
Abstract
Stem cell-based cardiac tissue engineering is a promising approach for regenerative therapy of the injured heart. At present, the small number of stem cell-derived cardiomyocytes that can be obtained using current culture and enrichment techniques represents one of the key limitations for the development of functional bioartificial cardiac tissue (BCT). We have addressed this problem by construction of a novel bioreactor with functional features of larger systems that enables the generation and in situ monitoring of miniaturized BCTs. BCTs were generated from rat cardiomyocytes to demonstrate advantages and usefulness of the bioreactor. Tissues showed spontaneous, synchronized contractions with cell orientation along the axis of strain. Cyclic stretch induced cardiomyocyte hypertrophy, demonstrated by a shift of myosin heavy chain expression from the alpha to beta isoform, together with elevated levels of atrial natriuretic factor. Stretch led to a moderate increase in systolic force (1.42 ± 0.09 mN vs. 0.96 ± 0.09 mN in controls), with significantly higher forces observed after β-adrenergic stimulation with noradrenalin (2.54 ± 0.11 mN). Combined mechanical and β-adrenergic stimulation had no synergistic effect. This study demonstrates for the first time that mechanical stimulation and direct real-time contraction force measurement can be combined into a single multimodal bioreactor system, including electrical stimulation of excitable tissue, perfusion of the culture chamber, and the possibility of (fluorescence) microscopic assessment during continuous cultivation. Thus, this bioreactor represents a valuable tool for monitoring tissue development and, ultimately, the optimization of stem cell-based tissue replacement strategies in regenerative medicine.
Introduction
In cardiac tissue engineering, scaffold materials have two major functions: providing a physiologic extracellular environment and stabilizing the constructs generated in vitro.2,3 One beneficial effect of artificial patch transplantation is the mechanical stabilization of injured tissue, thus preventing further dilation of ventricles and the risk of congestive heart failure. 4
However, and in addition, the optimal tissue replacement construct should be able to exert contractile forces comparable to that of native cardiac tissue. With this in mind, the ideal cell source for cardiac tissue engineering would be patient-specific stem cell-derived cardiomyocytes. Cardiac differentiation has been demonstrated successfully for both embryonic stem cells5,6 and induced pluripotent stem cells7–10 from various species; however, these cells are currently not suitable for clinical application in patients due to the methods used in their generation. Despite recent promising results regarding scalable expansion11,12 and differentiation 13 of embryonic and pluripotent stem cells, the available numbers of stem cell-derived cardiac cells have not been sufficient for the generation of BCT of appropriate sizes for therapeutic application so far. Therefore, the aim of this study was to generate a bioreactor for the development and testing of miniaturized bioartificial tissue, enabling small-scale stem cell-based cardiac tissue engineering.
It has been proposed that mechanical stretch can improve tissue formation and contractility of bioartificial cardiac grafts generated from primary chicken cardiomyocytes 14 or cardiomyocytes of neonatal and fetal rodent origin.15,16 This has also been demonstrated for human heart cells seeded on gelatin scaffolds. 17 Probable underlying mechanisms include a hypertrophic response of myocardium to a prolonged increase in cardiac workload (reviewed in ref. 18 ). Analysis of the gene expression profile of neonatal ventricular rat cardiomyocyte cell cultures exposed to mechanical stretch in vitro showed increased expression of a number of genes, for example, heat shock protein 70, growth differentiation factor 15, and metallothionein-1. 19 Interestingly, a robust change in expression levels of these genes was observed specifically after stretch-induced hypertrophy, whereas pharmacologically induced hypertrophy by treatment with phenylephrine yielded only moderate effects. 19 Downstream targets of stretch-induced genes included several proteins of the cardiomyocyte contractile apparatus, as well as secreted proteins.20,21
In the past, bioreactors were used to improve the mechanical properties of three-dimensional tissues generated in vitro from cardiomyocytes. However, it has been difficult to directly monitor the effects of mechanical stretch and other stimulants continuously during the course of tissue formation. Usually, measurements require transfer of the generated tissue from a cultivation vessel to a measurement device or organ bath chamber14,15,22; therefore, force measurement has been predominantly used as an end-point analysis. With this approach, long-term data acquisition can only be managed with numerous replicate samples requiring a multiple of the cell numbers that would be needed in continuous analyses. Alternatively, indirect measurement of contraction forces via optical analysis has been proposed for noninvasive on-line monitoring during prolonged culture. 23
Herein we describe the design of a novel multimodal bioreactor for mechanical stimulation and real-time direct measurement of contraction forces under continuous sterile culture conditions. The bioreactor's transparent cultivation chamber allows for microscopic assessment of tissue development including fluorescence imaging. Additional functions include electric pacing of tissues, as well as the possibility to perfuse the central cultivation chamber, allowing for continuous medium exchange and/or controlled addition of pharmacologically active agents. In this study, our novel multimodal bioreactor was used to monitor the development of contractile properties during tissue maturation, investigating the effect of mechanical stretch in comparison to β-adrenergic stimulation with noradrenalin.
