
Abstract
Chronic kidney disease (CKD) is a global health problem; the growing gap between the number of patients awaiting transplant and organs actually transplanted highlights the need for new treatments to restore renal function. Regenerative medicine is a promising approach from which treatments for organ-level disorders (e.g., neurogenic bladder) have emerged and translated to clinics. Regenerative templates, composed of biodegradable material and autologous cells, isolated and expanded ex vivo, stimulate native-like organ tissue regeneration after implantation. A critical step for extending this strategy from bladder to kidney is the ability to isolate, characterize, and expand functional renal cells with therapeutic potential from diseased tissue. In this study, we developed methods that yield distinct subpopulations of primary kidney cells that are compatible with process development and scale-up. These methods were translated to rodent, large mammal, and human kidneys, and then to rodent and human tissues with advanced CKD. Comparative in vitro studies demonstrated that phenotype and key functional attributes were retained consistently in ex vivo cultures regardless of species or disease state, suggesting that autologous sourcing of cells that contribute to in situ kidney regeneration after injury is feasible, even with biopsies from patients with advanced CKD.
Introduction
Kidney transplantation is the definitive standard of care for CKD, providing better long-term survival 3 and cost-effectiveness 4 than dialysis; however, there is a chronic shortage of organs. Despite increases in both cadaveric and living kidney donors, the rate of transplantation per 100 dialysis patient-years in the United States is actually decreasing. 5 These realities underscore the need for new treatment paradigms that can alleviate some of the healthcare burden imposed by dialysis and transplant by providing substantial and durable augmentation of kidney functions to slow progression of CKD. Regenerative medicine technologies hold promise for providing next-generation therapeutic options for CKD, especially if they can be initiated effectively after the onset of progressive disease.
Regenerative medicine approaches that combine autologous cells derived from the target organ (also known as an “autologous/homologous cell sourcing strategy” 6 ) with biomaterials have transitioned to clinical trials designed to augment bladder, a tubular organ.7,8 We sought to investigate the feasibility of applying a similar strategy to kidney, a solid organ, in the experiments reported here. The choice of unfractionated primary kidney cells (UNFX)9,10 as a starting population was based on literature support of the therapeutic potential for augmenting renal function in the context of CKD when administered after the onset of disease. In a rodent model, chronic renal disease was established by hemi-nephrectomy and ischemia to the intact kidney. 11 Injection of a heterogeneous mixture of human fetal kidney cells, containing embryonic tubular cells, via the vena cava into rats with established disease led to transient improvement of all functional parameters. The recovery observed in rats receiving fetal kidney cells was more rapid in the context of chronic disease state than that observed in rats receiving bone marrow-derived mesenchymal stem cells. When acute damage was superimposed in the model, the fetal kidney cells showed therapeutic benefit at a level equivalent to that observed with bone marrow-derived mesenchymal stem cells.
One subpopulation of renal cells that has been repeatedly associated with restoring renal function after acute renal injury in both preclinical and clinical studies is tubular cells. After acute injury, resident, surviving tubular cells dedifferentiate and proliferate to replace the lost cells throughout the nephron. 12 In a series of experiments designed to assess the effects of transplanted bone marrow cells in restoring renal function in a model of chronic renal injury that leveraged genetic tools for discerning resident from transplanted cells, resident tubular cells were observed to be key participants in the recovery process.13–15 Finally, phase II clinical trials of an extracorporeal renal tubule assist device, which combined a bioreactor containing living proximal tubular cells with conventional hemodialysis, demonstrated that the bioreactor-augmented dialysis procedure conferred a significant survival benefit and more rapid restoration of kidney function in patients with acute renal failure. 16
An autologous/homologous cell sourcing strategy for developing cellular components of candidate therapies for CKD could be considered feasible if methods for isolating, characterizing, and expanding (ICE methods) defined renal cell populations that had the potential for conferring therapeutic benefit could be designed and applied to disease-relevant animal models and humans. 6 Of particular relevance to feasibility are ICE methods that yield target cell populations from patients with CKD using a clinically relevant tissue harvesting procedure. Developing these ICE methods for UNFX and a tubule-enriched subpopulation and applying them to rodent, canine, and human tissues was the goal of this study. Additionally, we limited our evaluation to protocols that are compatible with process development and scale-up.
Healthy canine and human kidney tissue samples were used to demonstrate that ICE methods yielding rodent UNFX and B2, a tubule-enriched subpopulation of UNFX, could be adapted successfully to a relevant preclinical species (dog) and the clinically relevant human. Proof of principle for isolation and expansion of UNFX and generation of B2, a tubule-enriched subpopulation of UNFX, from diseased kidney tissue was achieved in the ZSF1 (fa/facp) rat model, which is a metabolic disease model characterized by overt and progressive kidney disease secondary to obesity and type 2 diabetes. 17 Finally, whole kidney tissue was collected from five human organ donors with advanced CKD to demonstrate that functional B2 cells were present in the tissue and could be isolated and propagated successfully, even from small needle core biopsies. The data presented herein provide evidence that an autologous/homologous cell sourcing strategy for UNFX and B2 renal cell populations is translatable to large mammals and to humans with CKD, thus supporting the feasibility of these ICE methods for obtaining defined renal cell populations with potential for conferring therapeutic benefit in the treatment of CKD.
Materials and Methods
Tissue sourcing
Lewis and ZSF1 tissues were sourced from Charles Rivers Laboratories and canine tissue from the Integra Group in compliance with policies governing the use of laboratory animals. With the exception of HK023, human tissue was sourced through the National Disease Research Institute in compliance with policies governing the use of human tissues for research. Human specimen HK023 was obtained as biopsy material under informed consent with approval from the Institutional Review Board of the University of North Carolina at Chapel Hill.
Cell isolation, culture, and separation
Unfractionated heterogeneous primary renal cell cultures (UNFX) were established and maintained as described for all species.9,10 For human samples initiated from biopsy, 2 mm core biopsies (0.02 g tissue) were collected with an Easy Core™ Biopsy System (Boston Scientific) and washed 2 × in Hanks balanced salt solution (without magnesium chloride–MgCl2–and calcium chloride–CaCl2), minced, and digested for 30 min in Dispase (Stem Cell Technologies) containing 300 U of collagenase type IV (Worthington Biochemical Corporation) and 5 mM calcium chloride (Sigma). Cells were cultured in 35 mm tissue culture dishes (Corning/Costar). Cell yields were determined by counting viable cells at passage and growth rates were calculated as ln(fold increase)/time. Population doubling times were calculated as ln(2)/growth rate.
