
Abstract
Astrocytes are a major transplant cell population to promote neural repair in a range of pathological conditions. In this context, the development of robust methods to label neural transplant populations (for subsequent detection and cell tracking in vivo) is key for translational applications. Magnetic iron oxide nanoparticles (MNP)-based vector systems offer a range of advantages for neural cell transplantation, notably, as contrast agents for magnetic resonance imaging, which allows for MNP-labeled cells to be detected using minimally invasive methods. Additionally, MNPs have other key features such as safety, the potential for linking with genetic material/drugs, and magnetic cell targeting. Therefore, MNPs can potentially be developed as a multipurpose nanoplatform for neural cell transplantation. The feasibility of labeling astrocytes derived for transplantation with MNPs has not been assessed to date. Here, we have established simple protocols to safely label astrocytes with MNPs; the survival and differentiation of labeled cells was assessed in three dimensional neural tissue arrays. Additionally, we have established the major mechanisms of MNP uptake by astrocytes.
Introduction
Recent advances in nanotechnology have highlighted the significant potential offered by delivery systems employing magnetic iron oxide nanoparticles (MNPs) for imaging applications in tissue engineering. Such particles have emerged as a major class of contrast agent for magnetic resonance imaging (MRI) with advances in chemical methods to control particle properties such as size, coatings, biodegradability, and toxicity. The ability to label cells with MNPs ex vivo therefore provides a technique to detect transplant cell populations in vivo using MRI 17 allowing for the noninvasive monitoring of the biodistribution, integration, and survival of transplanted cells. In this context, it has been demonstrated that neural progenitor/stem cells, including glial-restricted precursors, can be safely labeled with magnetic particles and tracked using MRI after transplantation into areas of pathology (with no demonstrable loss of functional capacity and limited effects on cell survival/differentiation).18–21 To date, however, the feasibility of using MNPs for cell labeling in astrocyte transplantation therapies has not been assessed. In addition to cell tracking, MNPs have applications in magnetic targeting (including magnetic cell targeting) and—because they can be readily functionalized with drugs/genetic material—they have the potential to serve as a multifunctional nanoplatform for cell transplantation applications. 22 We recently demonstrated, for the first time, that astrocytes derived from primary cultures can be transfected using MNPs in the presence of applied static/oscillating magnetic fields (the so-called magnetofection approaches), achieving efficiencies close to that of some viral vector systems. We further proved in this study that transfected cells could survive and integrate into host neural circuitry after transplantation into three-dimensional neural tissue arrays (organotypic cerebellar slice cultures). 22 These findings taken in conjunction with the multiple benefits of MNP vector systems led us to suggest that MNPs could represent the multifunctional vector system of choice for astrocyte transplantation therapies.
Here, we have established protocols to safely label astrocyte transplant populations with MNPs ex vivo for cell tracking applications. To achieve this, we have employed fluorescently labeled MNPs and astrocyte cultures to examine a range of parameters related to particle uptake and toxicity. We also demonstrate the survival and differentiation potential of astrocytes post-MNP labeling after transplantation into organotypic slice cultures derived from the rat cerebellum.
Materials and Methods
Astrocyte cell culture
Mixed glial cultures were established from cerebral cortices of neonatal Sprague-Dawley rats and astrocytes were purified using a published procedure. 23 The adherent astrocyte cell layer was trypsinized, and subcultured once in D-10 medium (Dulbecco's modified Eagle's medium plus 2 mM glutaMAX-I, 1 mM sodium pyruvate, 50 U/mL penicillin, 50 μg/mL streptomycin, and 10% fetal bovine serum) in poly-D-lysine (PDL)-coated T75 flasks. All cultures were incubated at 37°C in 5% CO2/95% air, with medium changes every 2–3 days, unless otherwise specified. Subconfluent astrocyte cultures were trypsinized, resuspended in D-10 medium, and used for MNP uptake, transplantation, and toxicity studies, as described below.
Cerebellar slice cultures
Brains of Sprague-Dawley rat pups at 10 postnatal days were collected into ice-cold slicing medium (Earle's balanced salts solution buffered with 25 mM HEPES). Cerebella were dissected, meninges removed, and 400 μm parasagittal slices prepared using a McIlwain tissue chopper. Slices were transferred to slicing medium at 4°C for 30 min, and then individual slices were transferred to pieces of Omnipore membrane on Millicell culture inserts (Millipore) in six-well plates containing the culture medium (50% minimum essential medium, 25% Earle's balanced salts solution, and 25% horse serum; supplemented with 1 mM glutaMAX-I, 36 mM D-glucose, 50 U/mL penicillin, and 50 μg/mL streptomycin). Slices were cultured at 37°C in 5% CO2/95% air, with medium changes every 2–3 days. All experiments involving animals were conducted in strict accordance with United Kingdom Home Office guidelines.