Material and Methods
Cell isolation
Neonatal rat cardiomyocytes were isolated from 1- to 3-day-old Sprague-Dawley rat pups as described previously 24 and enriched by discontinuous Percoll (GE Health Care) gradient centrifugation. During all experiments, the Principles of Laboratory Animal Care (NIH publication No. 86-23, revised 1985) as well as the Animal Welfare Law of Lower Saxony were followed.
Preparation of BCTs
A bipartite Teflon mold with a volume of 200 μL was designed for BCT casting. Between the lower and the upper part, two titanium rods are held in position (distance 6 mm) and can be cast integrally with a liquid cell/matrix mixture. In brief, 106 freshly isolated cardiomyocytes per construct were mixed with a liquid matrix based on rat tail collagen type I (R&D Systems) and Matrigel (BD Biosciences) as described by Zimmermann et al.
15
BCTs were cast in the Teflon molds placed on agarose coated dishes; consequently, the bottomless molds allow for microscopic assessment of early tissue formation. After solidification, the tissue is suspended between the rods (Fig. 1A) and can be transferred to the cultivation chamber of the bioreactor. A detailed description of matrix and medium composition can be found in the online Supplementary Data (available online at

Multimodal modular bioreactor system and experimental design. For bioartificial cardiac tissue (BCT) casting, a bipartite Teflon mold (
Multimodal bioreactor system
A modular expandable bioreactor was designed to apply cyclic stretch to BCTs; each module connects a cultivation chamber to both a linear motor with integrated position measurement, and a force sensor (Fig. 1B, C). The bioreactor modules are placed in a standard cell culture incubator at 37°C, 5% CO2, and 80% humidity (Fig. 1D). The cultivation chamber with a working volume of 1–8 mL was made of sterilizable polysulfone thermoplastic resin combined with a glass bottom for microscopic assessment. Two opposite mounting brackets hold the titanium rods and allow connection of the BCT to the linear motor on one side and the force sensor on the other via two opposing feedthroughs sealed with 0.2 mm silicon membranes to prevent medium leakage. Two lateral connections can be used for medium perfusion through the cultivation chamber; two fittings for electrodes integrated into the chamber lid allow for indirect electrical stimulation of the tissue embedded in conductive medium or Tyrode's solution (Fig. 1C). A stimulation current generator was developed for generating stimulation current pulses from 0 to 25 V with alternating polarity to prevent hydrolysis. Pulse width is software controlled ranging from 5 ms to 10 s. The linear motor consists of a PS01-23Sx80 stator and a PL01-12 × 130/80 slider and has a maximum stroke of 30 mm with a peak force of 44 N and a position repeatability of 0.01 mm, driven by an E400-AT servo controller (all from LinMot). The displacement is measured with an SM277 inductive displacement transducer (Schreiber Messtechnik). Forces are measured using a miniature bending beam load cell (Burster Praezisionsmesstechnik) with a measuring range of 0–1 N.
In this setting, the cultivation chambers can be easily detached from the bioreactor without leakage; therefore, they are fully interchangeable between individual bioreactor stands and can be placed on a microscope stage for imaging. Alternatively, for continuous assessment within the bioreactor an integrated optical imaging system could be included.
Software control
Controllers are connected to a PC via USB; software (written in LabVIEW 8.6 for Windows XP) was developed for controlling motors and sensors of up to four bioreactor modules in parallel. Force sensors were calibrated with class F1 calibration weights (KERN & Sohn). Constant data acquisition and storage can be performed; all parameters of linear displacement (zero position, stroke, frequency, and wave shape) and electric stimulation (frequency, amplitude, and pulse width) can be adjusted while operations are running. Force and displacement data are displayed on-line in real-time diagrams and captured as CVS files (comma-separated values), which can be exported to common spreadsheet programs, e.g. MS Excel. For an improved signal-to-noise ratio of force data, an averager integrating multiple signals can be activated.
Experimental design
The experimental design consisted of cyclic strain stimulation experiments with BCTs and corresponding static control experiments under varying culture conditions (Fig. 1E). Each experiment started with the preparation of BCTs from freshly isolated neonatal rat cardiomyocytes cast in Teflon molds. As noradrenalin (in ascorbic acid solution) has been described to support cardiomyocyte contractility in cell culture, 25 β-adrenergic stimulation was started on day 6 (d6) of culture by addition of 10 μM noradrenalin and 30 μM ascorbic acid (both from Sigma-Aldrich). After 7 days of cultivation, BCTs were transferred to the bioreactor. Mechanical stimulation was started on d7 of culture using 10% cyclic longitudinal strain (0.6 mm, sinus shaped) with a frequency of 1 Hz. Stretch stimulation was continuously applied for 7 days, force measurement was performed daily from d7 until d14. During mechanical stimulation, the connection of the BCT to the force sensor was fixed. The perfusion system was not active during these experiments; cell culture medium was exchanged manually every day.