Cell subpopulations from dog and human were obtained from UNFX cultures that were placed into low-oxygen (2%) conditions for 24 h before harvest via trypsin, which facilitates the subsequent distribution of cell subpopulations and enhances specific phenotypic attributes (unpublished observations). Two-milliliter suspensions of 60–75 × 106 UNFX cells were loaded onto four-step iodixanol (OptiPrep; 60% w/v in unsupplemented Keratinocyte Serum Free Media; Invitrogen) density gradients layered specifically for rodent (16%, 13%, 11%, and 7%), canine (16%, 11%, 10%, and 7%), or human (16%, 11%, 9%, and 7%). Gradients were centrifuged at 800g for 20 min at room temperature (without brake). Bands of interest were removed via pipette and washed 2× in sterile phosphate-buffered saline (PBS).
Gene expression analysis
RNA isolation and cDNA preparation
Tissue or cell samples were homogenized using the QIAshredder (Cat #79656; Qiagen) and RNA was isolated using an RNeasy Plus Mini isolation kit (Cat # 74134; Qiagen). RNA integrity was verified and samples were quantified via spectrophotometric analysis. 18 cDNA was synthesized using the Superscript® Vilo cDNA synthesis kit (Cat# 11754-250; Invitrogen).
Quantitative reverse transcriptase-polymerase chain reaction assays
Expression of erythropoietin (EPO), E-cadherin (ECAD), N-cadherin (NCAD), cubilin, 1 alpha, 25-dihydroxyvitamin D3-24-hyroxylase (CYP24), aquaporin-2 (AQP2), nephrin (NEPH), vascular endothelial growth factor, and CD31 (PECAM) were examined via quantitative real-time polymerase chain reaction (PCR) using TaqMan Gene Expression Master Mix (Cat #4369016), catalogued TaqMan probes and primer sets (Table 1) from Applied Biosystems, and an ABI-Prism 7300 Real-time PCR System.
n/a, not applicable.
Determination of relative quantification values
Relative quantification values were calculated according to manufacturer's protocols by 7300 System SDS Software version 1.4.0.25 (Applied Biosystems). Gene expression in UNFX cultures was quantified relative to cDNA amplified from peptidylprolyl isomerase B (for rodent genes), TATA binding protein (for canine genes), or peptidylprolyl isomerase A (PPIA) (for human genes). Gene expression in B2 and B4 subpopulations was quantified relative to the same cDNA amplified from a sample of the corresponding UNFX input culture used to generate the subpopulations. Reproducibility of B2 gene expression patterns among human tissue biopsies was measured as the fold change in expression of target genes in comparison to endogenous control (PPIA). Briefly, fold change cycle threshold (Ct) values of samples were generated by 7300 System SDS Software version 1.4.0.25 An average Ct value for the endogenous control (PPIA) was calculated for B2 samples across all plates. ΔCT for B2 target genes was calculated as Ct(target gene) – AvgCt(endogenous). Fold change for the samples was calculated as 2−ΔCt.
Flow cytometry
Phenotypic characterization of UNFX and B2 cells isolated from rodent, canine, and human tissues was performed by standard flow cytometric methods 19 on a FACS Aria (Becton Dickinson). Data were analyzed with FlowJo (Tree Star) software. Primary antibodies (used at 1 μg/mL) and isotype-matched controls (used at 1 μg/mL) are listed in Table 2. Briefly, cells fixed in 2% paraformaldehyde were washed 2× in sterile PBS and stained with primary antibodies at a concentration of 1 μg/106 cells/mL and incubated for 2 h at 4°C. After incubation with primary antibody the cells were washed 2× and stained with secondary antibody at a concentration of 1 μg/106 cells/mL and incubated for 30 min at room temperature protected from light. After incubation the cells were washed 2× and subjected to flow cytometry.
GGT, gamma glutamyl transpeptidase; ms, mouse; rb, rabbit.
Albumin uptake assay
Renal proximal tubular cell function was assessed by the activation and inhibition of albumin endocytosis mediated by specific proximal tubule receptors, megalin and cubilin, as described. 20 Cultures were grown to confluence (7–10 days) and assayed 1–2 days post-confluence. Eighteen to 24 h before assay initiation, the growth medium was replaced with assay buffer consisting of serum-free, phenol red-free, low-glucose (1 g/L) Dulbecco's modified Eagle's medium containing 2 mM glutamine, 10 mM HEPES [4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid] buffer (all from Invitrogen). On the day of assay, cells were washed twice with assay buffer consisting 1.8 mM CaCl2 and 1 mM MgCl2, and incubated with assay buffer for 30 min in a humidified chamber at 37°C/5% CO2 (carbon dioxide) to ensure adequate exposure to cofactors. Cells were then exposed to a final concentration of 10–30 μg/mL rhodamine-conjugated human albumin (ALB-RHO) (Abcam, Inc.) for 30 min at 37°C/5% CO2. Wells were washed three times with ice cold PBS to stop endocytosis and fixed immediately with 2% paraformaldehyde containing 10 μg/mL Hoechst nuclear dye (Invitrogen). Addition of 1 μM receptor-associated protein (recombinant human; Innovative Research™), a specific competitive inhibitor of megalin:cubilin-mediated albumin uptake 20 in control cultures served to demonstrate specificity of the reaction. Inducibility of ALB-RHO endocytosis was examined by pretreatment of UNFX cultures with 10−8 M human angiotensin II (Sigma-Aldrich) for 4 h before exposure to ALB-RHO, as described. 21 Cells were observed and images captured via microscopy using a BD Pathway 855 High-Content BioImager (Becton Dickinson) and quantitative image analysis was conducted with IN Cell Investigator software with Multi-Target Analysis Module (GE Healthcare).
