
Abstract
One of the factors that can impact human embryonic stem cell expansion in stirred microcarrier culture reactors is mechanical stress caused by agitation. Therefore, we have investigated the effects of agitation on human embryonic stem cell growth and expression of pluripotent markers. Agitation of HES-2 cell line in microcarrier cultures in stirred spinner and agitated six-well plates did not affect expression of pluripotent markers, cell viability, and cell doubling times even after seven passages. However, HES-3 cell line was found to be shear sensitive, showing downregulation of three pluripotent markers Oct-4, mAb 84, and Tra-1-60, and lower cell densities in agitated as compared with static cultures, even after one passage. Cell viability was unaffected. The HES-3-agitated cultures showed increased expression of genes and proteins of the three germ layers. We were unable to prevent loss of pluripotent markers or restore doubling times in agitated HES-3 microcarrier cultures by addition of five different known cell protective polymers. In addition, the human induced pluripotent cell line IMR90 was also shown to differentiate in agitated conditions. These results indicate that the effect of agitation on cell growth and differentiation is cell line specific. We assume that the changes in the growth and differentiation of the agitation-sensitive (HES-3) cell line do not result from the effect of shear stress directly on cell viability, but rather by signaling effects that influence the cells to differentiate resulting in slower growth.
Introduction
The development of large-scale, reproducible, and controlled cell culture methods is required for expansion and differentiation of human ESCs (hESCs) to meet the projected demand. One such method is the agitated three-dimensional microcarrier culture, which offers large surface area per unit volume for cell propagation leading to high cell density. Moreover, the well mixed nature of the agitated culture system allows for cell culture sampling, efficient mass transfer, and the ability to monitor and control the culture conditions, resulting in higher cell concentrations and consistency between batches. 7 To date, mouse ESCs (mESCs) have been successfully cultured on microcarriers in stirred bioreactor systems.8,9
Recently, our group 10 and others11,12 have reported on the propagation of hESCs on a variety of microcarriers. A multitude of passaging methods and carrier types were experimented on, with varying success ranging from 14- to 25-fold expansion over a week. Our group has shown that stable propagation of two hESC cell lines, HES-2 and HES-3, is possible on matrigel-coated cellulose-based carriers for >25 passages without loss of pluripotency.
Shear stress and hydrodynamic forces associated with agitation are known to bring about lowered cell viability and cell growth for many types of animal cells,13,14 and especially in microcarrier cultures due to further stress arising from turbulent eddies and particle-particle impact. 15 Stress effects have also been found to cause nonlethal effects, including changes in cell morphology. 16
To overcome the shear stress effects in animal cell cultures, protective polymers have been routinely added to the culture medium. These polymers aid in minimizing the damage by directly increasing viscosity of the medium, or changing the surface tension properties to minimize cell damage from stirring, bubble-burst events and foaming. 17
Effects of mechanical stress on stem cells have been found to be diverse, ranging from inhibition of hESC differentiation with biaxial cyclic strain,18,19 to promotion of differentiation of Flk-1-positive stem cells into the endothelial cell lineage with shear stress. 20 However, in these studies, the stress was applied by either stretching of a membrane seeded with cells, or varying flow rate in an enclosed chamber. While these methods allow for better control over magnitude of stress applied, it does not mimic the conditions in an actual stirred bioreactor.
In pluripotent stem cell microcarrier cultures, agitation is essential for suspending the cell covered microcarriers and achieving homogeneity of the system. Thus, in this article, we have investigated the effects of shear stress generated by agitation on cell growth and pluripotency of hESC and human induced pluripotent cell (hiPSC).
Materials and Methods
Two-dimensional human pluripotent stem cell culture
Two hESC line, HES-2 (46, XY) and HES-3 (46, XX), were obtained from ES Cell International. The cells were routinely cultured at 37°C/5% CO2 on matrigel-coated organ culture dishes. They were passaged by enzymatic treatment and fed daily with the conditioned medium from immortalized mouse embryonic feeders supplemented with 10 ng/mL of basic fibroblastic growth factor (Invitrogen) as previously described. 21 hiPSC, iPS (IMR90) was obtained from Jaime Thomson's lab. 22 Cells were cultured in the mTeSR1 (Stem Cell Technologies) serum-free medium on matrigel.
