
Abstract
Providing fully mature and functional osteoblasts is challenging for bone tissue engineering and regenerative medicine. Such cells could be obtained from multipotent bone marrow mesenchymal stem cells (MSCs) after induction by different osteogenic factors. However, there are some discrepancies in results, notably due to the use of sera and to the type of osteogenic factor. In this study, we compared the osteogenic differentiation of bone marrow MSCs induced by dexamethasone (Dex) or bone morphogenetic proteins (BMPs) by assessing phenotypes in vitro and functional osteoblasts in vivo. Reducing the content of fetal calf serum from 10% to 2% significantly increased the mineral deposition and expression of osteoblastic markers during osteogenesis. In comparison to Dex condition, the addition of BMP4 greatly improved the differentiation of MSCs into fully mature osteoblasts as seen by high expression of Osterix. These results were confirmed in different supportive matrixes, plastic flasks, or biphasic calcium phosphate biomaterials. In contrast to Dex-derived osteoblasts, BMP4-derived osteoblasts from MSCs were significantly able to produce new bone in subcutis of nude mice in accordance with in vitro results. In conclusion, we describe a convenient ex vivo method to produce consistently mature functional osteoblasts from human MSCs with use of BMP4 and low serum.
Introduction
MSCs are commonly selected by a culture step based on basal medium containing a specific serum screened for its capacity to maximize the expansion of multipotent cells.6,7 However, such sera contain numerous factors such as cytokines, growth factors, chemokines, hormones, substrate-attachment molecules, and binding proteins for transport and presentation. Although the utility of sera is crucial for the growth and survival of cells, their undefined content, as well as their batch-to-batch variability, could increase the difficulties in optimizing osteo-regenerative cell production for tissue engineering. In addition, sera can have inhibitory activities in osteogenic differentiation. Notably, sera contain molecules that can activate the extracellular receptor kinase pathway, thereby suppressing BMP-induced osteogenesis by decreasing Smad translocation. 8 Recently, Maeda et al. analyzed mechanisms allowing serum-derived TGF-β1 to inhibit the osteogenic maturation of the myofibroblastic cell line C2C12. 9 The authors also confirmed the results with some data about human MSCs but did not explore the full maturation and function of the generated human osteoblasts. Therefore, the use of BMPs as osteogenic factors combined with high serum content seems to be inefficient for inducing bone fate except with very high doses of BMPs (i.e., >300 ng/mL). In addition to sera, the glucocorticoid molecule dexamethasone (Dex) is widely used to induce MSC osteogenic differentiation in vitro and to study bone biology in vivo. However, Dex effects are controversial because the molecule increases mineralization in culture while decreasing the activity of bone cells in vivo.10–13 Such discrepancies could be explained by disturbances observed in nonosteoblastic cells in vivo known to have strong functions in bone formation, notably in osteoclastic cells. 14 For example, osteoclast functions are inhibited with Dex treatment. 15 As a result, consensus is lacking on efficient osteogenic factors, despite their crucial role in studying human bone biology and bone regenerative medicine.
Therefore, we aimed to investigate the benefits of reducing serum content in osteogenic media whatever the osteogenic inducer, Dex or BMPs, for MSC osteogenic differentiation. We confirmed the effects in vitro and in vivo and further showed that, compared to Dex, BMP4 was the strongest factor, even at a low concentration, for inducing full osteogenic differentiation of MSCs and ectopic bone formation in subcutis of nude mice, thus reducing the time needed to obtain the full maturation of osteoblasts.
Materials and Methods
Cell isolation and expansion
According to the Ethics Committee of Tours University Hospital and after receiving the written informed consent of normal donors (n = 17 donors; age = 59.8 ± 3.9; range = 37–80), 20 mL of bone marrow was harvested by iliac crest aspiration with a Jamshidi needle.
MSCs were obtained as previously described for clinical use.
16
Briefly, bone marrow nucleated cells (5 × 104 cm2) were cultured in proliferation medium of alpha-modified Eagle's medium (α-MEM; Invitrogen) supplemented with 2 mM L-glutamine, 100 U/mL penicillin, 100 U/mL streptomycin, 10% fetal calf serum (FCS; Hyclone), and 1 ng/mL fibroblast growth factor-2 (R&D Systems) and incubated in a humidified atmosphere with 5% CO2 at 37°C. The medium was renewed twice weekly until cells reached confluence. Then, cells were harvested, seeded at 1 × 103 cm2, and expanded in culture flasks until passage 2 before being seeded onto plastic dishes or ceramic discs (described in detail later) for osteogenic induction. Analysis of MSC characteristics was as described previously,
17
which confirmed the phenotype and multipotency of all expanded cells tested (Supplementary Fig. S1; Supplementary Data are available online at
Induction of osteoblastic differentiation
Different α-MEM-based osteogenic media were tested during the indicated times. The first contained β-glycerophosphate (βGP) (10 mM), ascorbic acid-2-phosphate (AAP) (0.2 mM), and Dex (10−7 M; all Sigma-Aldrich) supplemented with 10% FCS (v/v; Dex-10% FCS) or 2% FCS (Dex-2% FCS). The others consisted of α-MEM-based media with 50 ng/mL BMP2, 4, or 7 (R&D Systems), BGP + AAP supplemented with 10% or 2% FCS.
Histochemical analysis
MSCs were cultured at 1 × 104 cells/cm2 in chamber slides (Lab-tek/Nalge Nunc) in the evaluated differentiation media. To detect mineralization after induction, cells were treated with Alizarin red and Von Kossa reagents (both Sigma-Aldrich) for calcium detection and then observed under a photonic microscope (Leica Microsystems).