Imaging
For assessment of cellular morphology during tissue formation, continuous live cell imaging of BCTs was performed with 25 nM tetramethylrhodamine methyl ester (TMRM; Invitrogen). TMRM is a nontoxic cationic, mitochondrial selective probe, which can be used to selectively stain cells with high metabolic activity such as cardiomyocytes. 26 Fluorescence images and time-lapse videos of BCTs were captured from d0 to d6 (in Teflon molds) and from d7 to d14 (in cultivation chambers detached from the bioreactor) using the AxioObserver Z1 fluorescence microscope with Axiovision software 4.70 (Zeiss). Three-dimensional analyses for volumetric cellular distribution were performed on d7, whereby BCTs were fixed and used for whole-mount 4′,6-diamidino-2-phenylindole (DAPI) staining of nuclei. Reconstructed images were generated from data acquired with a custom-made optical projection tomography setup. The resolution of the reconstructed data was calculated to be 5 μm. Fixed BCTs were kept in propylene glycol for index matching to decrease tissue scattering properties. DAPI was excited using a 355 nm microchip laser (Soliton). Fluorescence was captured using a 465 ± 15 nm band pass filter (Semrock) with a CCD LUCA R (Andor).
Force measurement
Long-term spontaneous systolic contraction force was monitored daily for each tissue at 37°C, 5% CO2 in an incubator, 30 min after medium exchange. Spontaneous active contraction forces were detected with no preload. On d14, maximum forces were captured upon electric stimulation of the tissues at different preloads. For these end-point analyses, all tissues were washed three times with Tyrode's solution (140 mM CaCl2, 5.4 mM KCl, 1.8 mM CaCl2, 10 mM Glucose, and 10 mM HEPES) and allowed to equilibrate in Tyrode's solution for 30 min before force measurement. With preload increasing by 0.1 mm increments, tissues were paced five times at each step (rectangular, 5 ms at 25 V).
Measured forces are given as absolute values in mN per BCT (mean ± SEM for each group), thus representing forces normalized to equal starting quantities of cell number and matrix components.
Histological evaluation
For histological evaluation, BCTs were frozen, sectioned, and immunostained. Detailed procedures as well as specifications on the antibodies used can be found in the online Supplementary Data.
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA was prepared from BCTs and used for cDNA synthesis. Real-time-polymerase chain reaction (PCR) was performed to assess expression levels of the cardiac myosin heavy chain isoforms alpha and beta (α-MHC, β-MHC), atrial natriuretic factor (ANF), connexin 43 (Cx43), as well as calsequestrin-2, all relative to the reference gene β-actin. Detailed procedures and specifications on the primers used can be found in the online Supplementary Data.
Statistical analysis
Statistical analysis was performed with GraphPad Prism software (version 5.03 for Windows; GraphPad Software). Values reported are means and standard errors of the mean. Unless stated otherwise, data were analyzed by one-way analysis of variance, with the Bonferroni multiple comparison test for comparison of any two groups, probability values <0.05 were considered significant.
Results
Generation of BCTs
After seeding neonatal rat cardiomyocytes with a liquid matrix mixture into Teflon molds, BCTs solidified within 30 min. Tissues formed by shrinkage and detachment from the mold resulted in a biconcave tissue band suspended between the two titanium rods (Fig. 2A). During the initial cultivation period, shrinkage led to a decrease of BCT cross-sectional area from 20 mm2 at seeding to 1.26 ± 0.04 mm2 on d6 (data not shown). Further compaction of cells and matrix was observed for all BCT groups treated in our novel bioreactor. Compared to the untreated control, shrinkage was less pronounced in BCTs treated with stretch alone, whereas both β-adrenergic stimulation alone and combined mechanical and β-adrenergic stimulation led to a further reduction of the cross-sectional area to a minimum of 0.60 ± 0.01 mm2 on d14 (Fig. 2B).