Measurement of hydrolase activity
Enzymatic activity of two proximal tubule-associated hydrolases, gamma glutamyl transpeptidase (GGT or GGT1 for the gene product) and leucine aminopeptidase (LAP), was measured in human UNFX cultures at passage 1. Established cultures were trypsinized and replated at 4000 cells/cm2 and maintained in standard growth conditions for 7 days before assay. The GGT assay was adapted from published methods. 22 Briefly, wells were washed and 0.5 mL of substrate reagent comprised of 2.5 mM L-glutamic acid γ-p-nitroanilide (Sigma) in Tris-HCl (pH 8.6) containing 150 mM NaCl and 50 mM glycylglycine was added for 1 h at room temperature. GGT activity was calculated as mean absorbance at 405 nm (corrected with reagent blank) of triplicate samples. Leucine aminopeptidase activity was determined in a similar assay using 3 mM L-leucine p-nitroanilide in PBS as substrate reagent. LLC-PK1 cell lysates (ATCC) served as positive controls. 23
Isoelectric focusing and western blotting for detection of EPO
Frozen whole kidney tissue from human kidneys HK16, HK17, HK18, HK19, HK20, and HK21 was used for protein sample collection. All tissues were lysed for 15 min at room temperature in a buffer consisting of 50 mM Tris (pH 8.0), 120 mM NaCl, 0.5% NP40, and protease inhibitor cocktail (Roche Applied Science) with rocking followed by centrifugation for 10 min at 13,000 RPM. Protein concentrations were determined from supernatants by Bradford Assay. 18 Lysate prepared from cultured human foreskin fibroblasts served as a negative control. Proteins in each sample were separated by isoelectric focusing (IEF) by adding 40 μg of protein per well to pH 3–10 IEF Gels (Invitrogen). Colorimetric IEF marker protein standards (10 μg/lane; Serva Electrophoresis; Cat# 39212) were loaded in two lanes, one on either side of the sample lanes. The gels were electrophoresed for 1 h at 100 V followed by 1 h at 200 V and finally 30 min at 500 V in pH 3–10 cathode and anode buffer (Invitrogen). The proteins were then transferred to a nitrocellulose membrane using the I-Blot system (Invitrogen) following the manufacturer's instructions. Transfer quality was assessed visually on the membrane using the colorimetric standard bands at pH 4.5, 4.2, and 3.5 (Epo protein runs at pH 4.2). After transfer, the membrane was blocked with 15 mL of 4% w/v low-fat milk dissolved in Tris Buffered Saline with 0.1% Tween-20 (TBS-T) (Sigma) for 2 h at room temperature with rocking. The membrane was probed overnight at room temperature with anti-human EPO monoclonal IgG2a MAb 2871 (R&D Systems) at a 1:600 dilution in 5 mL TBS-T with 2% w/v low-fat milk. The membrane was washed three times for 10 min each wash with TBS-T, and then probed with an horseradish peroxidase-conjugated rabbit anti-mouse IgG (Zymed) at a 1:50,000 dilution (within manufacturer's recommended range for detection with ECL Advanced Chemiluminescent reagent) in TBS-T with 2% w/v low-fat milk for 1.5 h at room temperature with rocking. The membrane was washed three times for 10 min each wash in TBS-T, followed by two 10-min washes in deionized water. The blot was developed using ECL Advance chemiluminescent reagent (GE Healthcare) following the manufacturer's instructions.
Renal histology
Representative kidney samples were collected and placed in 10% buffer formalin for 24 h. Sections were dehydrated in ascending grades of ethanol and embedded in paraffin. Paraffin blocks were cut at 5-μm thick sections, mounted on charged slides and processed for hematoxylin and eosin, Masson's trichrome and Periodic Acid Schiff staining (PAS) in accordance with standard staining protocols. 24 Digital microphotographs were captured at total magnification of ×40, ×100, and ×400 using a Nikon Eclipse 50i microscope fitted with a Digital Sight (DS-U1) camera. Renal morphology changes were assessed adopting commonly used 25 severity grade schemes (grades 1, 2, 3, and 4), to which descriptive terms (minimal, mild, moderate, and marked/severe) are applied to determine the degree of glomerulosclerosis, tubular atrophy and dilatation, tubular casts, and interstitial fibrosis and inflammation.
Statistical analyses
Basic statistics were calculated using Excel 2003 (Microsoft Corporation).
Results
UNFX composition in normal rodent, canine, and human kidneys
UNFX is a heterogeneous mixture of renal cells isolated from the cortical and medullary zones. UNFX isolates contain cells from major compartments of the kidney and retain functional attributes (e.g., bioresponsive erythropoietin expression, receptor-mediated albumin uptake, and the ability to recapitulate renal structures in three-dimensional culture).9,10,26 Flow cytometric analyses of representative cultures demonstrated that UNFX isolates from rodent, canine, and human normal kidneys were composed predominantly of CK8/18/19(+) epithelial cells and contained significant numbers of GGT(+) tubular cells and AQP2(+) ductular cells (Table 3). Although initial yields of UNFX from human were lower than those observed from rodent and dog, cultures were established successfully from 3/3 human donors and had a predominantly epithelial phenotype (Table 3). Human and dog UNFX cultures had fewer nonepithelial cells (CK8/18/19-negative) than rodent UNFX and cultures derived from all species contained small proportions (<5%) of endocrine, vascular, and glomerular cells as well as some undefined cell types (data not shown).
Tissue was sourced from Lewis rat, canine (mongrel) >6 months old, and human whole kidney samples and UNFX cells were isolated. Cell yields were determined at primary isolation and normalized to gram of tissue based on the wet weight of tissue processed. Relative percentages of epithelial (CK8/18/19+), tubular (GGT1+), ductular (AQP2+), and vascular (PECAM+/CD146+) cells were measured by flow cytometry of primary (Lewis rat and canine) or passage 2 (human) UNFX cultures.
UNFX, unfractionated primary kidney cells.