Western blot analysis
Culture of cells incubated with the different osteogenic media was stopped at different times (0, 7, 14, and 21 days). Western blot analysis was performed as described. 17 The membranes were washed three times and incubated with the primary antibodies anti-α-smooth muscle actin (αSM-actin, clone 1a4; Sigma-Aldrich), antitissue nonspecific alkaline phosphatase (ALPL, clone B4-78; R&D Systems), anticalcium-sensing receptor (CaSR; clone F-19; Santa Cruz Biotechnology), anti-osterix (polyclonal; GeneTex), and anti-β-actin (clone AC15; Sigma-Aldrich) in phosphate-buffered saline containing 5% bovine serum albumin overnight at 4°C. After a washing, membranes were incubated with a secondary antibody (Bio-Rad Laboratories) for 1 h at room temperature. Protein bands were observed with the ECLplus Western Blotting Detection Kit (GE Healthcare), and images were acquired with use of Chemi-smart 2000 and Chemocapt software (Vilbert-Lourmat).
Flow cytometry
Monoclonal antibodies used for phenotyping MSCs were anti-CD45, anti-CD14, anti-CD73, anti-CD90, anti-CD105, anti-CD146, and anti-CD166 conjugated to phycoerythrin (Becton-Dickinson). Cells were also incubated with phycoerythrin-conjugated anti-ALPL for flow cytometry. An irrelevant isotypic antibody was used as a negative control. To determine Smad phosphorylation after BMPs induction, we used rabbit anti-phosphorylated Smad 1, 5 antibody after fixation and permeabilization of cells according to the manufacturer's protocols (Cell Signaling). Cells were then analyzed by use of a FACSCalibur flow cytometer (Becton-Dickinson) with a 488-nm argon laser. Samples were analyzed by collecting 30,000 events by Cell-Quest software (Becton-Dickinson).
Culture on biphasic calcium phosphate scaffolds
Macroporous biphasic calcium phosphate (BCP) ceramic discs 8 mm in diameter and 3 mm thick (Biomatlante SA) were composed of 20 hydroxyapatite and 80 wt.% β-tricalcium phosphate and sterilized by gamma irradiation (total porosity: 70–75%; macroporosity: 400 μm). To minimize ion release, discs were incubated for 48 h in twice-refreshed proliferation medium (α-MEM, 10% FCS, and 1% antibiotic/antimycotic) before cell seeding. Cells were then seeded at 7.5 × 105 cells per BCP disc in nontreated culture dishes (Corning Life Sciences) and cultured during 1, 7, 14, and 21 days in the assessed media (i.e., proliferation medium, or Dex or BMP-containing medium supplemented with 2% or 10% FCS).
Reverse transcription polymerase chain reaction analysis
Real-time reverse transcription polymerase chain reaction (RT-PCR) was used to evaluate the relative level of genes characteristic of osteoblastic commitment. Cells cultured on ceramic discs or plastic dishes were transferred into a 1.5-mL tube with lysis buffer and vigorously shaken to lyse the cells. Total RNA was extracted with use of an RNA extraction kit (Qiagen) according to the manufacturer's instructions.
After desoxyribonuclease I digestion (Invitrogen), RNA samples (1 μg) were reverse transcribed with avian myeloblastosis virus-reverse transcriptase and random primers in a total volume of 25 μL. Real-time quantitative PCR was performed in the Chromo4 System (Bio-Rad Laboratories) with SYBR Green detection and Titanium Taq DNA polymerase according to the manufacturer's recommendations. PCR amplification involved 39 cycles of 30 s at 98°C, 15 s at 95°C, and 30 s at 60°C. Expression of the target gene was normalized to that of the endogenous control glyceraldehyde 3-phosphate dehydrogenase. The 2−ΔΔCt (cycle threshold) method was used to calculate relative expression levels as previously described. 18 Primer sequences are summarized in Table 1.
Animal study and histology
All animal procedures were performed in accordance with a protocol approved by the local committee for animal care and ethics. Notably, standards and practices in animal experiments are considered statistical significant for 5% or p-value <0.05. This significant value is usually obtained for n = 5 animals per group at minimum. Two hundred microliters of medium containing 7.5 × 105 human MSCs was seeded onto BCP discs and after their absorption 1 mL of medium was added (experiments repeated with MSCs deriving from three different donors). The cells were cultured in the different tested media for 7 days in low attachment plates, and media were refreshed twice weekly. Three different media were studied: classical osteogenic medium with 10% FCS (Dex-10% FCS), osteogenic medium with 2% FCS (Dex-2% FCS), and BMP4 medium with 2% FCS (BMP4-2% FCS). Five samples were used for each condition. After this culture period, the BCP discs were subcutaneously implanted in the backs of 10 nuclear magnetic resonance imaging (NMRI) nude mice (Elevage Roger Janvier). A 1-cm-long incision was made on each side of the mouse back, and blunt dissection was performed to separate the skin from subcutaneous connective tissue to form pockets into which the discs were inserted. The mice were killed after 8 weeks by lethal intracardiac injection of thiopental, and the implants were retrieved. The explants were processed according to standard operating procedures for nondecalcified histology. The samples were immersed in Karnovsky's fixative (4% formaldehyde and 5% glutaraldehyde), dehydrated in a graded series of ethanol, and embedded in polymethylmethacrylate. After polymerization, 100-μm-thick cross sections of the samples were obtained by use of a diamond-saw microtome (Leica SP 1600; Leica Microsystems), stained with methylene blue and basic fuchsin, and analyzed by light microscopy
Bone quantification
Newly formed bone within each explant was quantified by back-scattered scanning electron microscopy. Sections of the explants were polished and gold sputter-coated by use of a Cressington 108 auto apparatus (Cressington Scientific Instruments). Before histomorphometrical analysis, back-scattered scanning electron microscopic images were taken at 15 kV by use of an electronic microscope (Leo 1450 VP Carl Zeiss). This technique allows for separating bone from the implanted ceramics on the basis of their respective electronic densities. The amount of new bone formation within the macroporous BCP discs was then quantified by image analysis (Quantimet 4; Leica Microsystems).