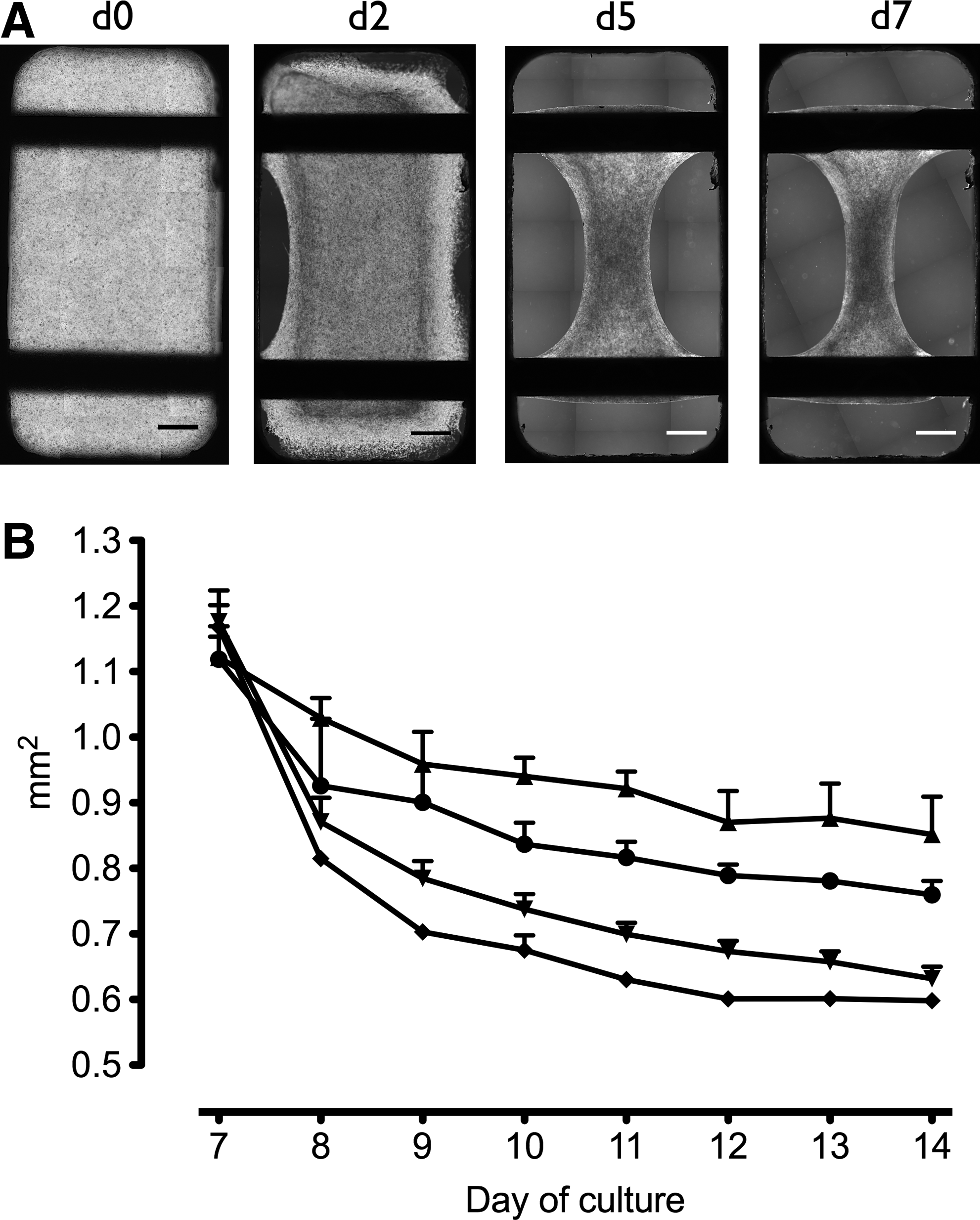
Microscopic assessment of early tissue formation in BCTs on day 0 (d0), d2, d5, and d7 (from left to right) shows shrinkage and detachment from the Teflon mold resulting in a biconcave tissue band suspended between the titanium rods (
Live cell fluorescence imaging after TMRM staining confirmed the presence of viable and metabolically active cardiomyocytes within the BCTs and demonstrated that tissue formation is associated with cardiomyocyte elongation and cellular alignment along the longitudinal axis of the constructs (Fig. 3A and Supplementary Video S1). Spontaneous contractions of single cardiomyocytes started from d2, and further cultivation led to synchronous beating of whole BCTs (Supplementary Fig. S1).

Live cell fluorescence images of mitochondria-rich cardiomyocytes labeled selectively with tetramethylrhodamine methyl ester on d0, d4, d8, and d12 (top down) demonstrate cell elongation and alignment along the longitudinal axis of the BCTs during the course of tissue formation and maturation (
Three-dimensional analysis of whole-mount DAPI-stained BCTs using optical projection tomography showed cell distribution throughout the whole tissue on d7, although at lower numbers in the center of the tissue (Fig. 3B and Supplementary Video S2). BCTs could be maintained in culture for up to 6 weeks, with sustained spontaneous contraction activity (data not shown).