Isolation of defined subpopulations from established cultures of UNFX
Iodixanol gradients are used in the processing of primary autologous cells for clinical trials.7,8,27 Four-step density gradients, optimized for each species, separated UNFX cells into five subpopulations, designated B1 to B5. Detailed gene array analysis of rodent cells revealed that the B2 subpopulation was enriched for tubular epithelial cells (see Supplementary Data available online at
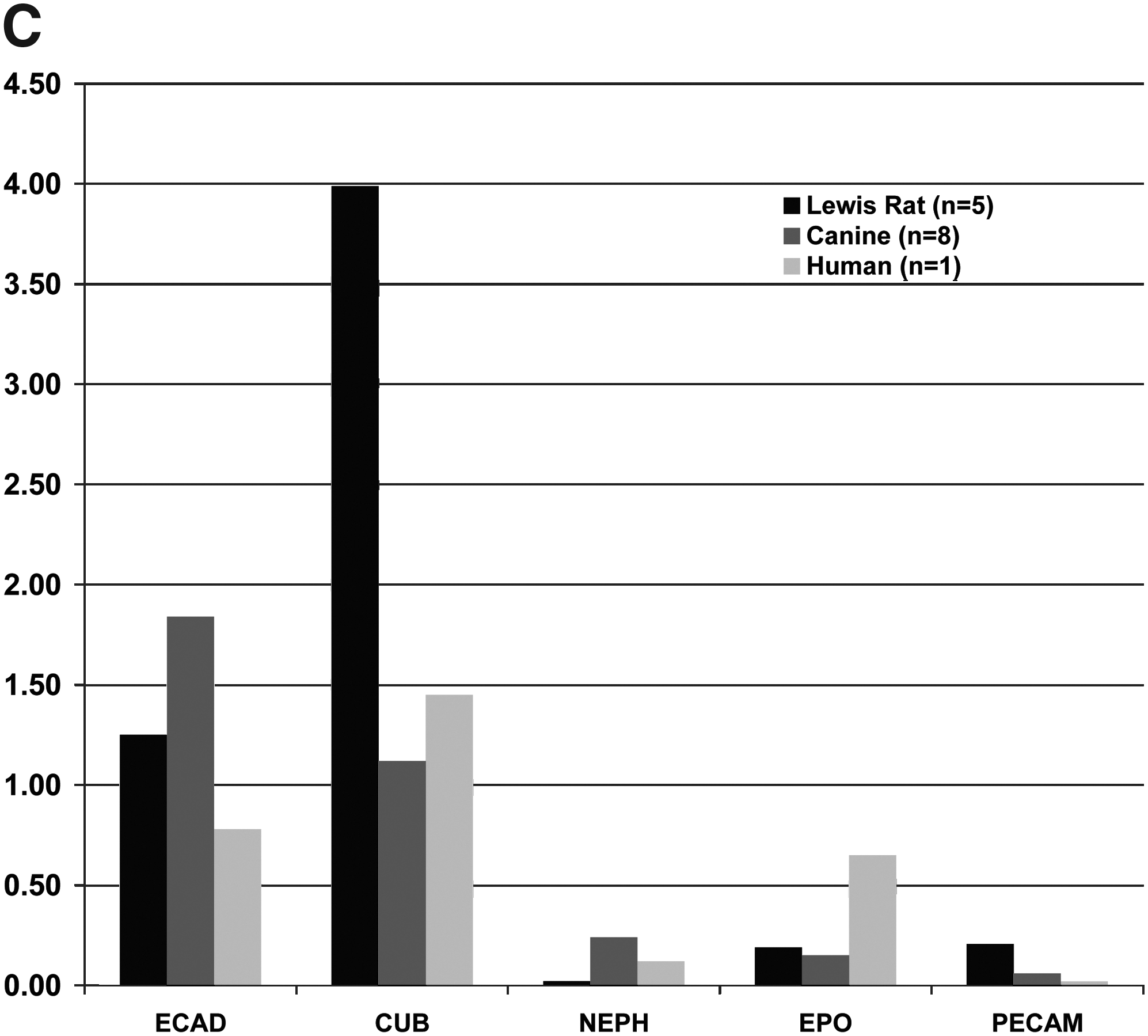
B2 and B4 subpopulations were generated from UNFX cultures derived from Lewis rat (passage 0), canine (passage 0), and human (passage 2) kidneys. Reported for each species are
n = 2.
n = 1.
nd, not determined.
Five markers were selected to characterize B2 subpopulations by flow cytometric analysis of protein expression (Fig. 1B) and gene expression relative to the tubule-depleted B4 fraction by quantitative reverse transcriptase (RT)-PCR (Fig. 1C): E-cadherin, CK8/18/19, GGT1 (protein only), AQP2, PECAM (CD31), and NEPH (gene expression only). For protein expression in canine tissues, substitute antibodies used for flow cytometric analysis were a pan-cadherin antibody, E-cadherin, and a CD146 antibody for PECAM. These characterization profiles were used to translate ICE methods to diseased tissues from rodents and humans with established CKD. Comparative gene expression analyses of B2 and B4 among samples confirmed the epithelial/tubular nature of B2 from rodent, dog, and human, and confirmed the segregation of vascular, EPO-expressing, and glomerular cells to B4 (Fig. 1C). The selected B2 subpopulation was more homogeneous than UNFX. The relative proportion of epithelial cells (i.e., ECAD+, Pan-Cadherin+, or CK8/18/19+) was increased to >90% in B2 across species, but the most notable distinction between UNFX and B2 was the enrichment in B2 of the proportion of GGT1+ tubular cells—ranging from a 1.4-fold increase in the dog to >2.0-fold increase in the human (compare Table 3 with Fig. 1B).
UNFX and B2 cells from diseased rodent kidneys
Obese ZSF1 (fa/facp) rats develop CKD by 8 weeks of age, whereas lean ZSF1 rats retain near normal renal structure (Fig. 2A) and function.17,28–30 UNFX cell suspensions were prepared from 10 to 35-week-old obese and lean ZSF1 rats (Fig. 2B) and subpopulations B2 and B4 isolated (Fig. 2C). Of note was the reduction in the B4 component in ZSF1/obese UNFX cultures (0.5%) compared to ZSF1/lean (3.1%). The resulting B2 subpopulation profiles (Fig. 2D, E) also showed some variability (consistent with the reduction in B4) but demonstrated that B2 from ZSF1/obese diseased tissue was a predominantly tubular epithelial population with characteristics similar to B2 isolated from ZSF1/lean kidneys.