Statistical analysis
Data are expressed as mean ± standard error of the mean and were analyzed by the nonparametric Kruskal-Wallis test. A two-sided p < 0.05 was considered statistically significant.
Results
Effect of serum concentration
After expansion, cells were cultured in differentiation media containing 2% or 10% FCS and Dex as the osteogenic factor. Proliferation and mineralization were assessed after days 7 and 21, respectively. As compared with day 0, at day 7, cells cultured with 10% FCS showed a significant increase in proliferation, whereas those cultured with 2% FCS showed lower mitogenic activity (Fig. 1A).
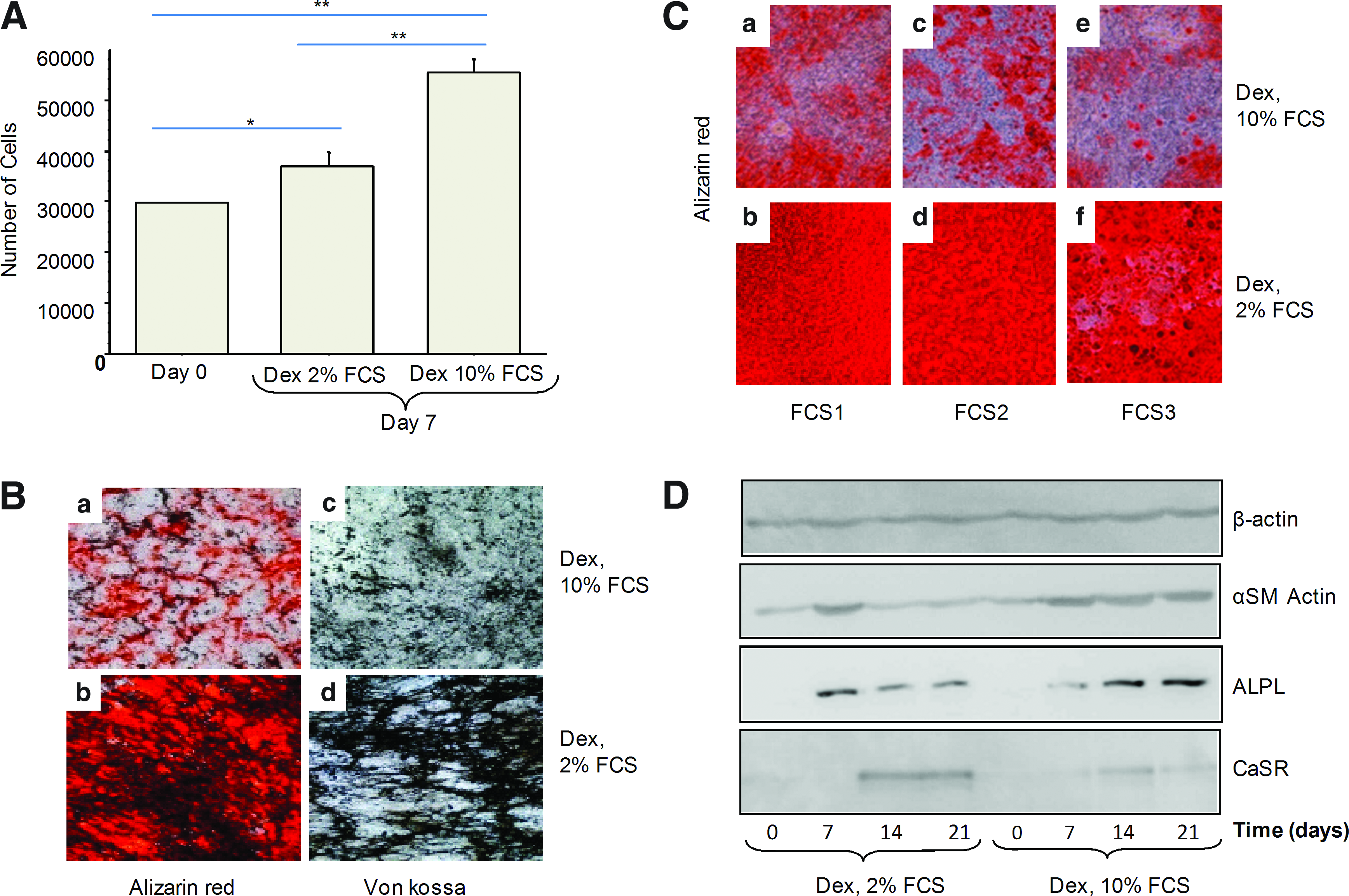
Effect of different FCS concentrations on cell proliferation of MSCs. Human MSCs were cultured in osteogenic medium (Dex) containing 10% or 2% FCS to evaluate their proliferation
With regard to mineralization, 2% FCS produced greater calcium and phosphate deposition than did 10% FCS (Fig. 1B). Similar effects were observed when using three serum batches from different donors (Fig. 1C).
We examined expression of several molecular markers delineating different phases of MSC osteoblastic differentiation. αSM actin is a well-known marker of immature proliferative MSCs, whereas ALPL is induced early in osteoblast maturation, and CaSR expression is detected later in the process.19–21 The level of αSM actin protein was decreased after 7 days of 2% FCS culture (Fig. 1D) but was constant with 10% FCS culture. In addition, with 2% FCS, the level of ALPL protein was transiently upregulated early during the differentiation process but was regularly increased with 10% FCS. However, the level of CaSR increased and remained high after 14 days with 2% FCS than with 10% FCS.
Potentiation of serum effect with osteogenic inducers
We determined whether the low serum effect could also be observed with other types of osteogenic inducers such as BMPs. Cells were cultured in media containing Dex or BMP2, 4, or 7. Because expression of osteoblastic proteins was observed early in culture, we chose day 10 as the endpoint for the following investigations. Flow cytometry revealed a higher ALPL expression level in cells cultured with 2% FCS, regardless of the other medium components (Fig. 2). The substitution of Dex by BMPs induced a further increase in ALPL expression, with BMP4 the most potent inducer.