Three-dimensional human pluripotent stem cell microcarrier culture
hESC microcarrier culture was previously described by Oh et al. 10 To summarize, the cylindrical microcarriers used in this study are DE53, preswollen microgranular cellulose anion exchange (Whatman). These are positively charged (diethylaminoethyl) carriers having small ion exchange capacity of 2 milli-equivalents per gram dry material and dimensions of 40–50 μm in diameter, 80–400 μm in length. Before use, the carriers are washed with calcium- and magnesium-free phosphate-buffered saline (PBS), and sterilized by autoclaving. Thereafter, they are coated with matrigel (Becton Dickinson) at 4°C overnight. The matrigel-coated DE53 microcarriers (4 mg/mL) were seeded with either hESC or hiPSC at 0.8–6.0 × 105 cells/mL in ultra-low attachment six-well plates (Corning) containing 5 mL of the mouse embryonic feeders conditioned medium supplemented with 10 ng/mL basic fibroblastic growth factor. Eighty percent of the culture medium was refreshed every 24 h. The cultures were passaged every 7 days via mechanical dissociation.
Preparation of polymer additives
Polymers that are known to have a shear protective effect on cell culture were used in this study (Table 1). Stock solutions of polyethylene glycol (Fluka), methylcellulose (Sigma-Aldrich), and hyaluronic acid (HyaCare®) were prepared by dissolving in water and sterilized by filtration through 0.22 μm filters. Dextran (Sigma-Aldrich) was dissolved in water and autoclaved. Pluronic® F68 (Sigma-Aldrich) was available as a sterile solution for cell culture. Polymers were added to the conditioned medium at the concentration given in Table 1.
Human pluripotent stem cell agitation and spinner cultures
For agitation cultures, the six-well plates were placed on an orbital shaker at 100–120 rpm during the whole growth period. Six-well plates were chosen for the experiments so that multiple experiments could be conducted with different polymer conditions and with sufficient replicates for statistical comparisons, whereas spinner flasks simulate the agitated conditions in a bioreactor with stirrer blades, which would be realistically encountered in larger scale up volumes. The agitation conditions chosen were the minimum required for continuous suspension of the pluripotent stem cells on microcarriers.
Spinner cultures of hESCs were conducted in 100 mL magnetic spinner flasks (Scientific Industries; Bell-Flo™) precoated with Sigmacote® (Sigma-Aldrich). Cells, from static microcarrier cultures, were seeded into spinner flasks containing 8 mg/mL of new microcarriers and 25 mL of the culture medium achieving an initial cell density of 3.2–6.0 × 105 cells/mL. The culture was agitated at a speed of 20 rpm for 5 min at hourly intervals for the first 6 h, and left in static conditions overnight to allow for cell attachment. Thereafter, 25 mL of the culture medium was added and the culture was stirred continuously at 25 rpm and 80% of the growth medium was replaced daily.
Cell count
The total cell count was measured by the nuclei count method using NucleoCounter (ChemoMetec). Nonattaching/nonviable cell concentration was measured as per manufacturer's instructions, and cell viability was calculated.
Flow cytometry
Expression levels of pluripotent stem cell markers by the hESC and hiPSC populations were assessed by immunofluorescence using flow cytometry. The markers used were intracellular transcription factor Oct-4, and surface antigens recognized by mAb 84 and Tra-1-60, following previously published protocols.10,21,23
Single-cell suspensions of the hESC and hiPSC cultures were obtained by treatment with TryplE Express (Invitrogen) and filtration through a 40 μm sieve (Becton Dickinson) to remove the microcarriers from the cell suspension. hESCs or hiPSCs were then fixed, permeabilized (Caltag Laboratories), and incubated with mouse primary antibodies Oct-4 (Santa Cruz) at a 1:20 dilution (10 μg/mL), mAb 84 (BTI 23 ) at a 1:20 dilution (50 μg/mL), and Tra-1-60 (Chemicon) at a 1:50 dilution (20 μg/mL). Isotype controls were used for staining with all the antibodies. Cells were subsequently washed with 1% bovine serum albumin/PBS, and incubated in the dark with a 1:500 dilution (4 μg/mL) of goat α-mouse fluorescein isothiocyanate-conjugated antibody (Dako). The cells were then washed again and resuspended in 1% bovine serum albumin/PBS for analysis on a FACScan (Becton Dickinson FACS Calibur). All incubations were performed at room temperature for 15 min.