Uptake experiments
Paramagnetic, carboxyl-modified SPHERO Nile Red fluorescent magnetic particles (0.20–0.39 μm diameter, iron content 15%–20%) were obtained from Spherotech, Inc. These comprise a polystyrene core, stained with the fluorophore Nile Red, coated with a magnetite layer, and then overcoated with a functionalized monomer. For all incubations, unless specified otherwise, astrocytes were seeded into PDL-coated chamber slides (0.4 × 105 cells/cm2) and allowed to attach for 24 h before MNP addition in fresh D-10 medium. (A) For time course experiments, cells were incubated with 2–50 μg/mL MNPs for 1–24 h. (B) To determine the temperature dependence of uptake, cells were incubated with 20 μg/mL MNPs for 1 h at 4°C or 37°C (in 100% air); uptake at 37°C under these conditions was similar to that in incubations conducted in 5% CO2/95% air. (C) To determine mechanisms of MNP uptake, selective endocytosis inhibitors 24 (Sigma) were used at concentrations routinely employed for other cell types.24–26 Inhibitors comprised dynasore (80 μM) and tyrphostin 23 (350 μM) (inhibitors of clathrin-dependent endocytosis); filipin III (5 μg/mL) (inhibitor of caveolin-dependent endocytosis); and amiloride (1 mM) and 5-(N-ethyl-N-isopropyl) amiloride (EIPA; 100 μM) (macropinocytosis inhibitors); all stock solutions (×1000) were dissolved in dimethyl sulfoxide (DMSO) vehicle. Cells were preincubated for 20 min with inhibitor, and then with the same inhibitor plus MNPs (20 μg/mL) for 1 h. Controls included cells incubated with vehicle or medium alone (“no addition” control). The latter was included to verify that the vehicle (DMSO) was without effect on cells. (D) To determine the intracellular fate of MNPs by transmission electron microscopy (TEM), cells were seeded onto PDL-coated Aclar sheeting (0.6 × 105 cells/cm2). After 24 h, cells were incubated with 20 μg/mL MNPs for 24 h. (E) To determine the long-term disposition of particles (up to 3 weeks), cells were seeded onto PDL-coated glass coverslips or 12-well plates (0.1 × 105 cells/cm2). After 24 h, cells were incubated with either 0 or 20 μg/mL Spherofluor MNPs for 24 h. The medium was replaced with fresh D-10 minus MNPs and cells were cultured for up to a further 21 days. Cells were split (1 in 3) by trypsinization at weekly intervals. To terminate incubations, cells were washed three times with phosphate-buffered saline (PBS) then fixed, as described below.
Transplantation of labeled astrocytes into slice cultures
Astrocytes were seeded into PDL-coated plates (0.2 × 105 cells/cm2) and, after 24 h, incubated with fresh D-10 medium containing 20 μg/mL MNPs for 4 h. Medium was then replaced with fresh D-10 medium alone for 20 h. Nuclei were labeled by subsequent incubation of cells in medium containing 50 μg/mL 4,6-diamidino-2-phenylindole (DAPI) for 30 min, and then washed with PBS; this dual labeling procedure was used to robustly differentiate the transplant population from host astrocytes. Cells were trypsinized and resuspended at 4 × 107 cells/mL D-10 medium. Then, 0.25 μL of medium containing 10,000 cells was pipetted onto cerebellar slices that had been cultured for 9 days in vitro. To observe the passive spread of transplanted astrocytes due to the force of injection, immediately after application of cells (t = 0 h), slices were transferred to a slide, cells applied, and live slices observed using fluorescence microscopy. To monitor the survival and morphology of the trypsinized cells after transplantation and to confirm intracellular MNP localization, astrocyte transplant populations were additionally plated onto PDL-coated chamber slides. A total of 12 slices were examined.
Immunocytochemistry and histochemistry
Samples were fixed with 4% (w/v) paraformaldehyde (in PBS) for 20 min and then washed three times with PBS. For identification of astrocytes, polyclonal rabbit anti-glial fibrillary acidic protein (GFAP) antibody (DakoCytomation) was used. Fixed samples were incubated with blocking solution (5% normal donkey serum in PBS–0.3% Triton X-100) (room temperature [RT], 30 min), and then with primary antibody to GFAP (1:500 dilution in blocking solution; 4°C, overnight). After washing, samples were incubated with blocking solution (RT, 30 min), and then with fluorescein isothiocyanate-labeled donkey anti-rabbit IgG (Jackson ImmunoResearch Laboratoratories, Inc.; diluted 1:200 in blocking solution; RT for 2 h). Samples were then washed and usually mounted with Vectashield mounting medium containing DAPI (Vector Laboratories); for the slice cultures experiments, mounting medium without DAPI was used for cells and slices. For Perl's Prussian Blue staining to observe intracellular iron, fixed cells were incubated with 2% potassium ferricyanide in 2% HCl for 30 min, washed three times with distilled water, and mounted without DAPI.
Fluorescence microscopy and image analysis
Fluorescence microscopy was performed using an Axio Scope A1 microscope equipped with an Axio Cam ICc1 digital camera and AxioVision software (release 4.7.1; Carl Zeiss MicroImaging GmbH). Images were merged in Adobe Photoshop CS3 (version 10.0.1), and used for semiquantitative analysis, as described previously. 27 Purity of astrocyte cultures was assessed from merged images of GFAP and DAPI-stained cells (≥100 cells were scored per culture). To assess the effects of particles or endocytosis inhibitors on cell viability, counts of DAPI-positive nuclei were compared between control and treated wells with assessment of nuclear morphology; a minimum of three microscopic fields at ×400 magnification (≥100 nuclei in total) were assessed. Particle uptake by astrocytes (including endocytosis inhibitor experiments) was assessed from triple merges (×400 magnification) of DAPI, Nile Red, and GFAP fluorescent images, and the proportion of astrocytes (GFAP-positive cells) demonstrating particle uptake (termed % labeled cells) was calculated. To determine the extent of particle accumulation, labeled astrocytes were categorized based on the area of the cell occupied by the accumulated nanoparticles relative to the area of the cell nucleus; categories were low (≤10%), moderate (11%–50%), and high (≥51%). Labeled astrocytes were also classified with respect to the subcellular localization of nanoparticles; the proportion of labeled cells with a perinuclear particle localization either exclusively or combined with a cytoplasmic particle distribution is reported here. For all uptake calculations, a minimum of 100 GFAP-positive cells per treatment were evaluated. Confocal laser scanning microscopy, using a BioRad MRC1024 confocal microscope, was employed to determine if cell-associated particles had been internalized rather than simply adsorbed onto the cell surface.