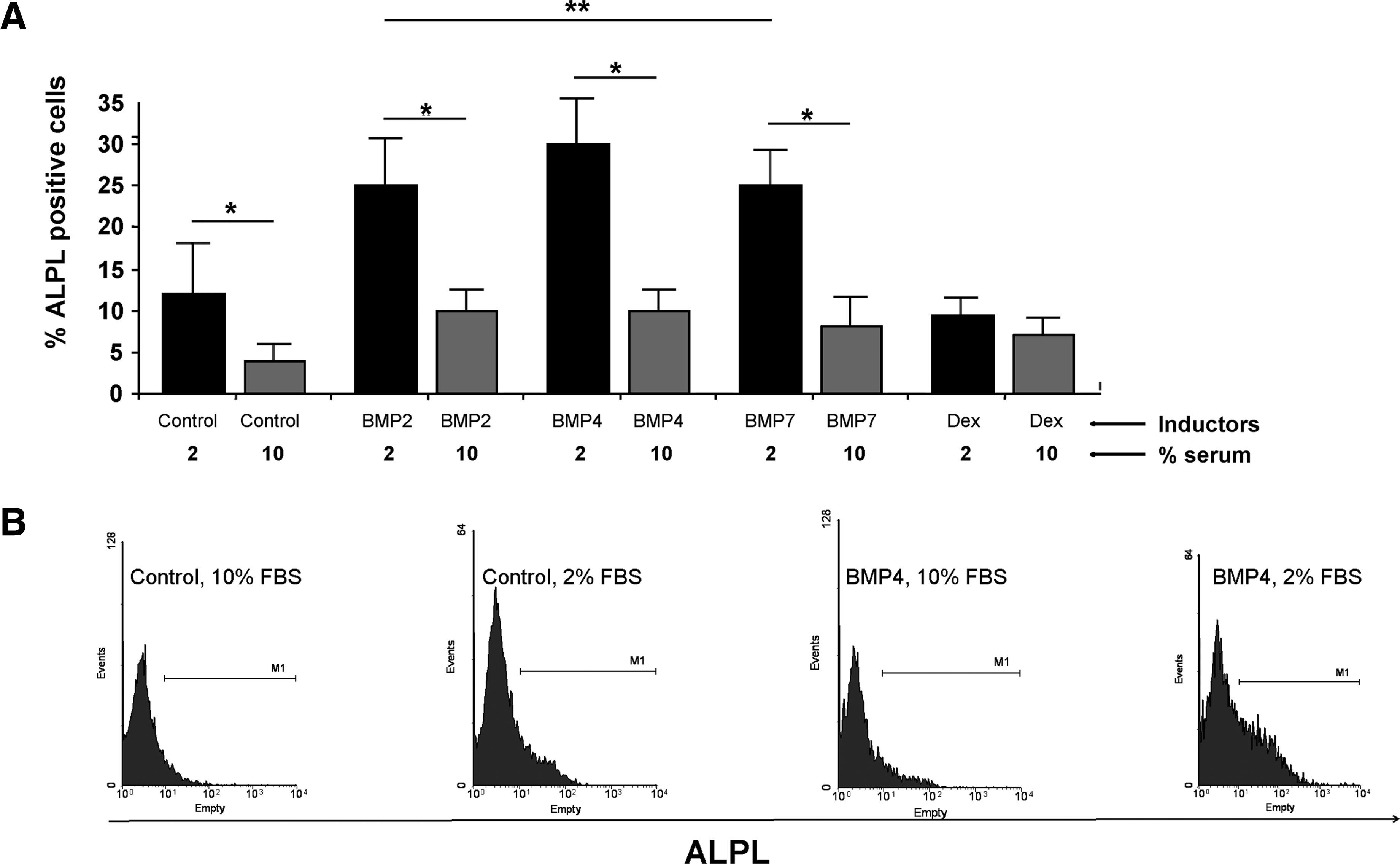
Flow cytometry analysis of ALPL protein expression. Human MSCs were cultured for 10 days in proliferation medium (control) or differentiation medium containing 2% or 10% FCS and different osteoblastic inducers (BMP2, 4, 7, and Dex).
Next, we determined expression of other osteoblastic markers with Dex or BMP4 and 2% FCS culture. Relative mRNA expression of early (ALPL) and late (PTHR1, BSP Osterix) osteoblastic markers was higher in cells cultured with BMP4-2% FCS than with Dex-2% FCS (Fig. 3A). These increases were accompanied by a decrease in αSM actin mRNA expression level, which indicated the loss of the immature state.