Quantitative real-time polymerase chain reaction
Single-cell suspensions free of microcarriers were obtained as described above. RNA was isolated using the NucleoSpin® RNA II kit (Macherey-Nagel) and quantified by ultraviolet spectrophotometry. cDNA was obtained using M-MLV reverse transcriptase (Promega) with 2 μg of RNA as template. cDNA was mixed with 200 nM of the forward and reverse primers shown in Supplemental Table S1 (available online at
Western blotting
Cells were lysed in RIPA lysis buffer (Cell Signaling Technology) and clarified by centrifugation. Twenty micrograms of protein was resolved on NuPage 4%–12% gradient gels (Invitrogen) and transferred onto polyvinylidene fluoride membranes (Millipore) at 110 V for 75 min. Membranes were blocked using 5% low-fat milk and incubated overnight at 4°C with the following primary antibodies: mouse anti-β-Actin (Santa Cruz), mouse anti-Oct-4 (Santa Cruz), mouse anti-AFP (A8452; Sigma), mouse anti-SMA (A2547; Sigma), and mouse anti-PAX6 (Developmental Studies Hybridoma Bank [DSHB]), followed by incubation with the appropriate secondary horseradish peroxidase-conjugated secondary antibodies (Dako). Signals were observed using the Immobilon Western Chemiluminescent HRP Substrate (Millipore).
Statistical analysis
All experiments were conducted at least in duplicates. Duplicate samples were obtained for calculations of growth kinetics, that is, average and min/max cell numbers. For experiments that were conducted in triplicates or more, statistical analysis was conducted. For these experiments, statistical significance was evaluated using unpaired Student's t test, whereby p < 0.05 is considered to be statistically significant.
Results
Effects of agitation on growth and pluripotency of hESC and hiPSC
HES-2 and HES-3 cell lines were propagated on cellulose microcarrier cultures under shear stress generated either by agitation of six-well plates in orbital shakers (agitated), or stirring in spinner flasks (spinner). Control cultures (not exposed to shear stress) were propagated under static conditions in six-well plates (static) as a reference.
Results presented in Table 2 show that HES-2 remains largely unaffected by the shear stress. Doubling time for both static microcarrier cultures and spinner cultures are similar at 49.5 and 54.8 h, respectively. Cells remain pluripotent in all three cases, as indicated by high Tra-1-60 expression of >90%, even for long-term agitation culture in orbital shakers (at least seven passages). Higher final densities were reached in both agitation and spinner cultures (23.45 and 24.60 × 105 cells/mL, respectively) as compared to static cultures (16.96 × 105 cells/mL). This higher cell yield can be attributed to a better mass transfer in a more homogeneous culture system.
Doubling time measured during one passage. Average of two runs.
HES-2 was seeded at 2.0 × 105 cells/mL for static and agitation cultures, and 6.0 × 105 cells/mL for spinner cultures. Data for static microcarriers and agitation cultures are average of results from seven passages. Spinners were only followed for one passage.
HES-3 was seeded at 0.8 × 105 cells/mL for static, and 1.6 × 105 cells/ml for agitation and spinner microcarrier cultures. Data represent average of seven runs for static cultures, five for agitated culture, and two for spinner culture.
N.D., not determined.
On the other hand, shear stress was found to have a profound effect on HES-3 propagation in microcarrier cell cultures. As presented in Table 2, in the presence of shear stress, doubling time was increased to 49.7 and 39.2 h in agitation and spinner cultures, respectively, compared to the shorter doubling time of 31.2 h in static cultures, which is similar to HES-3 two-dimensional (2D) colony culture of about 30 h. 10 In addition, the final cell densities achieved by HES-3 in the presence of shear stress, 10.9 × 105 and 9.75 × 105 cells/mL for agitation and spinner cultures respectively, were lower than that of static cultures of 12.2 × 105 cells/mL.
Representative growth kinetics and morphological appearance of HES-3 cell culture in agitated microcarrier cultures are presented in Figure 1. HES-3 cells were seeded at a concentration of 1.6 × 105 cells/mL into static, agitated, and spinner microcarrier cultures. During the first stages of cell growth, all cultures behaved in a similar manner, with a long lag period of about 72 h, in which viability dropped to as low as 75%–87% on day 1. Thereafter, cells began to expand and reach the exponential growth phase from 72 to 144 h after seeding. At this stage, the difference in doubling times and final cell yield can be observed, with agitation cultures reaching only about 60% of the cell density (~7.0 × 105 cells/mL) of the static culture (~11.5 × 105 cells/mL) 144 h after seeding. After 144 h, cell growth in both static and spinner culture plateaued, whereas agitation cultures remained in the exponential phase. The final cell densities of the static, agitation, and spinner cultures were 12.7 × 105, 10.3 × 105, and 9.7 × 105 cells/mL, respectively, and doubling times were 31, 49, and 39 h, respectively (Table 2).

