on a three-dimensional scaffold in different flow conditions on days 3 (left) and 7 (right). Cells are stained with classical H&E and biliary markers (CK19) (red cells). White arrows: direction of flow of medium; horizontal black arrows: collagen fibers. Circles: double nucleated cells. HLSC, human liver stem cell; H&E, hematoxylin and eosin. Color images available online at
Figure 4 (right column) shows the 7-day cell cultures. Here again, the cells were stained with classic hematoxylin and eosin and immunostained for biliary markers (CK19; red cells). The cultured cells initially seeded on the surface coinciding with the inlet for the flow of medium (white arrows) soon started to become organized into multiple layers, migrating through the collagen sponge (horizontal arrows). By day 7 (Fig. 4), some clusters appeared to be trapped inside the sponge (red cluster), showing layered cells in the holes in the sponge. The differentiated phenotype showed a “hexagonal shape” often associated with double nuclei (cells inside the circle) in contrast to the flat shape initially seen, characteristic of monolayer cultures (data not shown).
Proliferation test
Cells seeded onto the sponge were cultivated inside the bioreactor for up to 7 days under different conditions, based on the type of medium (hepatic cell selective, stem cell selective, or both), the type of cell (HLSC, ITO, or both), and combinations of the two (see Material and Methods section).
Biological parameters were tested on days 1, 3, 5, and 7, for example, the MTT was used to detect the proliferation rate and albumin was quantified to establish differentiation status.
The proliferation rate (as detected with the MTT) is shown in Figure 5a, where it is clear that cells survived up to day 7 in all culture conditions (i.e., there was no drop in MTT values), confirming the good functionality of the bioreactor. In more detailed analyses, we found there was no detectable increase in the MTT value during the days of culture when HLSCs were cultured alone; a slight increase was only visible when SCm was used. A higher increase in MTT values over time was observable in scaffolds containing ITO cells alone or HLSC and ITO cocultures, irrespective of the combination of perfusion media used. The greatest improvement in cell proliferation rate was obtained when HLSC and ITO cocultures were run in the presence of SCm. The proliferation rate of ITO cultures showed a marked increase in the MTT result, characteristic of the biology of nonparenchymal cell types.

(
Albumin production
Having established that a continuous flow of medium through the bioreactor is able to sustain cell viability, we investigated liver functionality. Secreted albumin levels for each of the above-described culture conditions were determined by ELISA in samples of medium. Extracted DNA values from the cellularized scaffolds in the different culture conditions were used to normalize all functional results. Results in Figure 5b confirm that albumin was synthesized by the liver cells grown in the bioreactors throughout the experiment. We compared the ability of HLSCs to maintain protein synthesis in the different culture conditions in the perfusion bioreactors (Fig. 5b). HLSCs cultured alone showed an albumin production strongly related to the medium involved, the presence of SCm giving rise to a higher production. The same trend was evident in coculture, where a higher albumin secretion again coincided with the presence of SCm medium. There was naturally no albumin production detectable in the culture containing ITO (nonparenchymal cells) alone.
Discussion
With a view to develop an efficient differentiation method to improve the commitment of adult precursor liver cells to mature hepatic cells, we optimized the culture method by using a 3D culture system in a perfusion bioreactor and testing several combinations of cell cultures (HLSC, ITO, and both) and media (HCm, SCm, and both). The first issue to consider was the well-known biological behavior of primary hepatocytes 18 : because of their high metabolic activity in vivo, their survival relies on an extensive vascularization to ensure an adequate nutrient and waste exchange.19–21 With this in mind, our experiments were conducted in dynamic culture conditions using a perfusion bioreactor. This system enabled us to simultaneously and separately culture eight porous scaffolds, ensuring an accurate control of the flow rate of medium through each scaffold. 14
The results obtained with the mathematical model were used to decide a flow rate a priori, which was set to 0.5 mL/min for up to 7 days. In these conditions, HLSCs in the presence of HCm not only showed a good proliferation rate (Fig. 5a, first histogram group from the left) and a marked clustering within the collagen scaffolds (horizontal black arrows in Fig. 4), but also a good phenotype resembling mature hepatocytes, confirmed by positivity for a biliary marker such as CK19 (red cells in Fig. 4), the presence of double-nucleated cells (cells inside the circle in Fig. 4), and albumin secretion (Fig. 5b).