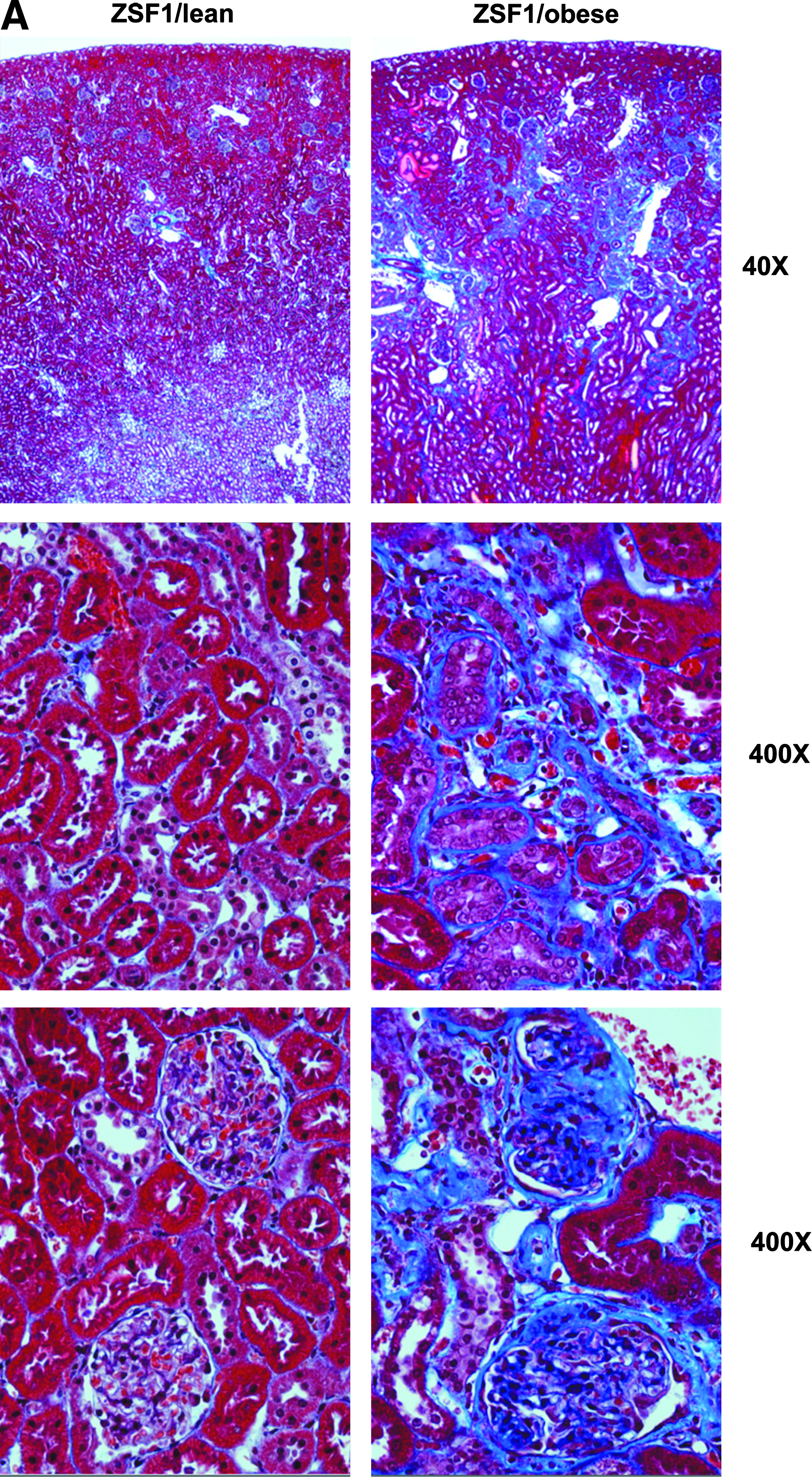
Tissue was sourced from ZSF1/lean rats (8 weeks old), ZSF1/obese rats (10–35 weeks old).
n = 1.
UNFX and B2 cells from humans with CKD
Eight human kidneys were obtained via approved organ procurement procedures (Fig. 3A): three from patients without CKD and five from patients with CKD. Renal disease was confirmed by serum chemistry at tissue harvest and histopathology (HK18 tissue in Fig. 3B; histopathology of all CKD samples available online as Supplementary Data). Severity of CKD in human patients varied, but without exception all human CKD patients were characterized by significant elevations in blood urea nitrogen (BUN) and serum creatinine (sCREAT) and were either anuric or had significant proteinuria.
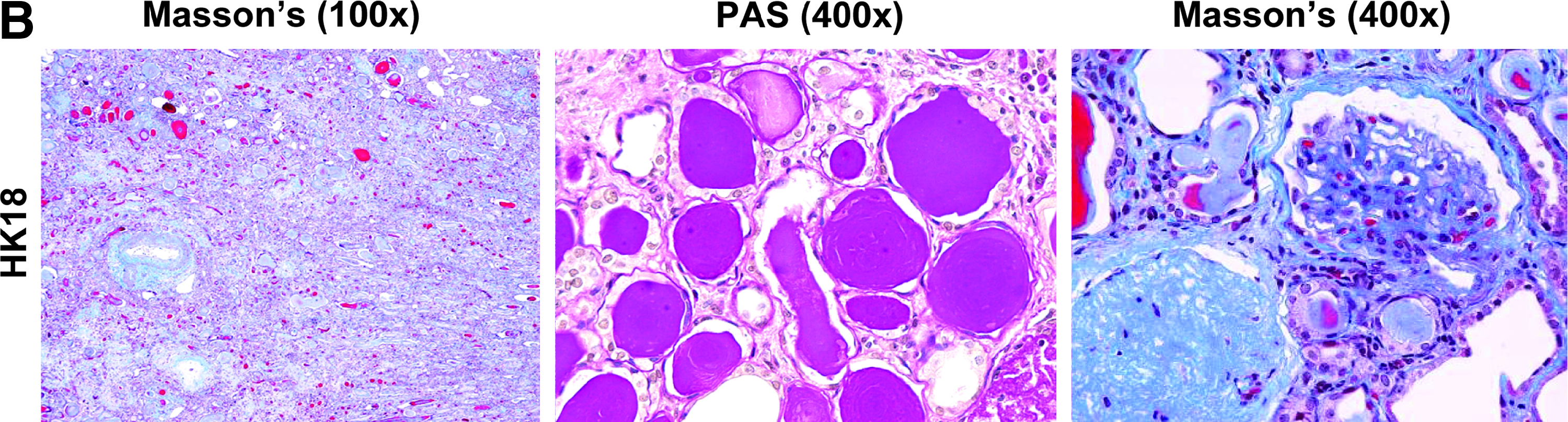
Human kidney tissue.
Detailed characteristics of kidneys procured are available online as Supplementary Data.
Obtained as biopsy sample from native kidney by informed consent from patient undergoing kidney transplant.