HES-3 cultured on microcarriers in the presence and absence of agitation.
We suspect that the relatively longer doubling times obtained in agitation and spinner cultures could be attributed to the partial differentiation of the HES-3 culture when subjected to shear forces. Indeed, expression of key pluripotency markers, Oct-4, mAb 84, and Tra-1-60, dropped within one passage in agitation (Table 2 and Fig. 1B), from above 90% Tra-1-60 expression in the seeding cultures, to 64.9% and 67% for the agitation and spinner cultures, respectively. Morphology-wise, both agitation and spinner hESC cultures gave rise to dense, spherical clumps of cells, showing cystic structures indicating the appearance of differentiating cell populations (Fig. 1C). On the other hand, in static microcarrier cultures, sheet-like structures without cystic regions were observed.
This loss of cell pluripotency and decrease in growth properties is intensified during continuous passaging (Fig. 2). HES-3 cells were transferred for three consecutive passages in static and agitated microcarrier cultures. While the static culture maintained a stable final cell density of 13.8 × 105 cells/mL and high expression of pluripotent markers like Tra-1-60 at 92.1% (Table 2), we observed in the agitated cultures a gradual decrease in cell yield (dropping down to 5.6 × 105 cells/mL after three passages), and in expression of pluripotent markers like Oct-4 and Tra-1-60 to below 40% (Fig. 2).
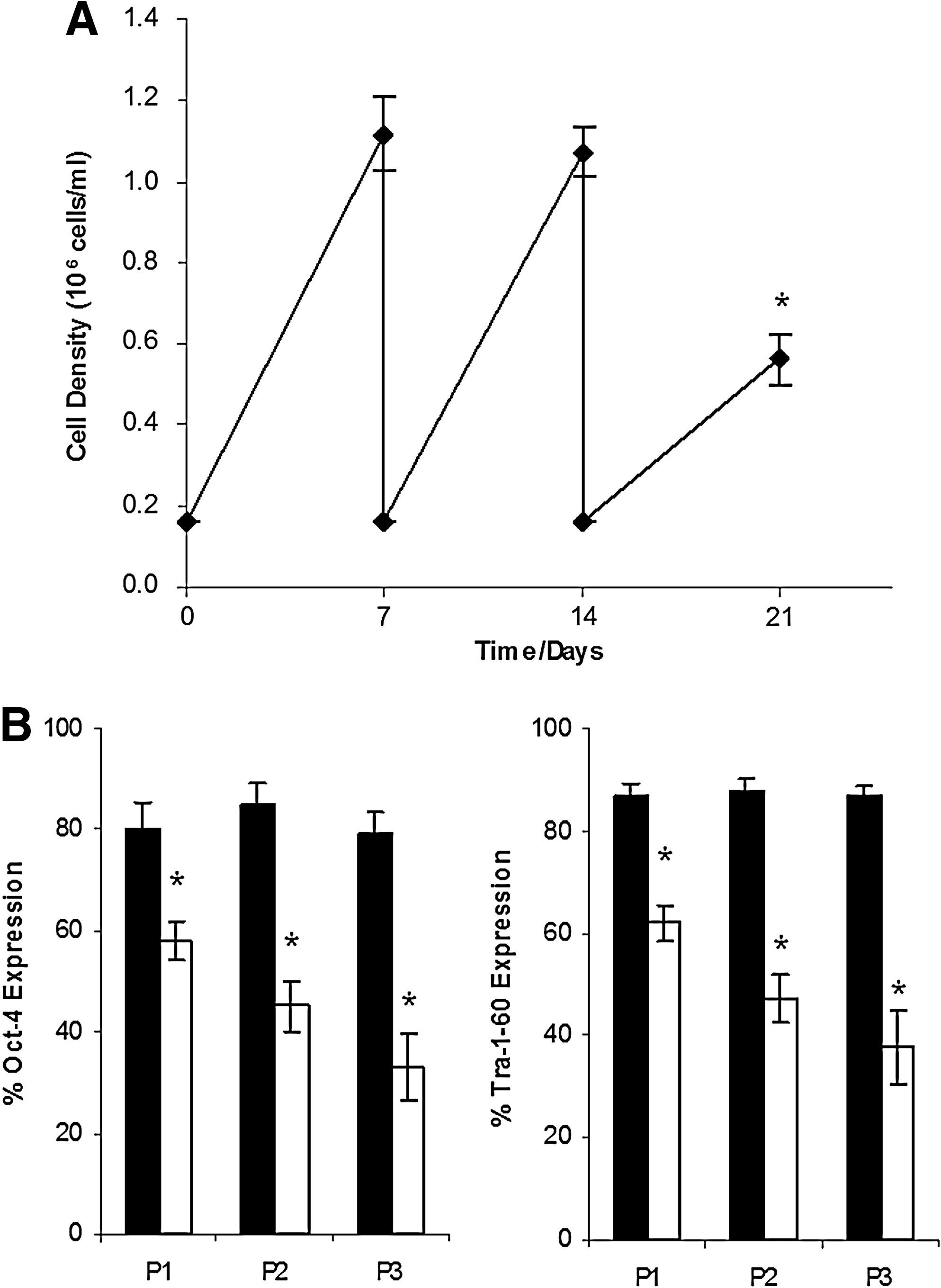
Long-term effects of continuous agitation on HES-3 cultured for three passages. Final cell density
To determine if agitation could also induce spontaneous differentiation of hiPSC, we subjected the IMR90 hiPSC line to 3 weeks of passaging with continuous agitation (100 rpm in six-well plates) in microcarrier cultures. This line also downregulated the pluripotent marker Tra-1-60 to 50% expression levels, and exhibited significantly reduced cell growth compared to the control static conditions (Supplemental Fig. S1, available online at