Electron microscopy (scanning and transmission)
To assess particle size and shape, Spherofluor particles were placed in pure water, air-dried onto aluminum stubs, and observed uncoated using a high-resolution field emission scanning electron microscope (Hitachi S4500) operated at an accelerating voltage of 5 kV. To assess the intracellular localization of MNPs, samples were fixed with 2.5% (w/v) glutaraldehyde (in 0.1 M sodium cacodylate buffer containing 2 mM CaCl2; SCB) for 2 h, and then washed three times with SCB. Samples were postfixed with 1% osmium tetroxide in SCB for 1 h, washed, and then dehydrated in a graded series of ethanol, before infiltration in Spurr resin. After polymerization of the resin at 60°C for 16 h, ultrathin sections were cut on a Reichert Ultracut E microtome, and placed on 200 mesh thin bar copper grids and stained with 2% uranyl acetate in 70% ethanol and 2% Reynold's lead citrate. Sections were examined in a JEOL 1230 TEM operated at 100 kV.
MRI of agar-embedded MNPs
An initial proof of principle study was undertaken using a 1.5 T Philips Intera-Achivea MRI Scanner (Best) and a medium sense flex coil, to provide an assessment of the clinical feasibility of detecting MNP labeled astrocytes using clinical scanning methods. Spherofluor particle concentrations of 5, 20, and 50 μg/mL were embedded in agar gel; a test transfection-grade MNP with low iron content (0.5%, Neuromag; Oz Biosciences) was also included and a gradient echo sequence undertaken to qualitatively assess visibility.
Toxicity assays
Astrocytes were plated into PDL-coated 96-well plates (1 × 104 cells/cm2; 0.2 mL medium/well). After 24 h, the medium was replaced with fresh D-10 containing 0–50 μg/mL concentrations of Spherofluor MNPs, and cells were incubated at 37°C in a humidified atmosphere of 5% CO2/95% air for 24 h. The medium was then replaced with fresh D-10 alone, and the medium was changed every 2–3 days. MTS assays (CellTiter 96 AQueous One Solution Cell Proliferation Assay; Promega) were performed at 0–96 h postmedium change according to the supplier's instructions. Blank incubations, comprising medium plus MNPs but no cells, were run with each assay and the corresponding absorbance readings at 490 nm (A490) were subtracted from the appropriate test readings. Values were expressed as percentage cellular viability relative to control incubations minus MNPs.
Statistical analysis
Data are expressed as mean ± standard error of the mean. To assess the effects of MNP concentration and time of exposure on proportion of cells showing particle uptake, extent of particle accumulation, and degree of perinuclear localization, data were analyzed by a two-way analysis of variance followed by Bonferroni's post-tests; for clarity, comparisons versus the 2 μg/mL MNP group at a given time point (in the case of concentration dependence) and comparisons versus the same MNP concentration at 1 h (in the case of time dependence) only are displayed on bar charts. For temperature dependence of uptake, data were analyzed by Student's t-test (two-tailed). One-way analysis of variance, with either Bonferroni's multiple comparison test or Dunnett's multiple comparison test, was used to analyze data for the endocytosis inhibitor and cytotoxicity experiments, respectively. GraphPad Prism 4 for Windows software (version 4.03) was used for statistical analyses.
Results
Culture characteristics
Astrocyte cultures were of consistently high purity, as judged by GFAP immunostaining (97.8% ± 0.3%; n = 8 cultures). The majority of cells exhibited a flattened, polygonal appearance, characteristic of type 1 astrocytes, while cells with the complex, branching morphology of type 2 astrocytes accounted for <1% of the astrocyte population.
MNP uptake by astrocytes
Cell-associated fluorescent MNPs were clearly visible within both type 1 and type 2 astrocytes. Confocal microscopy revealed that fluorescence associated with MNPs and cytoskeletal GFAP was coincident; the former was only clearly visible in internal slices of cells (Fig. 1A and Supplementary Fig. S1, available online at
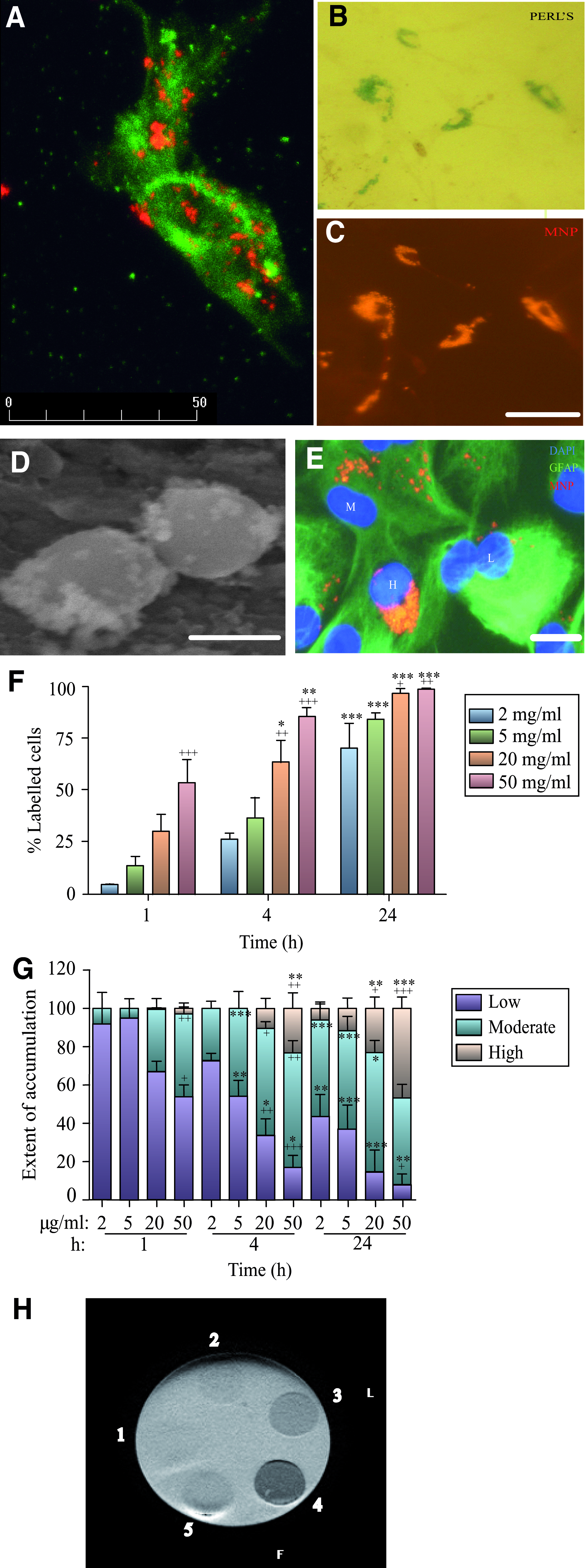
Uptake of magnetic iron oxide nanoparticles (MNPs) by rat astrocytes varies with particle concentration and length of incubation.