Cardiac-specific gene expression
To investigate whether mechanical stimulation or β-adrenergic stimulation of BCTs in our novel bioreactor could elicit a cellular response, differential gene expression of BCTs was analyzed by quantitative real-time PCR and compared to untreated controls (normalized to equal 1.00). We investigated several genes that have been associated with a hypertrophic response.20,27 Indeed, on d8 of cultivation (after 24 h of mechanical stimulation) the ratio of β-MHC to α-MHC expression was higher in all treated groups compared to the untreated controls (Fig. 4A). Similar ratios were detected for stretched (1.64 ± 0.04) and β-adrenergic-stimulated BCTs (1.51 ± 0.06); application of both β-adrenergic stimulation and stretch resulted in a further increase of β-MHC to α-MHC expression level ratio to 2.25 ± 0.13 (all p < 0.05 vs. control). Similarly, mechanical stimulation led to elevated ANF expression levels (2.34 ± 1.46; p > 0.05) (Fig. 4B). Compared to the untreated control group, significantly higher ANF expression levels were detected in the two groups treated with β-adrenergic stimulation, either without additional stretch (6.95 ± 1.14) or with stretch (6.17 ± 1.10; both p < 0.05); in this case, stretch did not lead to a further increase. Notably, on d14 (after 7 days of mechanical stimulation), treated BCTs showed indications of a sustained and intensified hypertrophic response. Here, BCTs stimulated both β-adrenergic with noradrenalin and with stretch showed the highest increase in β-MHC to α-MHC expression level ratio of 3.85 ± 1.44 (Fig. 4A; p < 0.05 vs. control) together with an increase in ANF expression level of 17.94 ± 7.59 (Fig. 4B; p > 0.05 vs. control).

Expression levels of cardiac marker genes were analyzed by quantitative real-time polymerase chain reaction on d8 and d14 of cultivation for individual groups treated with different stimuli: none (negative control), stretch, β-adrenergic stimulation, and β-adrenergic stimulation + stretch. Depicted is the ratio of β-MHC to α-MHC expression (
Tissue morphology and composition
BCT morphology on d14 was assessed by costaining of tissue sections using cell type-specific antibodies. Viable cardiomyocytes were detected in all constructs and usually showed cellular alignment along the longitudinal axis of the tissue (Fig. 5, left panels). Cardiomyocytes were surrounded by nonmyocytes, with lower numbers of cardiomyocytes in the center of the tissue, as seen in the optical projection tomography analysis. Bundles of cardiomyocytes were notably thicker in the treatment groups and were occasionally found deeper in the tissue's center compared to the nontreated tissues. Cardiomyocytes showed well-developed sarcomeres in all groups and the presence of Cx43 indicated interconnections through gap junctions similar to neonatal rat heart (Fig. 5, middle and right panels). Interestingly, noncardiomyocytes expressing Cx43 were found at the rim of the neonatal rat heart, as well as at the borders of BCTs. It has been suggested that overall cardiomyocyte content or efficiency of coupling via gap junctions in aligned cells might play a role in force development.28–30 Using quantitative real-time PCR, total expression levels of Cx43 on d14 of tissue formation were found to be slightly elevated in both groups treated with mechanical stimulation (either with or without β-adrenergic stimulation); however, the effect did not reach statistical significance (Supplementary Fig. S3A). Also, the cardiomyocyte content of BCTs assessed as calsequestrin-2 expression levels did not differ significantly among groups; here, a minor increase was observed after β-adrenergic stimulation (Supplementary Fig. S3B). The reduced shrinkage of BCTs treated with stretch only, resulting in the largest cross-sectional area on d14 (Fig. 2B), correlated with an increased protein-to-DNA ratio in this group (Supplementary Fig. S3C).

Representative fluorescence microscopy images of immunostained tissue sections of different BCT groups: (
Mechanical properties
Long-term observation of spontaneous systolic forces with our bioreactor confirmed synchronous beating of BCTs. Recordings of contractions and a comparative analysis of beating frequencies over time are shown in the Supplementary Figure S1. While groups treated with β-adrenergic stimulation by noradrenalin initially showed an increased frequency of spontaneous contractions (d7 and d8), long-term exposure to noradrenalin resulted in beating frequencies similar to the other groups from d9 to d14. For tissues kept in the cell culture medium, systolic force was measured daily and found to increase over time for all groups (Fig. 6A). Compared to the untreated control group, significantly higher forces were observed for the two groups treated with β-adrenergic stimulation: without additional stretch already on d8, with stretch on d9 (two-way analysis of variance, p < 0.05). From d13 onward, BCTs treated with β-adrenergic stimulation alone yielded significantly higher systolic forces than BCTs treated with β-adrenergic stimulation and stretch. Spontaneous systolic forces on d14 reached a maximum of 0.96 ± 0.09 mN for the untreated control group (Fig. 6B). While stretch alone resulted in a moderate increase to 1.42 ± 0.09 mN (p > 0.05), significantly higher contraction forces were measured on d14 after β-adrenergic stimulation (2.54 ± 0.11 mN) and β-adrenergic stimulation + stretch (2.09 ± 0.08 mN; both p < 0.05).