Differentiation of agitated HES-3 microcarrier cultures
The shear-induced differentiation process of HES-3 cell line was further investigated at the genetic and protein levels to find out if it is directed to spontaneous differentiation to all three germ lines similar to embryoid bodies (EBs) or toward a particular lineage. To this end, HES-3 cells were propagated for three consecutive passages in agitated and static (control) microcarrier cultures. Thereafter, cells were harvested and analyzed for RNA expression by quantitative real time (qRT)-polymerase chain reaction (Fig. 3A) and protein expression by western blotting (Fig. 3B). In the agitated culture, the pluripotent marker, Oct-4, dropped in expression in both the transcript and protein level by 33% and 80%, respectively, as compared with static culture. This was congruent with our previous observation of decreased pluripotency of HES-3 on microcarriers after prolonged culture in agitation (Fig. 2). At the same time, analysis of the qRT-polymerase chain reaction and western blot data showed upregulation of markers of all three germ layers, in the agitated culture, without bias toward any particular lineage. This was only a partially differentiated culture, and Oct-4 was not completely downregulated, especially at the transcriptome level, whereby mRNA levels were still 67% that of static microcarriers. However, Oct-4 protein levels were significantly reduced. Prolonging the agitation culture may allow better understanding of the differentiation induced by shear forces, though at the expense of cell yield.
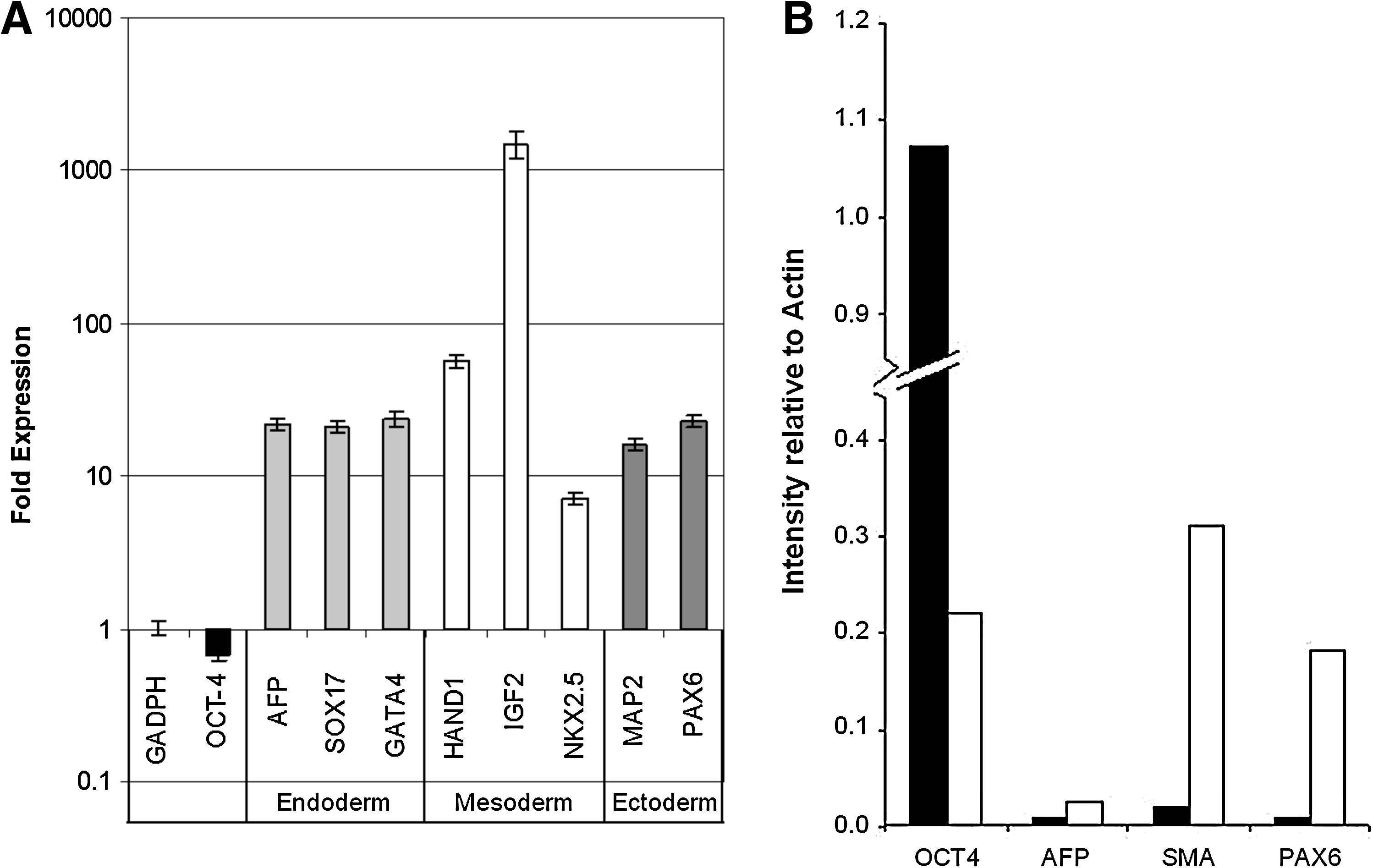