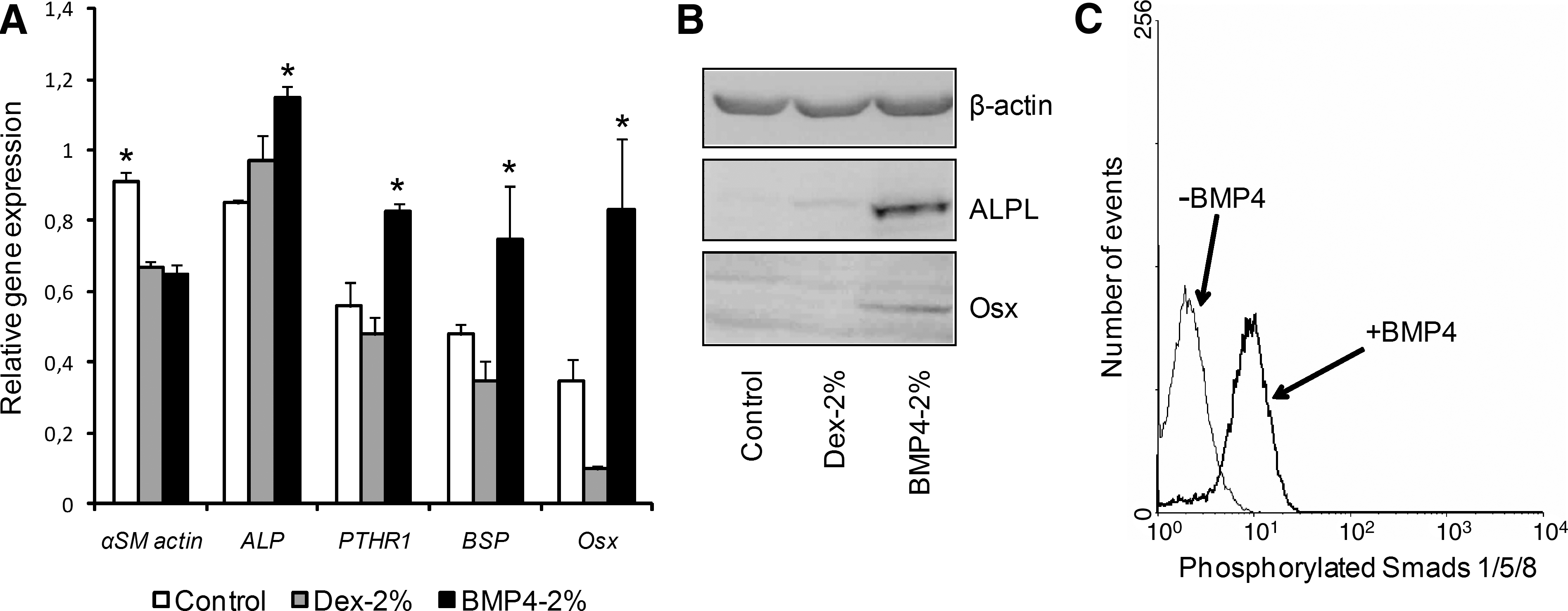
Osteogenic effect of BMP4 and 2% FCS on human MSCs. Cells were cultured for 10 days in proliferation medium (control, 10% FCS), osteogenic medium with 2% FCS (Dex-2% FCS), or BMP4 with 2% FCS (BMP4-2% FCS).
Protein expression of the markers after 10 days of culture was in agreement with mRNA expression results. Indeed, ALPL protein expression was higher with BMP4-2% FCS than with Dex-2% FCS (Fig. 3B). Further, BMP4-2% FCS but not Dex-2% FCS was able to induce Osterix expression after 10 days of culture. As well, the addition of BMP4 induced the phosphorylation of Smad1, 5 proteins, which demonstrates activation of the BMP signaling pathway (Fig. 3C). Therefore, osteoblastic differentiation induction is significantly more potent at low serum content and the addition of BMP4 regardless of Dex further increased osteogenesis.
Gene expression profile of differentiating cells seeded on ceramic scaffolds
We next asked whether benefits observed when cells were cultured on plastic could also be obtained when BCP biomaterials are used as scaffold. Thus, we checked mRNA expression of osteoblastic markers over 21 days of culture. ALPL gene expression with BMP4-2% FCS was increased until 21 days of culture and remained higher than that with Dex-2% FCS and the proliferation medium (Fig. 4). Similarly, BSP gene expression was higher with BMP4-2% FCS, although to a lesser extent than that for ALPL. Expression of late osteoblastic markers (i.e., Osteocalcin and Osterix) was low with control and Dex-2% FCS but high with BMP4-2% FCS. Therefore, we observed an upregulation of gene expression whatever the gene tested with BMP4-2% FCS used as the osteoinducer. Moreover, Osterix (the crucial transcription factor for osteoblastic maturation) was induced early with BMP4-2% FCS because its expression was detected at day 1 (Supplementary Fig. S2). Such upregulation was never detected with Dex-2% FCS. Taken together, these results further showed that the effect of BMP4-2% FCS on osteogenesis was substrate independent.