Isolation and functional characterization of UNFX from human CKD kidneys
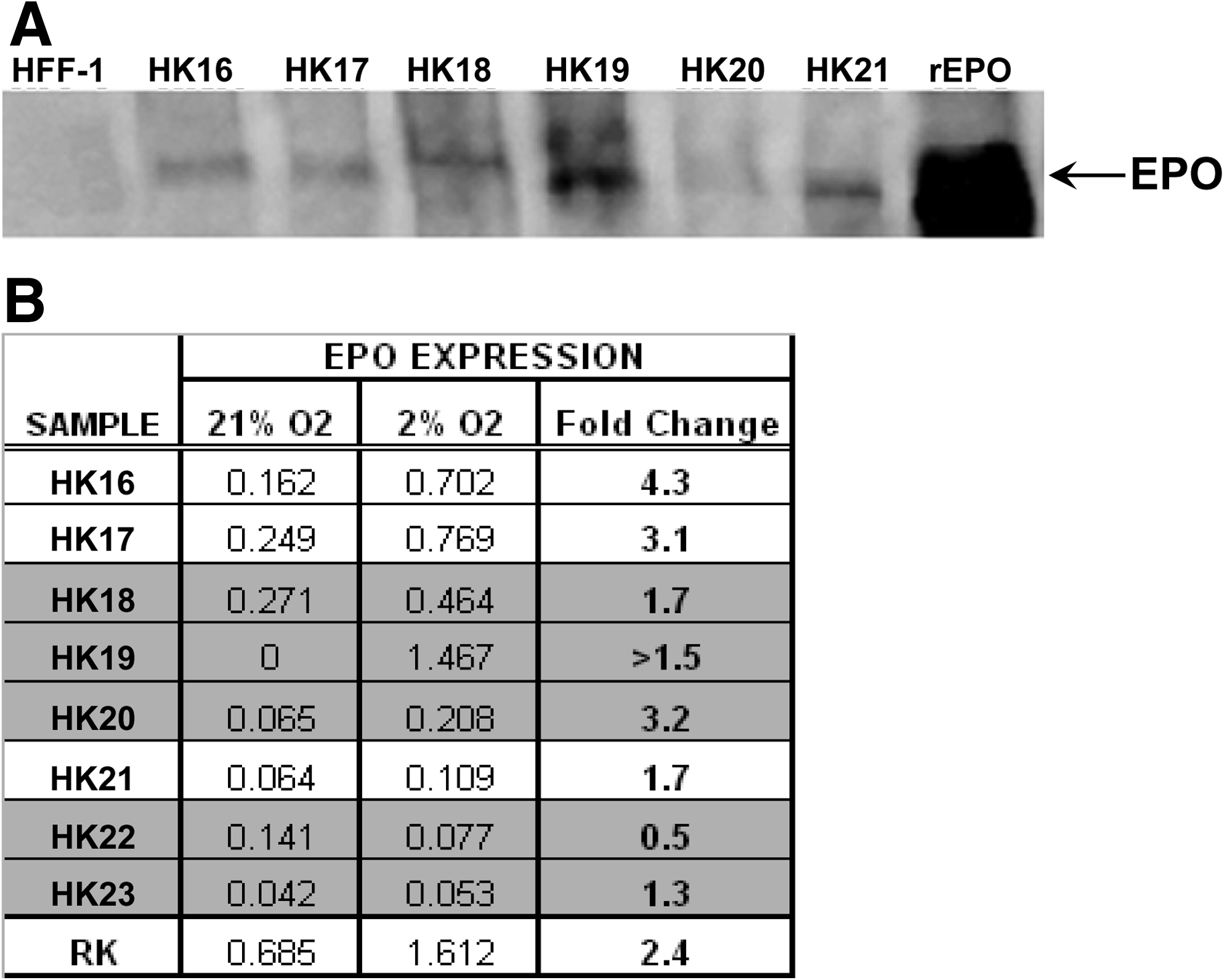
Initial cell yields per gram of tissue were lower from CKD (Table 4), which was anticipated from ZSF1 results (Fig. 2B). Without exception, application of the ICE methods developed for normal human tissue (Table 3) to human CKD tissues (5/5) yielded UNFX cultures that could be passaged, cryopreserved, and used to generate the B2 subpopulation (Table 4). To evaluate the functional status of UNFX isolates from human CKD tissues, oxygen regulation of EPO expression and receptor-mediated albumin uptake was assayed because previous studies of rodent UNFX demonstrated that oxygen-regulated EPO-producing endocrine cells and albumin-transporting tubular cells were retained in UNFX.9,26 EPO protein was assayed by western blot analysis and EPO levels in CKD-derived kidney tissues were comparable or greater than in non-CKD tissues (Fig. 4A), indicating that EPO-producing activity was intact in the CKD kidneys despite systemic anemia. To assess the continued expression and oxygen-responsiveness of EPO, one group of human UNFX cultures was subjected to a hypoxic stimulus and the levels EPO mRNA in 21% and 2% O2 were quantified by RT-PCR. EPO mRNA upregulation in response to hypoxic stimulus was retained in UNFX cultures established from 4/5 CKD and 3/3 non-CKD kidneys (Fig. 4B). In the case of HK022, EPO expression was present in the UNFX culture without hypoxia-induced upregulation. UNFX cultures from HK019 failed to express detectable EPO mRNA at 21% O2, but were significantly induced at 2% oxygen. Taken together, these data suggest that cells capable of producing EPO are retained in the kidney, even in advanced CKD, and these cells can be isolated and propagated in vitro while maintaining oxygen-regulated EPO-producing function.
In UNFX cultures from all species, epithelial cells are a predominant phenotype. Functionality of cultured renal tubular cells has been examined historically by a variety of assays, including albumin uptake 20 and the detection of hydrolase activities characteristic of the apical brush border membrane of the renal proximal tubule such as GGT and LAP.31,32 These assays were adapted to human UNFX to enable quantitation of albumin uptake and measurement of GGT and LAP activity and evaluate tubular cell function in UNFX cultures derived from CKD and non-CKD tissue. Regardless of disease state, UNFX cultures retained the functional capacity for protein transport (Fig. 5A–C) and key enzymatic activities involved with proximal tubular cell-specific metabolism (Fig. 5D). Performance of CKD-derived cells was equivalent or superior to non-CKD-derived cells in all three functional assays.

Functional attributes of human UNFX cultures (passage 1) derived from non-CKD (HK21) and CKD (HK22) kidneys.
Generation of B2 from biopsy-initiated human UNFX cultures
Clinical scenarios for using an autologous/homologous cell sourcing strategy for CKD therapies have indicated that renal biopsy would be a likely procedure for tissue procurement; therefore, we explored the feasibility of establishing UNFX cultures and B2 subpopulations from tissue harvested with a hollow core device used in clinical practice to collect diagnostic/prognostic biopsies from CKD patients. Population growth of UNFX was examined for 2–4 passages in cultures established from biopsies collected from non-CKD (HK021) or CKD (HK022 and HK023) samples and expansion rates and postculture yields were comparable (Fig. 6A). Because each biopsy of the kidney was taken with a device that permits observation of only the kidney capsule/surface before collection, it was not possible to select areas with healthy tubules or glomeruli; rather, core tissue samples spanning cortex through medulla were collected randomly from different regions of the kidney. Thus, it was of interest to know how reproducible the cellular composition was among biopsies generated from a single kidney.