Astrocytes demonstrated clear heterogeneity in particle uptake (Fig. 1E); both the proportions of cells exhibiting MNP uptake and the extent of particle accumulation within cells showed time and concentration dependence (Fig. 1 F, G). Importantly, near-complete labeling of cells (>95%) was observed in incubations with ≥20 μg/mL MNPs for 24 h (Fig. 1F).
MRI of agar-embedded MNPs
Spherofluor particles embedded in agar gel at the higher concentrations studied (20 and 50 μg/mL) were readily observed using MRI. The lowest particle concentration and particles with low iron content yielded limited contrast. Preliminary findings indicate that the different concentrations resulted in ∼25%–75% reduction in T2* relaxation times compared with agar gel alone.
Perinuclear targeting of MNPs
Perinuclear clustering was a prominent feature after incubation of astrocytes with MNPs (Fig. 1E); presence of cytoplasmic and perinuclear collections of MNPs were confirmed by TEM (Fig. 2A). Perinuclear particles were never observed fusing with the nuclear membrane, localized within membrane bound structures or in obvious association with the endoplasmic reticulum or Golgi apparatus.
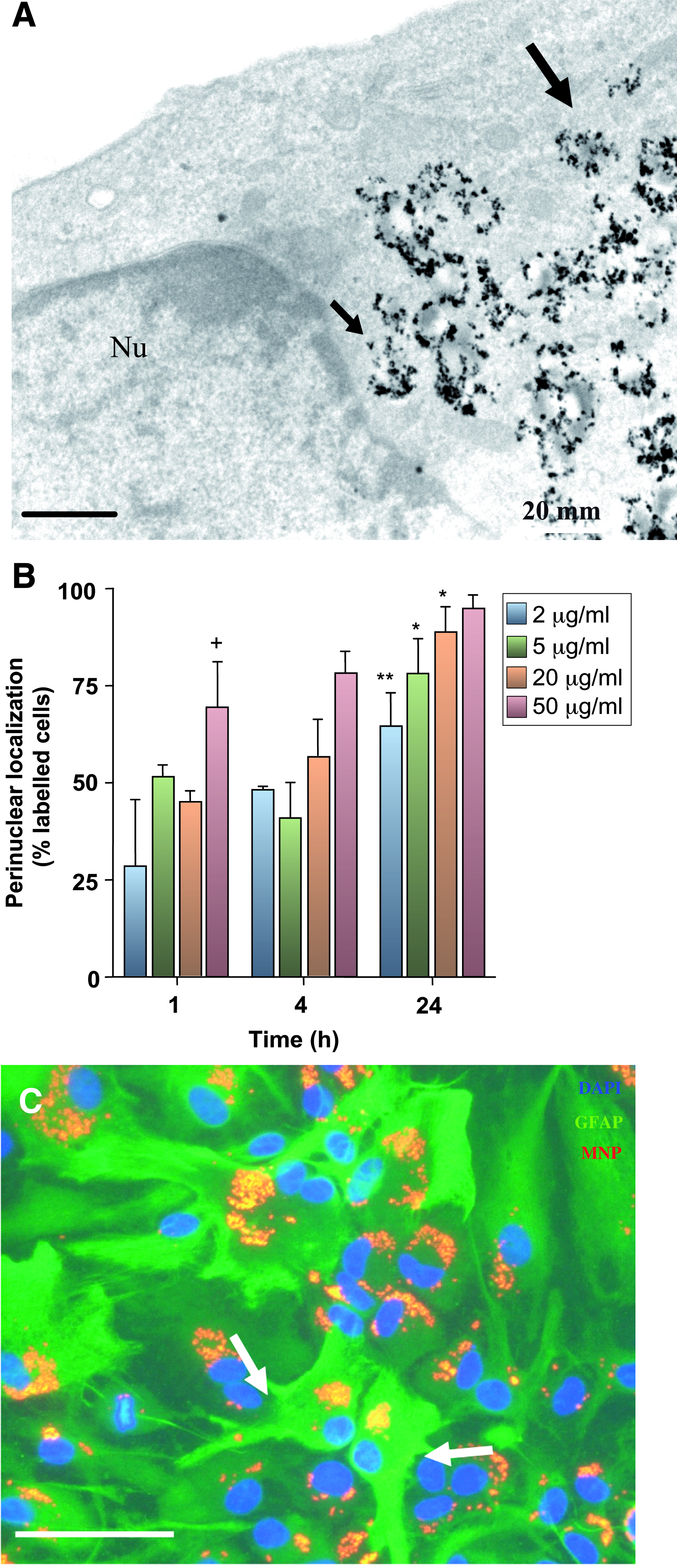
Perinuclear localization of MNPs in astrocytes.