mRNA and protein levels in HES-3 partially differentiated agitated cultures (after three passages).
Prevention of shear stress by use of protective polymers
Various protective polymers are used to reduce the effects of shear stress in agitated animal cell cultures.7,13,15,24 The polymers are classified into three main categories: surface acting polymers (Pluronic F68 and polyethylene glycol), viscosity enhancing polymers (methylcellulose and dextran), and polymers with mixed effect (hyaluronic acid), which increases viscosity of the medium, and is a prominent component in extracellular matrices.
The polymers were used at concentrations indicated in Table 1, which were reported in the literature to have a protective effect on cells (see references in Table 1). Addition of the polymers to 2D colony cultures, or static microcarrier cultures, did not affect cell growth and pluripotency with the exception of Pluronic F68. When Pluronic F68 was added to the 2D colony and static microcarrier cultures, FACS data showed a 28% drop in expression of Oct-4, and 22% drop in Tra-1-60 expression within one passage. Prolonged culture with Pluronic F68 resulted in further loss in pluripotency, and spontaneous differentiation could be observed from the morphology of the 2D cultures (data not shown).
At the second stage, the polymers were added to HES-3-agitated microcarrier cultures, which were propagated for three consecutive passages. Final cell yield and expression of pluripotent markers in polymer supplemented as compared to nonsupplemented agitated and static cultures are presented in Figure 4. All the tested polymers did not protect the cells from decrease in cell growth (about 50.2% with respect to static microcarrier cultures) and loss of pluripotent markers (about 44% for Oct-4 and 43% for Tra-1-60 with respect to static cultures) generated by the shear stress under agitation conditions.