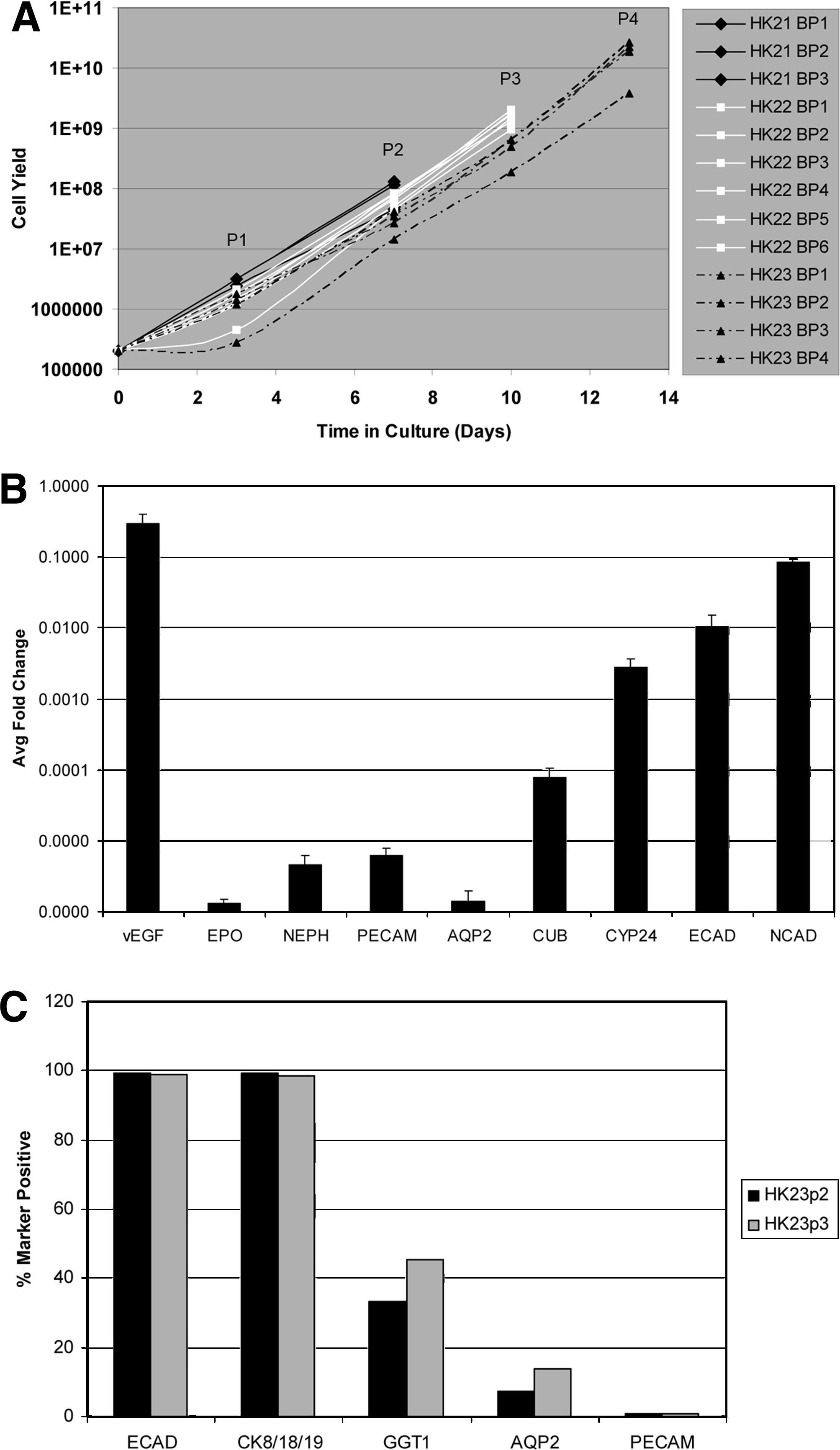
Reproducibility of core needle biopsy punch sourcing of UNFX and B2 from non-CKD (HK21) and CKD (HK22 and HK23) kidneys. UNFX cultures were harvested and subcultured (passaged) 2–4 times and quantified with each passage.
Six biopsies were obtained from HK022–a CKD kidney–and each biopsy was expanded as an independent UNFX culture. At passage 2, B2 subpopulations were isolated independently from each HK022 UNFX culture and characterized by gene and protein expression profiling to examine reproducibility of the isolation process. Gene expression of the previously identified markers and additional markers were measured in HK022 B2 isolates by quantitative RT-PCR relative to an endogenous control marker (PPIA). Genes associated with proximal and distal tubular cells were expressed abundantly in 6/6 of the HK022/B2 replicates with very low variability, indicating the composition of B2 is highly reproducible (Fig. 6B).
Additional studies were conducted to examine reproducibility of the B2 phenotype after serial passage of UNFX. B2 was generated from HK023 UNFX cultures after passage 2 and again after passage 3 and the B2 five-marker protein expression profile was quantitated by flow cytometry (Fig. 6C). As expected, the phenotype of the HK023 B2 subpopulations were >95% epithelial (ECAD+ and CK8/18/19+) with a significant proportion of GGT1+ tubular cells and a detectable level of AQP2+ ductal cells. Trace quantities of PECAM+ vascular cells were present as well, but at levels <1%. Interestingly, the percentage of GGT1+ and AQP2+ cells increased from passage 2 to passage 3, suggesting that key epithelial cell types characteristic of B2 are retained during UNFX subculture.
Discussion
Currently, a defined regulatory approval pathway exists to move regenerative medicine therapies that incorporate cells sourced via an autologous/homologous strategy into clinics.6–8 Although numerous practical challenges are inherent to developing and delivering therapies that incorporate autologous cells, autologous approaches remain attractive clinically because they avoid the need for immune suppression and may circumvent some concerns regarding pathogens. However, autologous/homologous sourcing strategies may prove infeasible when cell(s) of therapeutic interest are not procurable from the organ of interest in the target patient population due to disease-related destruction (e.g., insulin-producing beta cells in type 1 diabetes mellitus) or because of clinical risk associated with direct sourcing. Such limitations have led some to procure therapeutically relevant cells from an ectopic site, also known as an “autologous/heterologous cell sourcing strategy,” 6 or to employ stem or multilineage cells. When the therapeutically relevant cells are highly specialized, tissue-specific cells, these alternative approaches may face additional development challenges (e.g., difficulty scaling-up due to inefficiencies of differentiation protocols, and need to assess long-term stability of differentiated phenotype). 33 Studies in both the liver and kidney have indicated that resident parenchymal cells are more efficient than ectopically derived stem and progenitor cells at restoring function to damaged tissue after injury.15,34,35 These factors guided our decision to evaluate the feasibility of ICE defined renal subpopulations from CKD-relevant animal models and from diseased kidney tissue in humans.