Analysis of the subcellular location of MNPs revealed that in incubations with 2 μg/mL MNPs for 1 h, ingested particles were localized to the cytoplasm in most cells with a minor proportion showing perinuclear/nuclear distribution. The proportion of labeled cells containing particles with a perinuclear localization showed both time and concentration dependence (Fig. 2B). Astrocytes prelabeled with MNPs for 24 h, then cultured for a further 6 days retained large collections of particles (Fig. 2C). There was clear evidence of cellular heterogeneity of astrocytes with respect to the subcellular localization of MNPs at this time. Thus, although the majority (∼85%) of labeled cells contained particles in close proximity to the nucleus; the remainder contained MNPs with a predominantly cytoplasmic localization (Fig. 2C). Particles showed progressive dilution of label with time, with ∼82% of cells labeled at 14 days and 63% at 21 days.
Mechanism of MNP uptake and intracellular trafficking
MNP uptake was temperature dependant, as reducing the incubation temperature to 4°C markedly inhibited (by 84%) the proportion of cells showing uptake (Fig. 3A), indicating the energy dependence of uptake mechanisms. In endocytosis blocker studies, the proportions of astrocytes that accumulated MNPs were similar to that under both control conditions (Fig. 3B). Macropinocytosis inhibitors markedly inhibited MNP uptake by astrocytes (Fig. 3B) compared with DMSO alone (reduced by 66% [amiloride] and 70% [EIPA]). Inhibitors of clathrin-dependent endocytosis reduced the proportion of astrocytes displaying particle uptake to a lesser extent than macropinocytosis blockers (Fig. 3B); (reduced by 31% [tyrphostin 23] and 28% [dynasore]). A blocker of caveolin-mediated uptake (filipin) did not inhibit particle uptake (data not shown). In experiments where both inhibitors of clathrin mediated uptake and macropinocytosis were applied to cultures, significant cellular toxicity was observed; therefore, it was difficult to draw conclusions from these experiments. Cell densities, as judged by counts of DAPI-stained nuclei per field, were similar for astrocytes treated with and without inhibitors (Table 1), indicating the absence of associated short-term toxic effects of inhibitors alone.
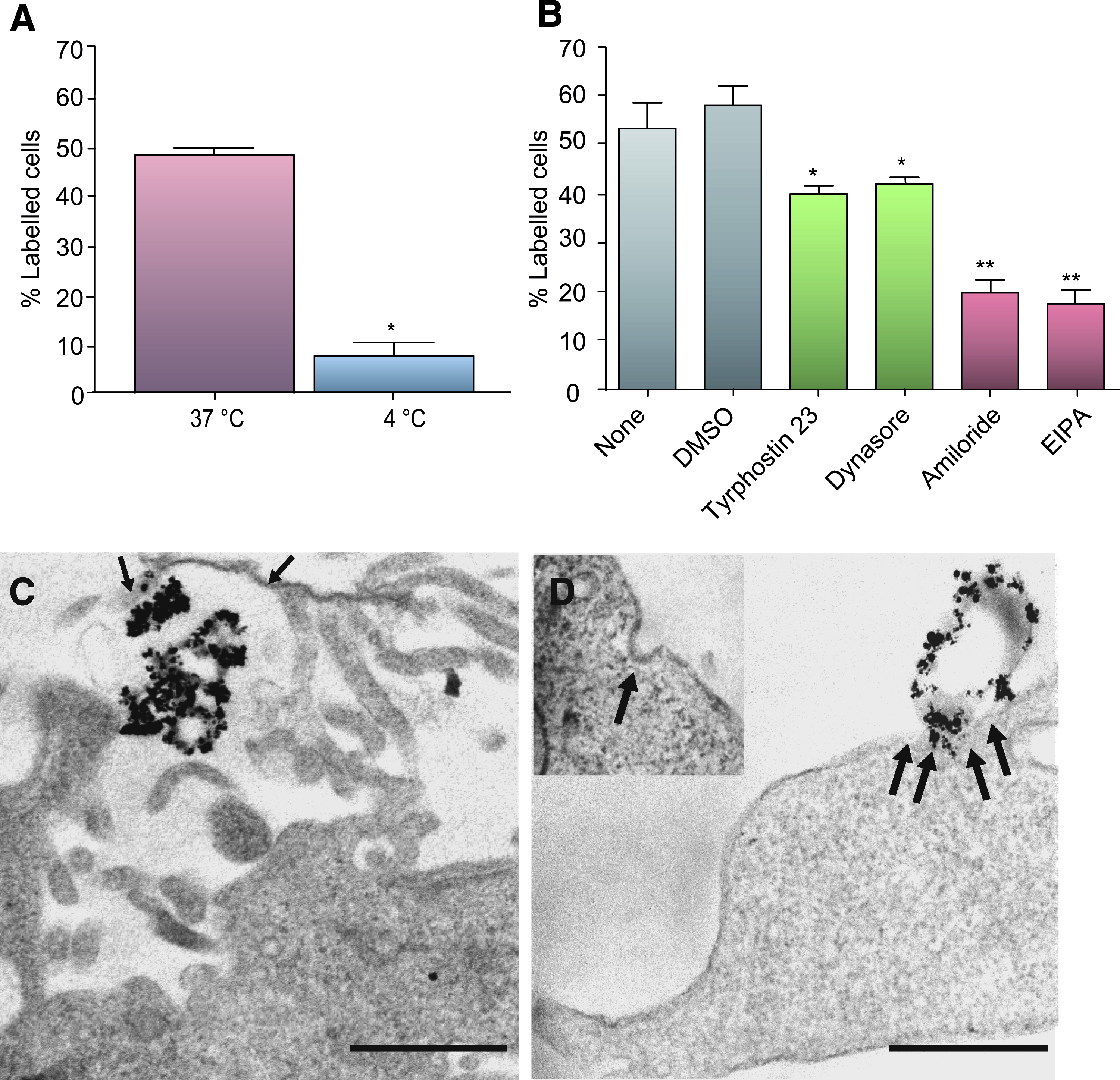
Mechanism of MNP uptake in rat primary astrocytes.