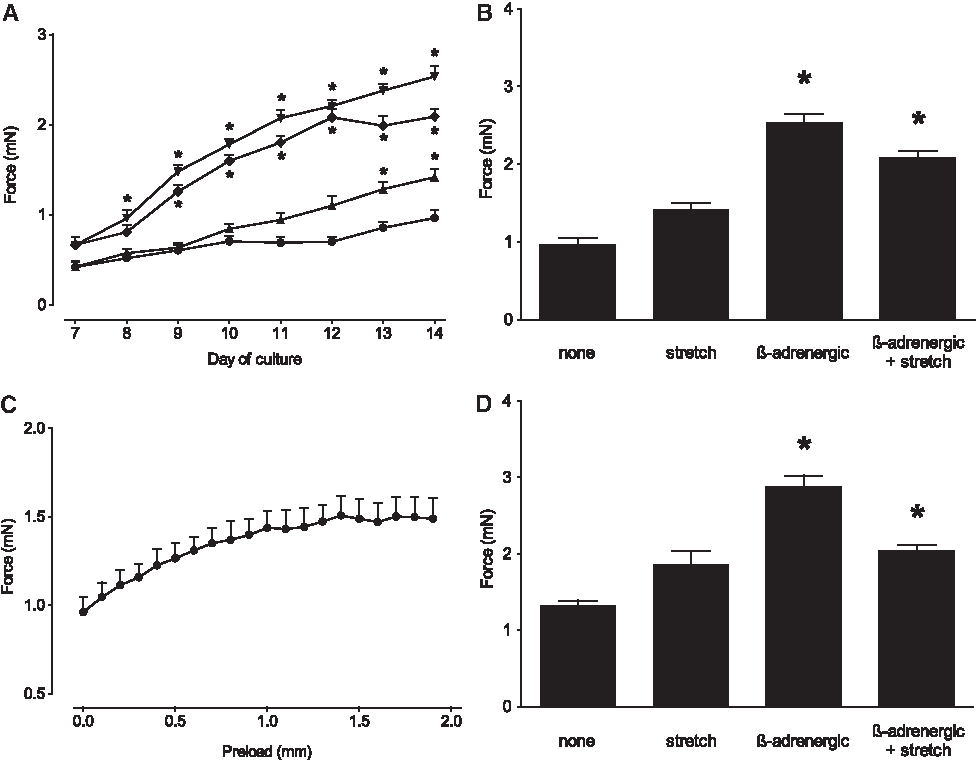
Systolic contraction forces of BCTs were measured in the bioreactor during continuous cultivation. For individual groups treated with different stimuli: •, none (negative control group; n = 11); ▴, stretch (n = 6); ▾, β-adrenergic stimulation (n = 17); ♦, β-adrenergic stimulation + stretch (n = 13); spontaneous systolic force development was assessed daily from d7 to d14 in the respective medium and analyzed using two-way analysis of variance (
Most often, and in contrast to the long-term observation performed in our study, end-point analysis of contraction forces is performed applying a defined preload to the tissue. Thus, for a comparative analysis, measurements were repeated with preloads between 0.0 and 1.9 mm upon electrical pacing of the tissue in Tyrode's solution without supplements. As expected for functional myocardial fibers, forces increased with higher preload due to the Frank-Starling mechanism (example given in Fig. 6C). For each group, the optimal preload length Lmax resulting in maximum forces was determined and used for end-point comparison of the four groups. While absolute forces tended to be higher when measured under these conditions, with a maximum systolic force of 2.88 ± 0.14 mN for the β-adrenergic stimulation group (Fig. 6D), differences between the analyzed groups were very similar to those observed for spontaneous contractions in the culture medium depicted in Figure 6B. With a BCT cross-sectional area of 0.63 ± 0.02 mm2, as determined by microscopic assessment, the strongest BCTs were those treated with β-adrenergic stimulation exerting a tensile strain of 4.57 ± 0.08 mN/mm2. Notably, electrical pacing did not have a significant effect on systolic forces compared to spontaneous contractions under similar conditions (data not shown).
In conclusion, long-term observation of spontaneous systolic forces in cell culture medium provides a good representation of maximum forces. Accordingly, for a valid comparison of systolic forces of different treatment groups, BCT transfer into a special organ bath is not required.
Discussion
Native heart tissue is characterized by a distinctive cycle of mechanical loading and unloading, which may be essential for successful cardiac tissue engineering.