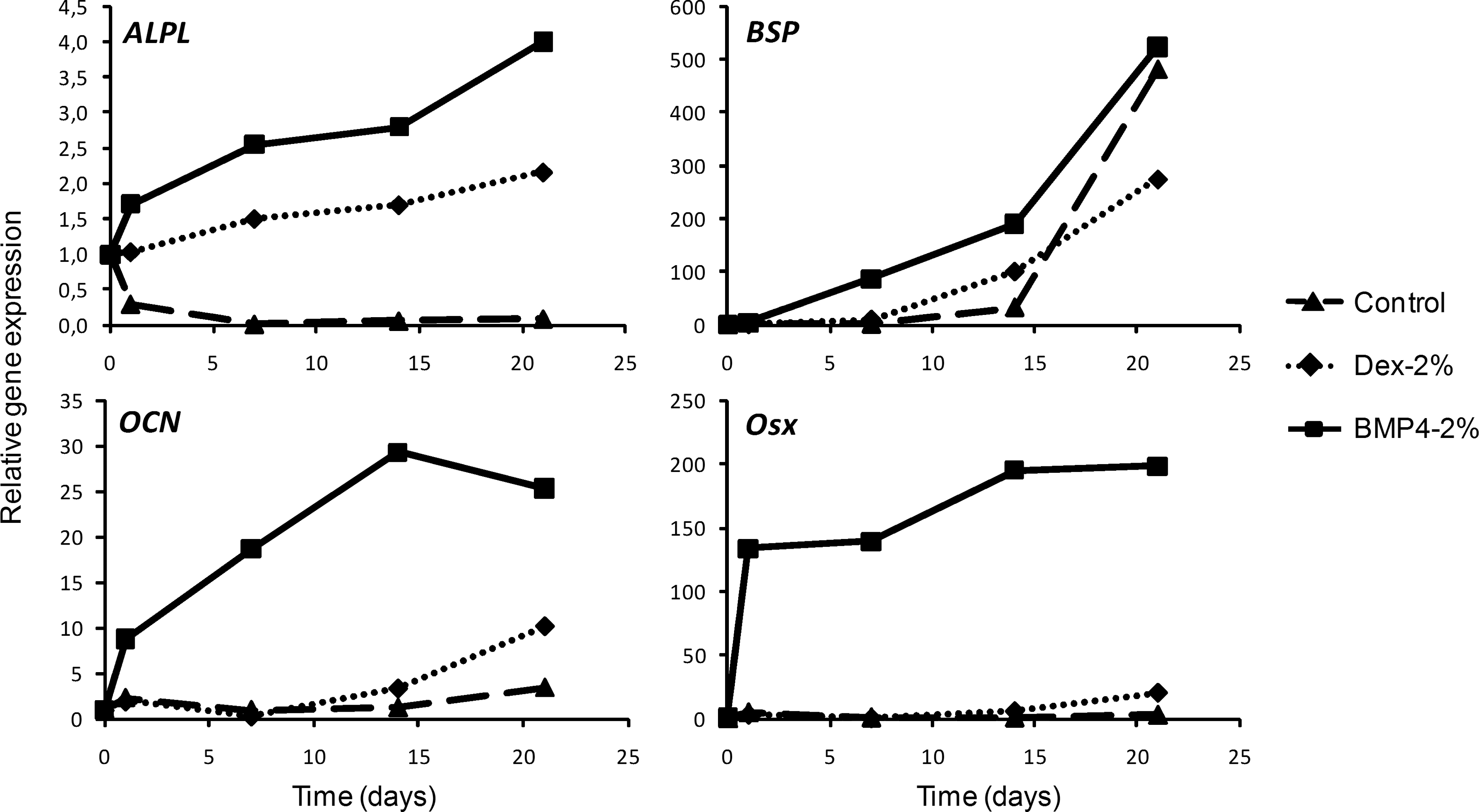
Expression profile during osteogenesis on BCP discs. Cells were cultured for 21 days on BCP discs in control 10% FCS or osteogenic (Dex-2% FCS or BMP4-2% FCS) media. Quantitative RT-polymerase chain reaction was performed on tissue nonspecific ALPL, BSP, OCN, and Osx mRNA. Gene expression was normalized to that of the control (cells before seeding on HA-TCP discs). Representative results of experiments with cells from three different donors. BCP, biphasic calcium phosphate; OCN, osteocalcin; Osx, osterix.
In vivo bone formation
To corroborate the consistency of in vitro results and assess the functional capacity of ex vivo-derived osteoblasts to form bone, differentiating cells were seeded on BCP scaffolds and implanted subcutaneously in nude mice. All osteogenic media evaluated in vitro induced bone formation ectopically (Fig. 5A). Bone tissue was found peripherally and within outer pores of the BCP discs. Newly formed bone was in direct contact with the BCP surface. The mineralized bone matrix enclosed numerous lacunae, which indicated the capacity of osteoblasts to terminally differentiate into osteocytes. However, Dex-2% FCS induced the lowest amount of bone formation within the ceramic discs (Fig. 5B). Conversely, BMP4-2% FCS resulted in the highest level of bone deposition, followed by the classical osteoblastic differentiation medium Dex-10% FCS. However, there were no significant differences in bone deposition between these two media.
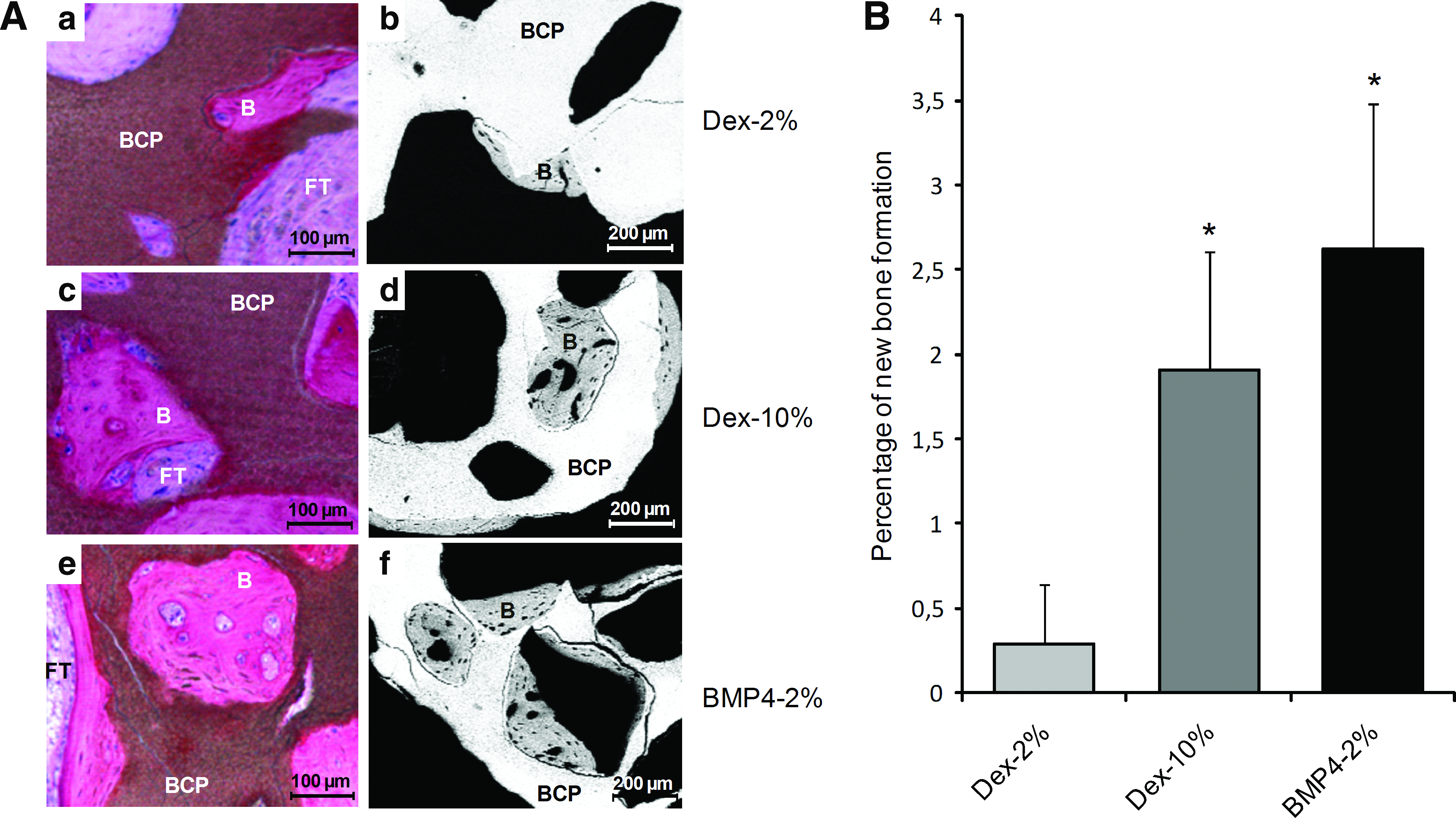
Evaluation of new bone formation in vivo. Cells cultured on ceramic discs during 7 days in differentiation medium were implanted subcutaneously for 8 weeks in nude mice.