Methods for isolating UNFX and the subpopulations obtained from UNFX by density gradient centrifugation were translated to two CKD-relevant animal models: ZSF1(fa/facp) rat and dog. The ZSF1(fa/facp) rat provides a model of renal failure secondary to obesity and insulin-resistant diabetes, and thus potentially represents a major portion of the CKD population worldwide.1,36,37 The obese ZSF1 model is characterized by age-dependent progressive nephropathy, 29 with systemically measurable renal degeneration (e.g., elevated BUN, sCREAT, and proteinuria). In a veterinary setting, aged dogs often present with spontaneously occurring chronic renal failure, the symptoms and progressive nature of which mimic key elements of CKD in humans. 38 Consequently, the dog is frequently selected as a preclinical model of kidney disease in the laboratory setting, where surgery, ischemia, and/or chemical treatment are used to achieve reduction in functional nephron mass.39–41
Demonstrating feasibility for autologous cell sourcing in disease-relevant animal models is necessary, but not sufficient, for translating product development into clinics; therefore, ICE methods were also evaluated with healthy and diseased human kidney tissues. In addition, the sampling methods used to obtain tissue from diseased kidneys (e.g., needle biopsy) replicated tissue sampling practices used in the clinic to obtain diagnostic biopsies. 42 The preliminary data obtained in this study on the isolation, characterization, and expansion of UNFX and UNFX-derived subpopulations will be valuable for guiding the design of future experiments as more information becomes available regarding the number and type(s) of cells needed to achieve a therapeutically relevant outcome.
The presence and function of specific kidney cell types (e.g., tubular cells and EPO-producing cells) were investigated in UNFX and B2 cultures established and propagated from rodents, dogs, and humans, including rodents and humans with confirmed CKD. Despite lower initial cell yields from CKD versus non-CKD kidney tissues, growth rates and population doubling times were comparable (Table 4 and Fig. 6). This study demonstrated that tubular cells are major components of UNFX cultures across species and disease states and are further enriched in the B2 subpopulation generated by density gradient centrifugation. In addition to the tubular phenotypic reproducibility measured by gene expression and flow cytometry, key tubule-associated functions such as receptor-mediated albumin transport and hydrolase activities involved in metabolism (GGT and LAP) were retained in B2. Functional attributes of the tubular cells present in CKD-derived cultures were comparable or superior to non-CKD-derived cultures (summarized in Table 4), despite significant systemic and histologic disease of the kidneys at the time of tissue procurement, suggesting that cells with therapeutic potential can be sourced by an autologous/homologous strategy from severely diseased tissue, even though such functions are compromised in situ. The ICE methods evaluated in this study reproducibly generated B2, a renal cell subpopulation enriched in functional tubular cell components and relatively depleted of other cell types, including large collecting duct cells, glomerular cells, endocrine cells, and vascular cells. Such phenotypic and functional reproducibility is compatible with the requirements of clinical manufacturing.
The EPO-producing cells of the kidney have been isolated and described as components of heterogeneous primary renal cell cultures.9,26 In the present study, EPO-producing cells were identified in UNFX cultures derived from rodent, dog, and human. EPO-producing cells were also present in UNFX cultures isolated from CKD tissue. In all cases tested, EPO expression was oxygen regulated; exposure of UNFX cultures to low (2%) oxygen levels increased EPO expression significantly compared to atmospheric (21%) oxygen. Multiple mechanisms have been hypothesized as explanations for anemia secondary to CKD; however, there is general agreement that EPO deficiency plays a role, as serum EPO levels are decreased in human CKD patients in proportion to the degree of anemia. 43 Some have postulated that EPO-producing cells are lost from the kidney as CKD progresses; our data provide evidence against a complete loss of EPO-producing cell mass, as four of five CKD kidneys and the UNFX cultures derived from these organs contained EPO-producing cells. During density gradient centrifugation of UNFX isolates, EPO-producing cells, along with glomerular and vascular cells become enriched in the B4 fraction in all species (Fig. 2B). Given the potential therapeutic value of oxygen-regulated EPO-producing cells, 44 it may be beneficial to investigate the therapeutic benefit of administering B4 alone or in controlled admixtures with other renal subpopulations in future in vivo experiments.
Of importance to tissue engineering is the expansion capacity of the input cultures from which therapeutically relevant cells are produced. 45 Ongoing studies are investigating the therapeutic potential of UNFX and the cell subpopulations described in this report; however, the focus of these studies was to investigate the feasibility of generating the cellular component of an autologous treatment for CKD from animal models relevant for product prototype evaluation and from diseased human kidneys. The initial yields of viable cells from tissues are presented for all species studied and data on the expansion capacity of human UNFX cultures derived from non-CKD and CKD tissues are presented in Figure 6. Although evaluations of optimal and maximal expansion parameters must await the identification and characterization of therapeutically relevant cell populations and dose ranges, Figure 6 illustrates consistency in UNFX expansion and presents growth rate data that is relevant for a preliminary assessment of feasibility. For example, the growth rates of four replicate UNFX cultures derived from HK23 (obtained from a CKD patient with serum creatinine of 6.4) did not yet exhibit a plateau at passage 4 (P4) and the cell yield of three out of four replicate cultures was >1 × 1010 cells. If a therapeutically relevant subpopulation was present at only 1% of UNFX, this yield would still represent 1 × 108, or 100 million cells. Such a yield, obtained without optimization of cell culture conditions, falls within a dose range that elicited de novo regeneration of urinary tissue in animals.46,47 B2, the tubular-enriched UNFX subpopulation B2, which previous studies suggest might have therapeutic potential based on its tubular-enriched composition,12–16 represents on average 50% of UNFX. Again, the yield of B2 that could be isolated from a UNFX culture derived from a biopsy sample of CKD tissue at <14 days in culture (HK23 at P4) is >5 × 109 B2 cells, a yield that falls within or exceeds dose ranges that elicited de novo urinary tissue regeneration in animals.46,47
In summary, these studies demonstrate that primary cultures containing cells from all major compartments of the kidney (UNFX) and two subpopulations whose compositions are each suggestive of potential therapeutic value12–16,44—a tubular cell-enriched subpopulation (B2) and a subpopulation containing oxygen-responsive Epo-producing cells (B4)—can be established reproducibly from healthy and diseased kidney tissue across multiple species. The successful adaptation of ICE methods to large animal and human CKD specimens highlights the feasibility of pursuing an autologous/homologous cell sourcing strategy for obtaining the cellular component of candidate regenerative therapies for CKD and translating such candidate therapies from preclinical to clinical evaluation. Despite advanced fibrosis and complex underlying metabolic disease, pockets of tubular, endocrine, collecting duct, and glomerular cells persist within CKD-derived tissue, providing access to the building blocks that may be required components of an autologous regenerative medicine therapy with the potential to preserve renal function and extend lifespan in many patients with CKD.
Footnotes
Disclosure Statement
All authors are employees of Tengion or were employees when experiments were performed.
Portions of this data appeared in an abstract published from the Experimental Biology 2009 meeting.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.