Data are from three different cultures.
DAPI, 4,6-diamidino-2-phenylindole; DMSO, dimethyl sulfoxide; EIPA, 5-(N-ethyl-N-isopropyl) amiloride.
TEM analyses supported findings from the inhibitor studies. Cells displayed extensive projections of the plasma membrane that appeared to be engulfing collections of MNPs (Fig. 3C), consistent with a role for macropinocytosis in particle uptake. Further, coated pit-like structures in the astrocytic plasma membrane were also observed (Fig. 3D; inset) and, occasionally, particles were seen externally, in close apposition to such structures (Fig. 3D), consistent with a role for clathrin-mediated endocytosis.
Toxicity
Incubation of astrocytes with 2–50 μg/mL MNPs for 24 h had no statistically significant cytotoxic effects as judged by a MTS assay, immediately after incubation with particles or after a further 24–96 h in medium alone (Table 2). Indeed, in the experiment to determine the time course and concentration dependence of MNP uptake, visual examination revealed that incubation of astrocytes with increasing concentrations of MNPs had no effect on counts of DAPI-stained nuclei per microscopic field, nuclear morphology (Supplementary Fig. S2, available online at
Data are from five different cultures.
Transplantation of labeled astrocytes into slice cultures
To assess if MNP-labeled astrocytes could survive and integrate within host tissue after transplantation, experiments were conducted using organotypic cerebellar slice cultures as the recipient tissue. Immediately after delivery of trypsinized cells, colocalization of DAPI and Nile Red (MNP) fluorescence could be observed (Fig. 4) in a highly localized focus within the slice (Fig. 4A; inset); cells typically display a rounded morphology at this stage with withdrawal of cellular processes, consequent upon trypsinization. At 10 days post-transplantation, a restricted area of DAPI fluorescence was detected within the slice (Fig. 4A); robust GFAP labeling was present throughout the slice that can be attributed to both host and transplanted astrocytes (Fig. 4B). A focus of Nile Red fluorescence associated with the injection site could also still be clearly observed (Fig. 4 C) with colocalization of DAPI and red fluorescence detected against a background of host tissue (Fig. 4D). Importantly, Nile Red and DAPI-labeled cells appeared to be radiating outward from the central application point, suggesting cell migration; the majority of cells appeared to remain at the application site. Further, high-power images of slices revealed the presence of multiple, well-differentiated DAPI/Nile Red/GFAP triple-positive cells (Fig. 4E), indicating that MNP label was retained by astrocytes and did not impair the ability of astrocytes to re-acquire their normal cellular morphologies after transplantation. When labeled astrocyte transplant populations were replated in culture wells, retention of particles was apparent (Fig. 4F), although a slight decline in the proportion of labeled cells with time observed (from ∼93% at 72 h to 70% at 10 days; this suggests that the majority of fluorescent signal detected in transplanted slices at 10 days is due to the transplant population). This was associated with a gradual decrease in the proportion of labeled cells showing MNPs in a perinuclear localization (from ∼80% at 72 h to 50% at 10 days), with particles showing a tendency to progressively accumulate near the cell membrane. Importantly, labeled cells were able to reacquire their normal morphological characteristics after trypsinization and transplantation (Fig. 4F), and cultures exhibited similar confluence and cell morphology to cultures that had been treated in a similar manner but incubated without either MNPs or DAPI (data not shown). The dominant morphology observed in both transplanted and plated astrocytes was that of type 1 cells (similar to the distributions observed before cell labeling), indicating that MNP labeling did not alter the phenotypic distributions of astrocytes.