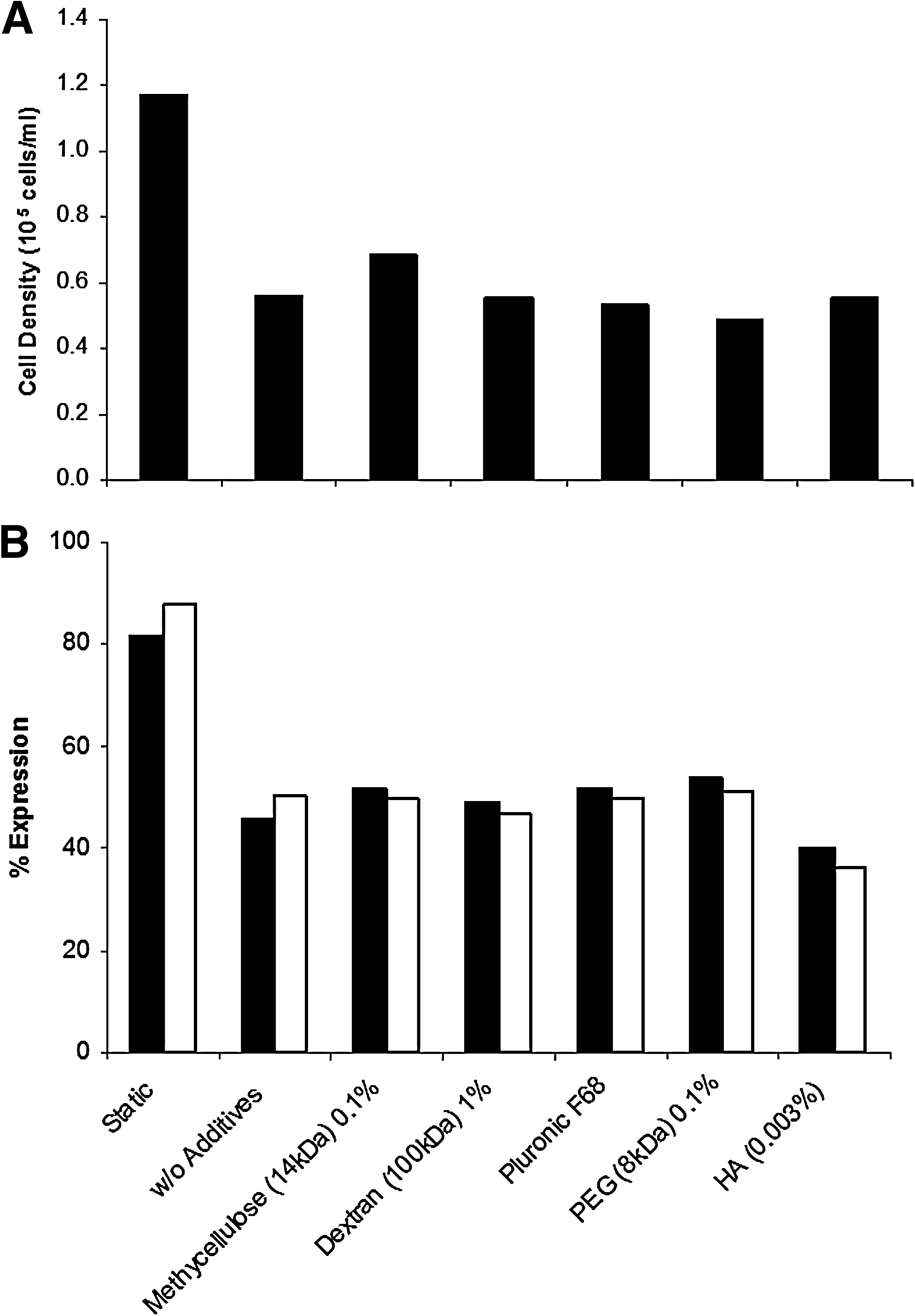
Effects of addition of protective polymers: methylcellulose, dextran, Pluronic F68, polyethylene glycol, and hyaluronic acid on HES-3-agitated microcarrier culture (after three passages at 120 rpm).
Discussion
mESCs,8,9,25 mESC-derived EBs, 26 hESC-derived EBs, 27 and human adult stem28,29 cells have been successfully cultured on microcarriers and suspension cultures in agitated bioreactors. However, the scale-up of human pluripotent stem cell culture on microcarriers in bioreactors10–12 is still a relatively novel area for research, and the knowledge about the effects of mechanical stress generated by agitation in bioreactors is currently very limited. In this study, the impact of mechanical stress on cell growth and differentiation in agitated hESC and hiPSC microcarrier cultures was investigated.
The effect of shear stress on mammalian cells in agitated bioreactors has been investigated by many researchers,13–15 addressing the issue of shear-induced reduction in cell viability and yield. Protective polymers, like methylcellulose and Pluronic F68, have been added to the culture medium to improve characteristics like increasing viscosity and/or changing surface tension properties to minimize cell damage resulting from agitation and sparging. Such polymers have been used successfully in many animal cell cultures to improve cell viabilities.24,30
In contrast to mammalian cells used in biologics production, different kinds of mechanical stresses are known to affect growth and differentiation of stem cells. Stem cells are known to have the mechanisms for sensing shear stress. 31 Extensive studies have been made especially on the impact of shear forces on endothelial and vascular tissue differentiation, in both mesenchymal stem cells (MSCs) and ESCs. It was found that various methods of exposure to mechanical stress and strain, including exposure to flow,20,32 or stretching on flexible membranes, 33 improved the efficiency of generating mature endothelial cells with improved angiogenic and vasculogenic potential as compared to without shear stress. Knippenberg et al. have also reported increased osteogenic potential in MSCs when exposed to fluid shear stress. 34 As can be seen, the impact of mechanical stress on stem cells is varied, depending on the type of strain, magnitude of strain, and the cell type used. 35 However, to the best of our knowledge, there are no data on the effect of agitation stress on hESC or hiPSC in a bioreactor-like condition.