Encouraged by these results, we focused on developing a system to improve the proliferation rate and differentiated status of the HLSC population. Our strategy was based on using a commercial SCm capable of influencing the stem cell biology of the undifferentiated cells in the HLSC population and cocultivating the HLSCs with a supporting cell population (ITO); the latter solution was prompted by evidence that 3D coculture models involving parenchymal and nonparenchymal cells can mimic the plate organization of the liver parenchyma, holding promise for the reconstitution of the structure and matrix geometry of the hepatic microenvironment. Interaction between HLSCs and ITO cells may have an important role, for instance, in modulating cell growth, migration, and differentiation.22–26
The cultures obtained with different combinations of cells and media were analyzed according to their proliferation rate and their ability to secrete albumin (Fig. 5).
In detail, our results could be explained in the following text.
Hepatic cell medium
This medium induces a lower proliferation rate and smaller albumin production in all cell cultures. No albumin production is detectable on ITO cultures (Fig. 5b), which shows that there are no hepatic progenitors in the ITO cell population. The coculture system with HLSCs and ITO cells showed the highest proliferation rate and greatest albumin production (p < 0.05 compared with HLSCs alone at the same time point). This might be due to a positive interaction between the stem cell population inside the HLSCs and the ITO cells.
Stem cell medium
In HLSC cultures, this medium induced a small increase in the MTT value by comparison both with different time points in the same culture and with the same time points in culture with HCm (p < 0.05; Fig. 5a). The same trend was seen in the effect on albumin production, but the differences were much more evident (p < 0.01 compared with HLSCs in HCm; Fig. 5b); this could be due to the activation of the undifferentiated stem cell population in the HLSCs. There was a clearly evident, considerable increase in the MTT value for the ITO cultures (p < 0.01); as mentioned previously, ITO cells are unable to produce albumin. In cocultures (HLSC and ITO), the presence of SCm induced a higher proliferation rate and a greater albumin production (p < 0.001 compared with HLSCs alone in HCm), probably because of a positive interaction between HLSCs and ITO cells.
Stem cell and hepatic media
This medium had a better effect on differentiation than on proliferation: the HLSC proliferation rate in the coculture stopped around day 5, whereas albumin was also produced during the last days of culture, with rising albumin concentrations. As in the other two cases, when this medium was used the best results in terms of MTT value and albumin production were obtained with the HLSC and ITO coculture.
In this particular case, ITO cells proved capable of enhancing the differentiation of precursor hepatic progenitors. Our data showed that the hepatocytes exhibited a differentiated morphology and much higher levels of mature functions, such as albumin secretion, when compared with the monoculture system (in which only hepatocytes were cultured) in the same condition. In addition, the higher values of the MTT test (Fig. 5a) of cocultures compared with the results of hepatocytes or ITO cells alone, independent of the medium, indirectly support our hypothesis.
However, as revealed by proliferation rate, it is evident that cells survived up to day 7 in every culture condition; then a tendency for the initial tissue-like structures to degrade started. This event could be likely related to the progressive death of ITO cells, unable to further support hepatocytes. Thus, we came to the conclusion that ITO cells are initially able to modulate the differentiation of HLSCs in cocultures either by direct cell contacts or via soluble factors in the medium.
In the light of these results, we can confirm that a combination of the SCm with a coculture including ITO cells definitely increased both cell proliferation and cell maturation.
In conclusion, HLSCs were induced to proliferate as cells with hepatic characteristics by creating and enhancing a mutual synergism with the medium and with neighboring nonparenchymal cells.
Our findings demonstrate the feasibility of this protocol for the differentiation and amplification of HLSCs as a useful source for cell transplantation as well as for use in pharmaceutical and toxicological investigations.
Footnotes
Acknowledgments
This study received partial funding from the University of Padova, Italy, and a grant from Fresenius Medical Care. The authors thank Professor Massimo Pinzani for kindly providing ITO cells.
Disclosure Statement
M.B. Herrera is the named inventor on a related patent application concerning liver stem cells. C. Tetta is a full-time employee of Fresenius Medical Care (Deutschland). U. Cillo has a financial interest in Fresenius Medical Care.