The objective of this study was to investigate whether our novel multimodal bioreactor for mechanical stimulation, direct real-time force measurement, and imaging of small-scale tissue-engineered cardiac constructs could both influence and assess transient development of constructs over a 1-week period in a sterile culture environment. To investigate the influence of mechanical load on engineered cardiac tissue, motorized stretching devices for the application of defined strain have been described.14,15,31 Traditional mechanical testing devices to measure mechanical properties of tissue-engineered constructs typically require discontinuation of the cultivation process and therefore provide only end-point measurements of the respective samples. On the other hand, sophisticated bioreactor systems have been developed allowing for long-term monitoring of contractile activity via software-based analysis of video-optically captured contractions. 23
This study demonstrates for the first time that these features can be combined into a single multimodal bioreactor system, including the possibility for electric stimulation of excitable tissue, perfusion of the culture chamber, and microscopic assessment of tissue formation. The system was designed for small-scale cardiac tissue engineering, allowing the generation of tissue from small (stem) cell numbers, yet with a construct size large enough both to provide a true three-dimensional culture environment in vitro, and to be of an appropriate size for small animal transplantation models in vivo, without the immediate need for scale-up. If desired, scale-up can be easily achieved due to the flexibility of the modular bioreactor system. Current molds can be used for cell numbers ranging from 0.5 million to 4 million cells; other molds can be implemented for larger volumes and different geometries. A range of motors with different parameters (maximum stroke, peak force) are available as well as force sensors with a variety of measuring ranges. All components can be easily exchanged, which is a clear advantage over most existing systems.
Here, the novel system was used to provide a continuous monitoring of the developmental effects of mechanical stretch and β-adrenergic stimulation with noradrenalin on BCT. BCTs were generated using neonatal rat cardiomyocytes and displayed synchronized spontaneous contractions. Electrical stimulation in the bioreactor induced contractions of whole BCTs demonstrating the existence of a functional and excitable syncytium. However, it must be acknowledged that due to the spontaneous contractile activity, mechanical strain was present in all groups of BCTs to a certain degree. Thus, in our system, we were only able to investigate the effects of additional external mechanical loading.
Frank et al. demonstrated differential gene expression for cardiomyocytes stimulated by chemically induced contraction-related strain in comparison to mechanical stretch applied externally. 19 In line with their results, known stretch-responsive genes were differentially up- or downregulated in response to mechanical stimulation in our system investigating three-dimensional tissue-engineered constructs. It should be noted that Frank et al. used phenylephrine, a selective agonist of α-adrenergic receptors, for pharmacologic induction of hypertrophy. In comparison, noradrenaline used in our study is a β-adrenergic stimulant of cardiomyocyte contractility; however, it has also been described to induce hypertrophy in cardiomyocytes via an α-adrenergic response. 32 The expected hypertrophic response of cardiomyocytes to mechanical loading was confirmed by demonstrating a shift of cardiac MHC expression from the α-MHC isoform toward β-MHC. A reduction in α-MHC expression levels, reflecting changes in gene expression more toward a fetal phenotype, 20 was also described for three-dimensional cultures of murine embryonic stem cell-derived cardiomyocytes after application of 10% stretch at 1 Hz; however, higher frequencies led to the opposite effect. 31 At the same time, we were able to detect upregulation of ANF expression both after mechanical and β-adrenergic stimulation, as was described before for stretched monolayer 19 and three-dimensional cultures of rat cardiomyocytes, 14 as well as ventricles of the hypertrophic rat heart. 33 From the quantitative real-time PCR data and the force measurements, we conclude that the mechanical stimulation applied in our bioreactor led to moderate hypertrophy in cardiomyocytes within the BCTs, which had a positive effect on systolic forces. However, it has to be considered that in an in vivo situation, hypertrophy can reflect either a compensatory or maladaptive response to loading. 27 Therefore, it will have to be investigated whether longer periods of stimulation might have adverse effects on our BCTs.
While gene expression analysis clearly showed a hypertrophic response, we found that protein/DNA ratio was not a valid indicator of hypertrophy in our three-dimensional cultivation system, most likely due to the artificial cellular environment including substantial protein supplementation. Nevertheless, protein/DNA ratio showed good correlation with total BCT diameters.
Besides the direct effect of mechanical stretch on cell fate via stretch-responsive differential gene expression, stretch might also lead to enhanced perfusion of cell culture medium within the constructs. This could result in improved nutrition and oxygen supply together with enhanced removal of metabolic waste products and/or nonintegrated matrix components. As this bioreactor system allows for perfusion of the central culture vessel, it can be used for a comparative analysis of the effects of mechanical stimulation and continuous medium exchange at defined rates, or alternatively, to monitor pharmacological effects of soluble compounds on tissue contractility. Nutrition and oxygen supply are also major limiting factors for the generation of larger myocardial tissue constructs needed for future clinical application. To allow for perfusion, recent studies introduced blood vessels within bioengineered myocardial tissue, either by adding endothelial cells to generate prevascularized tissue in vitro 34 or by promoting neovascularization after transplantation in vivo. 35 These strategies might be used to generate larger tissue that more closely resembles native myocardium.