The HES-2 cell line was found to be resistant to mechanical stress generated by agitation in plates and in spinner cultures. Pluripotent markers like Tra-1-60 remained above 90% even for long-term expansion of HES-2 under agitation at 100 rpm for at least seven passages (Table 2). Cell density was 50% higher than that of static microcarrier cultures in both agitation and spinner cultures (up to 2.5 million cells/mL), probably as a result of better mass transfer. This kind of shear resistant cell line can be a good candidate for undifferentiated hESC scale-up of microcarrier cell cultures in agitated bioreactors. These data complement research conducted by other groups, whereby other hESC lines were successfully cultured on microcarriers in spinner flasks and suspension cultures.11,12
On the other hand, the HES-3 and IMR90 cell lines were found to be very sensitive to shear. In spinner and agitated plate cultures, gradual loss of the pluripotent markers, Oct-4, mAb 84, and Tra-1-60, was observed throughout the 7 days of propagation, dropping from above 80% to below 60% within just one passage. The decline in pluripotent markers increased during serial passaging, dropping down to below 40% after three consecutive passages (Fig. 2 and Supplemental Fig. S1). The loss of pluripotency was accompanied by a parallel decline in cell growth, with agitated culture reaching less than half the maximal cell density (~6 × 105 cells/mL) of static microcarrier culture after three passages, probably as a result of the limited growth potential of the partially differentiated cells.
This loss of expression of cell pluripotent markers cannot be overcome by the use of protective polymers, which are known to protect animal cells from mechanical damage caused by agitation. This may suggest that the loss of cell pluripotency and decline in cell growth is not a result of mechanical effects generated on the cell membrane, but perhaps indirect signaling effects similar to those seen with endothelial and MSCs.20,34 The vastly different morphology exhibited by static microcarriers (multilayered sheet-like structures) as compared to that under agitation (tightly packed spherical clumps) may also hint at possible alterations in cell–cell interactions that may have resulted from constant agitation and disruption of cell–cell contact. Results from both gene and protein expression analysis show that the mechanical stress exerted on HES-3 cells in agitated cultures results in differentiation into all three germ lines without bias toward any particular lineage, similar to that of spontaneous differentiation induced by EB formation. These results show that it is problematic to propagate undifferentiated HES-3 cell line in conventional stirred reactors. Special low shear impellers, limited number of passages, low shear fluid-lift reactors, or static fixed-bed reactors could be explored as possible alternatives to expand HES-3.
Further investigations should be made regarding the differences in the transcriptome and proteome levels of HES-2 and HES-3 cultured under agitation conditions, given their vastly different response to shear stress. This would allow a better understanding of the mechanisms triggered under agitation conditions, and also the development of methods to screen for hESC and hiPSC cell lines more suitable to expansion in large scale stirred bioreactors.
Conclusions
The expansion of human pluripotent stem cells as suspended microcarrier cultures in bioreactors is one of the solutions to the growing demand of large amounts of hESC and hiPSC needed for research and regenerative medicine. We have investigated the effect of mechanical stress generated by the reactor agitation system on growth and differentiation of hESC and hiPSC. We have found that shear stress effects are cell line specific. In shear sensitive cell lines like HES-3 and IMR90, a rapid decline in pluripotency and growth yield is seen even during the first culture passage. Cells differentiated into all three germ lines without preference toward a particular lineage. Attempts to reduce the effects of agitation in culture with conventional protective polymers used in industry proved to be insufficient in terms of cell growth and maintenance of pluripotency. Thus, for large-scale expansion of human pluripotent stem cells in microcarrier culture in agitated bioreactors, there is a choice of either using a shear-resistant cell lines like HES-2, or shear-sensitive cell lines in low shear bioreactors (e.g., fixed-bed reactors or special low shear agitation systems).
Footnotes
Acknowledgments
We would like to thank Dr. Hia Hui Ching for critically reviewing this article and the Agency for Science Technology and Research (A*STAR) for generous funding of this research project.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.