Discussion
With the emergence of bone tissue engineering (BTE), promising results have been obtained in vivo in model animal experiments.22–26 However, convincing data have not been reported with human studies. Such absence can be linked to the lack of knowledge of the human MSC commitment into an osteoblastic lineage and the difficulties in maintaining this differentiation in vivo. The interests of BTE and regenerative medicine prompted us to find new methods to improve the speed of in vitro osteoblastic maturation with, at the origin, MSCs expanded in vitro with easy to use procedures.
Our method of reducing the serum content of the osteogenic medium and adding the osteogenic inducer BMP4 at a low concentration, 50 ng/mL, produced numerous functional osteoblasts in a short time. However, we observed that in addition to inducing MSC differentiation, the glucocorticoid-containing osteogenic medium (Dex-10% FCS) also induced high cellular proliferation. Cellular proliferation and differentiation are well known to be two opposite but interconnected phenomena.27–29 Notably, during early stages of osteogenesis, multipotent cells proliferate and increase their expression of extracellular matrix proteins. At that moment, genes inducing cell proliferation and cell cycle progression are predominantly expressed. At the next stages, cells slow their proliferation to initiate their maturation. Within these cells, postproliferative genes and those associated with the osteoblastic maturation are expressed, following that of the genes derived from the first stage. Finally, mature cells mineralize the extracellular matrix that they synthesized and reorganized previously Experimental approaches reducing the proliferative stage hasten the differentiation, and those extending the proliferation period delay the onset of differentiation.30–35 Hence, the decrease in osteoprogenitor proliferation is a crucial step for osteoblast differentiation. However, in vitro, the FCS medium currently used is designed to favor MSC expansion. Therefore, decreasing the FCS concentration could decrease the proliferation of osteoblastic cells and enhance their differentiation. Our results confirm such direct links notably through the FCS concentration in osteogenic media (Fig. 1). Indeed, low FCS concentration decreased expression of the immature state marker (αSM actin) and increased that of osteoblastic markers such as ALPL and CaSR. These results are in agreement with data reported by Katagiri et al., who demonstrated the need to decrease FCS content to achieve myogenic differentiation of the multipotent myoblastic cell line C2C12. 36
Several publications have illuminated the molecular mechanisms induced by a decrease in FCS content and highlight the crucial intervening proteins. Although a large variety of proteins contained in FCS enhance cellular survival and proliferation, other factors have a negative effect on the MSC commitment directly or indirectly. Among these proteins, TGF-β1 seems to play an important role in the inhibition of MSC differentiation. Maeda et al. demonstrated that TGF-β1 contained within serum inhibited the osteoblastic differentiation of both C2C12 cells and human MSCs and that the use of a TGF-β type I receptor inhibitor in high-serum culture conditions improved the differentiation process. 9 Similarly, other studies have elucidated the effect of TGF-β1 on the cell proliferation stimulation and differentiation inhibition notably by reducing the amount of cyclin-dependent kinase inhibitory proteins p57 and p27.37–39 Therefore, the decrease in proliferation of osteo-regenerative cells is required before their differentiation, and such decrease could be achieved simply by decreasing the FCS content in the medium.
To further enhance and hasten the low-serum effect on MSC induction into mature osteoblasts, several differentiation inductors were investigated. We used Dex as an osteoinducer, but its beneficial activities are still controversial, although in vivo it shows osteoporotic potential.40,41 Dex is a synthetic glucocorticoid hormone with a pleiotropic effect and is commonly used clinically for its antiinflammatory functions. When used chronically, glucocorticoids induce adverse effects such as bone loss, which results in higher fragility than with osteoporotic bones in postmenopausal women. 13 Further, Dex can decrease MSC mineralization when used at pharmacological doses. 42 Glucocorticoids are anti-inflammatory molecules, and Dex could disturb osteoclastogenesis and osteoclast function. 15 In addition, Dex is not restricted to osteoblastic differentiation and can be used to induce adipogenic and chondrogenic pathways, depending on other associated factors.7,43,44 Thus, according to the context, Dex could induce adipocytes as well as osteoblasts. Regarding osteoblastic differentation, continuous treatment with Dex can increase osteoblastic commitment of MSCs, but the cells are restrained to a premature osteoblastic or a less-differentiated state. 45 Other in vivo data demonstrated that glucocorticoids disturb osteoblastic functionalities in treated mice, with lower osteocalcin content than in control littermates and decreased amount of trabecular bone. 46 Therefore, Dex is still considered an osteoinducer with strong adverse effects, without consistent efficacies for generating functional osteoblasts.