In line with previous reports, mechanical stimulation led to increased systolic forces compared to untreated control tissues in our experiments. After application of 5% unidirectional phasic stretch for 6 days, Fink et al. reported two- to fourfold higher force of contraction both under basal conditions and after stimulation with calcium or the β-adrenergic agonist isoprenaline. 14 Zimmermann et al. demonstrated in a direct comparison of calcium response curves that engineered heart tissue benefited from mechanical load in addition to oxygen supplementation. 36 Using a similar system based on collagen and Matrigel, Clause et al. generated engineered early embryonic cardiac tissue using embryonic cardiac cells from chicken. 37 They reported a 1.6-fold increase in active stress at Lmax following cyclic mechanical stretch stimulation for 48 h and concluded that contractile properties and cellular proliferative response to stretch mimic developing fetal myocardium.
A number of these studies used adrenoreceptor agonists; however, mostly as an analytical tool.2,14,15,38 These studies demonstrated an increase in contractility in response to short-term application of isoprenalin, indicating functional adrenergic signaling in engineered heart tissue. Long-term application was described for instance by Claycomb et al. in a protocol designed for the maintenance of contractility in cultured cardiac HL-1 cells including continuous medium supplementation with noradrenalin. 25
Using our bioreactor system we were now able to demonstrate for the first time in a direct comparison that long-term chemical stimulation by addition of noradrenalin and ascorbic acid yielded significantly higher contraction forces in BCT, that is, increased by 2.5-fold, than mechanical stretch alone (1.4-fold). Notably, addition of noradrenalin was associated with an elevated contraction frequency in the first 2 days, but upon prolonged exposure, frequency returned to similar values as in the other groups. Moreover, interestingly, no additive or synergistic effect on systolic forces was observed for combined mechanical and β-adrenergic stimulation. Several studies investigated the influence of stretch on Cx43 expression in cultures of neonatal cardiomyocytes and described upregulation after mechanical stimulation.29,30 In our setting, no clear correlation was observed for either cardiomyocyte content or Cx43 expression levels with force development. However, Salameh et al. also demonstrated that application of stretch for 48 h resulted in lower Cx43 expression than stretch for 24 h. 29 This might explain why no significant upregulation of Cx43 was observed in our system after prolonged mechanical stimulation for 7 days.
For our strongest BCTs, we determined a maximum tensile strain of 4.57 ± 0.08 mN/mm2, which is still well below the contraction force of ∼50 mN/mm2 exerted by the papillary muscle of native rat myocardial tissue.39,40 For engineered cardiac tissues, a direct comparison of absolute contractile forces among different studies is difficult due to variations of protocols and depends on a comparable assessment of tissue construct diameter. Frequently, authors chose to denote the forces measured per tissue construct. Although this does not consider changes in cell number over time through cell proliferation and death, we have back-calculated forces reported in individual studies to a starting amount of 106 cells used for tissue generation. Typically, studies using rat cardiomyocytes together with a collagen/Matrigel matrix reported forces between ∼0.3 mN and ∼1.5 mN/106 starting cells.14,15,22,23,36,38 Based on this calculation mode, the systolic forces of ∼1 mN for untreated control BCTs were in the range of previous studies, indicating the functionality of BCTs generated with our bioreactor system. The maximum systolic forces of 2.88 ± 0.14 mN obtained after β-adrenergic stimulation were almost twofold higher than achieved so far.
In conclusion, first results with the novel bioreactor system substantiate the potential of mechanical stimulation to influence tissue-engineered cardiac construct development. At the same time, they provide an indication on the timing of structural and mechanical changes that occur during this process. In our experimental setup using primary cardiomyocytes, β-adrenergic stimulation had a stronger effect on systolic force development than mechanical stimulation. Nevertheless, it is conceivable that mechanical stimulation might provide important signals for cardiac differentiation of stem cells within three-dimensional tissues. While optimal mechanical properties for in vivo support of cardiac function have to be determined in future studies, the novel bioreactor described herein can be considered a valuable tool for in vitro analysis of BCT and thus for optimization of stem cell-based tissue replacement strategies in regenerative medicine.
Footnotes
Acknowledgments
The authors are grateful to Suzanne E. Dorfman for revising the article. We would like to thank Ingrid Schmidt-Richter and Anke Gawol for providing excellent technical assistance as well as Michael Breyvogel, Jörg Claus, Juri Huber, and Erik Mahnke for valuable contributing to the bioreactor setup. The monoclonal antibody 9D10 developed by M.L. Greaser was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by Department of Biology, The University of Iowa, Iowa City, IA. This work has been supported by the German Federal Ministry of Education and Research (01GN0520) as well as by the Cluster of Excellence REBIRTH.
Disclosure Statement
No competing financial interest exists.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.