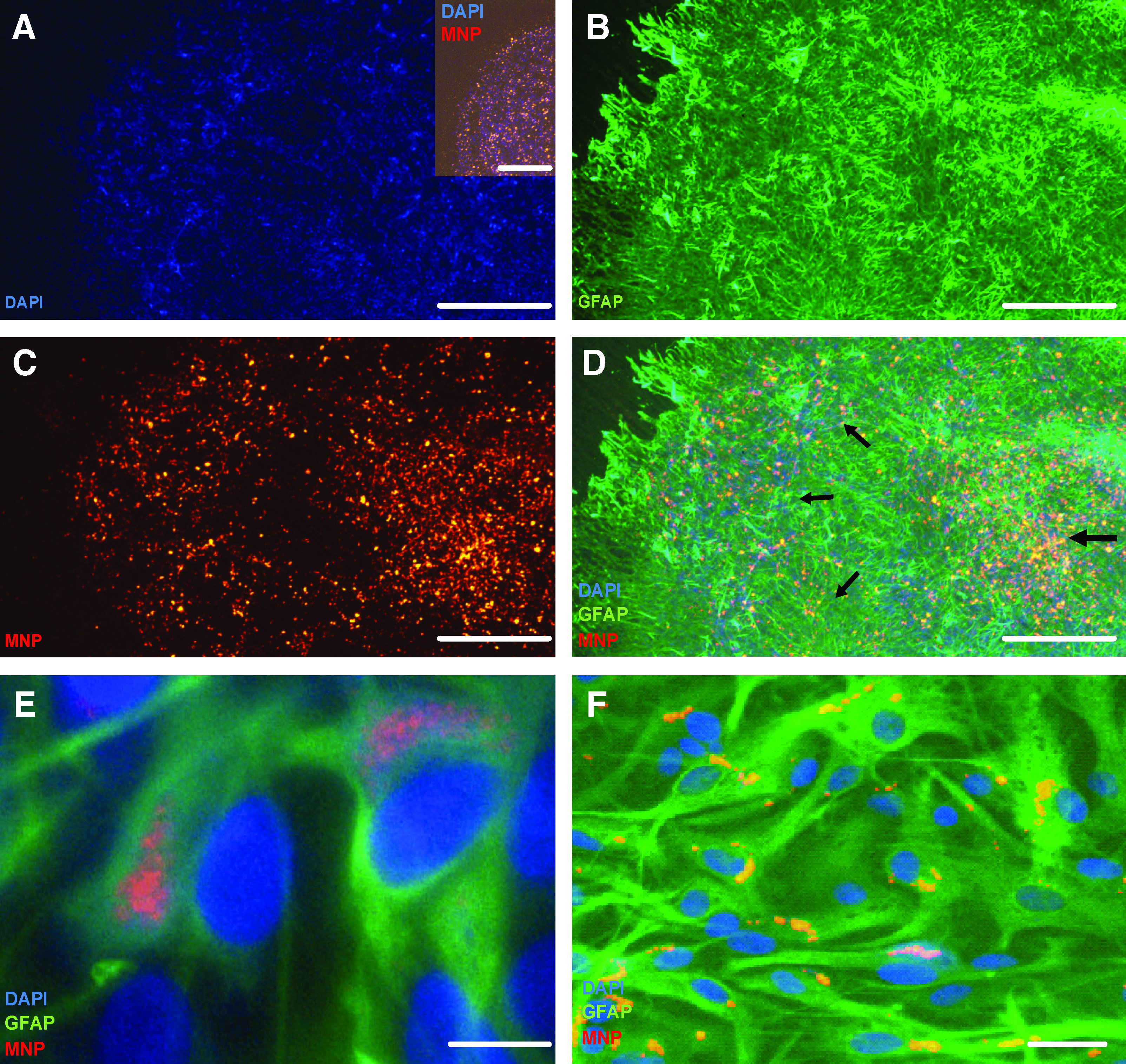
Survival and differentiation of astrocytes prelabeled with MNPs and 4,6-diamidino-2-phenylindole (DAPI) after transplantation into organotypic slice cultures of cerebellum.
Discussion
Here, we provide proof of principle (using cells obtained from primary cultures) that astrocytes derived for neural tissue engineering applications can be rapidly and efficiently labeled with MNPs. We employed relatively large (0.20–0.39 μm) particles for cell labeling in these studies; we have considered elsewhere the advantages of using particles with these dimensions. 27 We consider that the combination of methods used in this study is valid for several reasons. Experiments using cell lines have provided valuable information on the cellular dynamics of nanoparticle uptake; however, the use of untransformed cells can overcome a range of disadvantages that are inherent to the use of cell lines (such as microbial contamination, aneuploidy, the need for cellular karyotyping to establish cell identity, and altered cell physiology secondary to transformation). In cells that had been pulse-labeled with MNPs, close overlap was found between particle-associated fluorescence and iron-containing material, excluding possible experimental artifacts due to chemical instability of the particles or their intracellular degradation; this stability was further confirmed in electron microscopy analyses and may account for the limited particle toxicity observed for astrocytes. Confocal microscopy confirmed that cells had internalized particles rather than the latter adhering to the astrocyte surface. This observation, in conjunction with the marked reduction in MNP uptake at 4°C or in the presence of endocytosis inhibitors, suggests that the contribution of particles adherent to the astrocyte surface (to cell counts) was negligible and that cellular labeling is a true reflection of intracellular particle localization, which will be important for imaging applications involving transplanted astrocytes.
The extent and time course of particle uptake was found to be dependant on both extracellular MNP concentrations and the length of exposure to particles. These observations suggest that MNP loading of astrocyte transplant populations can be varied to suit the required downstream application by simply manipulating particle concentrations at the initial incubation step. Combined with the ease of detection of the particles using standard fluorescence techniques, these observations make the particles employed a desirable choice for studies related to cell tracking and intracellular MNP processing in astrocytes. Notably, we found that after embedding in gels, the particles can be readily observed using a 1.5 T MRI scanner that is used routinely in human clinical testing. Particle detection in vitro does not necessarily indicate that cells containing MNPs can be detected in vivo (as the sample itself will impact on image signal to noise ratio and contrast to noise ratio particularly in neural injury sites). However, our preliminary findings do suggest that MNP labeling of transplant populations may have considerable translational potential for clinical tracking of transplanted astrocytes in vivo, an issue that that warrants further investigation.