In contrast to Dex, BMP2, 4, and 7 are physiological molecules involved in bone development and formation.47–50 BMPs also play a crucial role in chondroblastic and osteoblastic differentiation of uncommitted progenitor cells,51–53 and these properties of BMP2 and BMP7 have been tested clinically to treat several bone abnormalities such as delayed union or nonunion postfracture and to improve spinal fusion.54,55 Although some in vitro and in vivo studies showed BMP2, 4, and 7 to have difference biological responses in inducing bone formation, no data have yet demonstrated sharp differences in their molecular pathways. However, among the BMPs tested in our culture conditions, BMP4 emerged as the most efficient osteoblastic inducer at low FCS concentration. mRNA and protein expression of ALPL, PTHR1, and Osterix was greatly enhanced even after a short period (10 days) with BMP4-2% FCS, confirming previous data.56,57 Such induction is crucial to generate true mature osteoblasts because Osterix−/− mice have normal cartilage but lack bone formations. 58 Physiologically, BMP4 acts as a neo-osteogenic inducer, 59 and several studies of BMP4 have demonstrated its major role during fracture healing.60–63 BMP4 was also shown to induce chondrogenesis, but we never observed chondroblasts in cultures, and more importantly, genes associated with chondrogenesis were downregulated (data not shown). Recent in vivo studies suggested that BMP2 and 4 have crucial roles in skeletogenesis because their deletion resulted in impaired bone formation without disturbing chondrogenesis. 64 However, in other in vivo models, a conditional BMP4 defect had no effect on bone reconstitution postfracture in adult mice. 65 Therefore, the role of BMP4 in neo-osteogenesis in adults and in skeletogenesis is crucial but remains to be further defined.
The strong osteoblastic stimulation we observed with BMP4 and low FCS culture could be of utmost importance for BTE at different levels: (1) depending on pathological context and on anatomical sites (e.g., limbs vs. mandibles), it could be more judicious to inject committed osteoblastic cells than immature MSCs. Indeed, some published data showed an improvement of bone/cartilage formation for MSCs when treated in vitro by osteoinducers before be implanted or when MSCs were coinjected with osteoinducers66–68 ; (2) in the cases where noninduced MSCs should be used, it could be interesting to evaluate ex vivo their osteogenic potentials and such evaluations need to have consistent in vitro methods. Similarly, these methods could be also performed to assess bone formation-supporting capabilities of new biocompatible materials.
Therefore, osteoblasts differentiated from MSCs must be as mature as possible and remain firmly engaged in this direction after implantation to form bone. In our experiments, human MSCs cultured on ceramic scaffolds and exposed simultaneously to low FCS concentration and BMP4 were undeniably engaged in a more mature osteoblastic state. Such commitment was shown by high ALPL and BSP gene expression as compared with expression with Dex-2% FCS incubation or proliferation media. Moreover, as compared with other media, BMP4-2% FCS boosted the differentiation process from day 1 of culture. Notably, we observed strong mRNA expression of late osteoblastic markers such as Osteocalcin and Osterix at this early time point.
Such effective osteoblastic differentiation obtained with BMP4-2% FCS was efficiently transcribed in vivo because of the amount of bone formation we found. Unlike the canonical osteoblastic medium (Dex-10% FCS), with BMP4-2% FCS, the amount of bone formation in vivo related strongly to the quality of osteoblastic differentiation observed in vitro. These results underline the controversial role of glucocorticoids in bone formation. Indeed, Dex-2% FCS successfully commits MSCs toward osteoblastic differentiation in vitro but surprisingly generates a low amount of bone deposition in vivo, which suggests a disturbance of osteoblastic functionality that could be related to the ineffective maturation in vitro. 45 On the other hand, cells cultured in Dex-10% FCS medium gave rise bone in vivo similarly to those treated by BMP4-2% FCS conditions. This result could be explained by the fact that cells cultured into Dex-10% FCS medium still proliferated unlike cells cultured in Dex-2% FCS. Therefore, Dex-10% FCS conditions could increase the number of osteogenic cells before be implanted improving bone formation potentials of biomaterials. This highlights the need to define the quantities of osteo-regenerative cells colonizing biomaterials before be implanted. In addition, their differentiation state seems crucial for bone formation in vivo. We described above some data showing the benefits to commit cells before their injection. Therefore, we can hypothesize that efficacy of a graft to regenerate bone is dependent on the number of differentiated cells contained within it. Data reported by Gronthos et al. support such assumption. Indeed, the use of telomerized MSCs demonstrated more ectopic bone formation potential in immuno-compromised mice than nontelomerized cells. These benefits were related to their proliferation rate and also to their high content of osteogenic molecular factors Runx2 and osterix. 69
In this work, we improved the osteoblastic differentiation of MSCs by reducing the FCS concentration in the culture medium. Concomitantly, we achieved fast and mature osteoblastic differentiation by substituting Dex with the osteogenic factor BMP4 in the medium. Unlike the standard differentiation medium, this alternative approach conferred consistent results between in vitro osteoblastic differentiation and effective in vivo bone formation. Such a relationship is crucial when studying bone biology to upgrade protocols for clinical use. The induction of osteoblasts from MSCs with low FCS concentration and BMP4 might be of interest for BTE and regenerative medicine. In addition, these latter aims prompt research labs to develop expansion media without animal sera and preliminary results are promising. Therefore, it could be very interesting to evaluate the bone formation by using BMP4 into serum-free medium or media containing human platelet lysates.70–73 Nevertheless, these media favored the proliferation of multipotential cells. We assume that such conditions should be modified with regard to data described here.
Footnotes
Acknowledgments
This work was funded by the Agence Nationale de la Recherche Technologies pour la Santé et l'Autonomie, ATOS project no. 024-03 (2007–2010). The research leading to these results has also received funding from the European Commission Seventh Framework Programme (FP7/2007–2013) under the Grant agreement no. 241879, through the REBORNE project. The authors would like to acknowledge Paul Pilet for technical assistance in microscopy and microtomography.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.