In pulse-labeling experiments, particles were retained in high numbers in astrocytes for up to 21 days. Indeed, recent preliminary data from our laboratory indicate significant particle retention for up to 28 days (the latest time point examined so far) although we observed a gradual dilution of MNP label with time. We can predict that such dilution may be due to cell division and/or cellular excretion (likely via exocytosis); the gradual reduction in perinuclear MNPs with time, in conjunction with progressive accumulation of particles near the cell membrane, would be consistent with a process of cellular excretion. During this period, cultures became confluent and exhibited normal morphologies, suggesting limited MNP effects on cell survival and differentiation—observations that were further supported by the MTS assays. Others have reported significant short-term toxicity of MNPs (of undisclosed formulation and size) on primary rat astrocytes, as evidenced by markedly reduced cell adherence, when nanoparticles were added to cells before attachment to the substratum. 28 We noted no effects on astrocyte adherence, possibly because cells were allowed to attach and mature before MNP addition. Our findings agree with a recent report that MNP addition (10 nm diameter particles) to astrocytes results in no acute loss of cell viability. 29
Importantly, astrocytes prelabeled with MNPs could survive and differentiate after transplantation into organotypic slice cultures of the rat cerebellum; the latter model provides a three-dimensional network of neural cells and has been widely employed in studies on neural regeneration, providing an ideal bridge between experiments on isolated cells and the intact CNS in vivo, while allowing for high-throughput assays. Although we cannot rule out that excreted particles and/or particles that are released from dying transplanted astrocytes are taken up by cells such as microglia and host astrocytes, the majority of the astrocyte transplant population (∼70%) was noted to be both MNP and DAPI positive at 10 days after plating on tissue culture plastic, suggesting that the majority of signal in host tissue is due to the transplant population. Our findings therefore indicate that this major neural cell population is highly amenable to labeling with MNPs and can provide the basis for the development of a technically simple and efficient method to safely label astrocytes derived for transplantation (which can be further adapted for long-term tracking of transplanted cells in host tissue). This method would be adaptable for either noninvasive imaging of transplant populations by MRI or for simple histological staining of tissue (to detect iron in transplant populations), and fluorescence/electron microscopy, making this a highly versatile labeling approach.
Findings from various mammalian cell types indicate that the intracellular accumulation of iron oxide-particles is mediated by endocytosis.30,31 With regard to the specific endocytotic pathways that mediate MNP uptake, there is limited information in relation to primary neural cells and no information with regard to primary astrocytes. In our experiments, the dramatic decrease in particle uptake by astrocytes after incubation at 4°C strongly suggests that the process is mediated by active transport; MNP uptake by primary astrocytes has been recently reported to exhibit a similar temperature dependence, although the actual uptake mechanisms were not addressed further. 29 Our endocytosis blocker analyses reveal that MNP uptake is mediated by two primary mechanisms: macropinocytosis (∼70%) and clathrin mediated uptake (∼30%); these findings are supported by our TEM analyses. Caveolin-dependent endocytosis, by contrast, does not appear to contribute to MNP uptake. In support of our findings, macropinocytosis is suggested to play a major role in the uptake of a range of nanoparticles (of a similar size to the MNPs employed in our study) in various cell types.32–35
We consider our observations on the mechanisms of MNP uptake to be significant from the point of view of developing high efficiency vectors to transfect astrocytes. The rapid perinuclear targeting of MNPs in astrocytes can clearly provide the basis for rapid delivery of genetic material to the nucleus for transfection applications (rapid trafficking of nanoparticles resulting in perinuclear accumulation has been reported previously for other cell types)36–38 but not, as far as we are aware, for astrocytes. However, a further point to consider with regard to transfection is the mode of particle uptake by astrocytes. Some studies argue that particle uptake via clathrin-dependent mechanisms is not advantageous for transfection, as endolyososomal trafficking is a predictable consequence of clathrin-mediated endocytosis representing a critical rate-limiting step. 39 By contrast, cellular uptake mediated by macropinocytosis may confer significant advantages for drug/gene delivery, since macropinosomes fuse with lysosomes at lower rates than clathrin-coated vesicles, 39 thereby minimizing lysosomal degradation of nanoparticles and/or their cargo (Fig. 5).32–34 As macropinocytosis accounts for 70% of MNP uptake in astrocytes, we can predict that without further manipulation, it will only be possible to transfect an upper limit of ∼70% of astrocytes. This is strongly supported by our recent report that particles (of similar dimensions to those used in this study and with perinuclear targeting properties) can be used to transfect between 43% and 75% of rat astrocytes.

Schematic diagram illustrating the primary modes of MNP uptake by astrocytes, subsequent intracellular processing and implications for gene delivery. Macropinocytosis, the major mechanism of astrocytic MNP uptake, can be considered a protransfection mechanism of uptake, as macropinosomes fuse with lysosomes at a low rate. Consequently, successful gene delivery to the nucleus is likely by this route. In contrast, clathrin-mediated endocytosis, a minor mechanism of astrocytic MNP uptake, can be considered a prodegradative mechanism of uptake, since clathrin-coated vesicles fuse with lysosomes at a high rate, resulting in degradation of cargo and particles. Color images available online at
Our findings that MNPs can be used effectively for labeling/imaging of astrocytes derived for transplantation (coupled with earlier findings that MNPs can be used for astrocyte transfection applications) lend support to the idea that MNPs are a versatile platform that can be employed for cell tracking and magnetic cell targeting, transfection, and noninvasive imaging applications for neural cell transplantation therapies. To develop the clinical translational potential of this approach, future challenges will involve collaboration between materials scientists and transplantation neurobiologists to develop multimodal iron oxide nanoparticles that can safely mediate the above combinatorial functions in neural cell transplant populations.
Footnotes
Acknowledgments
This work is supported by research grants from the British Biotechnology and Biological Sciences Research Council (New Investigator Award) and the Royal Society, UK. We would like to thank Ms. Karen Walker for expert assistance with TEM.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.