
Abstract
The developmental potential of human embryonic stem cells (hESCs) holds great promise to provide a source of human hepatocytes for use in drug discovery, toxicology, hepatitis research, and extracorporeal bioartificial liver support. There are, however, limitations to induce fully functional hepatocytes on conventional two-dimensional (2D) static culture. It had been shown that dynamic three-dimensional (3D) perfusion culture is superior to induce maturation in fetal hepatocytes and prolong hepatic functions of primary adult hepatocytes. We investigated the potential of using a four-compartment 3D perfusion culture to induce hepatic differentiation in hESC. Undifferentiated hESC were inoculated into hollow fiber-based 3D perfusion bioreactors with integral oxygenation. Hepatic differentiation was induced with a multistep growth factor cocktail protocol. Parallel controls were operated under equal perfusion conditions without the growth factor supplementations to allow for spontaneous differentiation, as well as in conventional 2D static conditions using growth factors. Metabolism, hepatocyte-specific gene expression, protein expression, and hepatic function were evaluated after 20 days. Significantly upregulated hepatic gene expression was observed in the hepatic differentiation 3D culture group. Ammonia metabolism activity and albumin production was observed in the 3D directed differentiation culture. Drug-induced cytochrome P450 gene expression was increased with rifampicin induction. Using flow cytometry analysis the mature hepatocyte marker asialoglycoprotein receptor was found on up to 30% of the cells in the 3D system with directed hepatic differentiation. Histological and immunohistochemical analysis revealed structural formation of hepatic and biliary marker-positive cells. In contrast to 2D culture, the 3D perfusion culture induced more functional maturation in hESC-derived hepatic cells. 3D perfusion bioreactor technologies may be useful for further studies on generating hESC-derived hepatic cells.
Introduction
One possible alternative cell source may be stem cell-derived human hepatocytes. 2 With their potential to differentiate into virtually any cell type and unlimited expansion capability, human embryonic stem cells (hESCs) might be the ideal source to provide human hepatocytes for use in drug discovery, toxicology studies, hepatitis virology, and bioartificial liver support systems. Numerous studies have been conducted to differentiate hESC into hepatic cells.3–9 These experiments, however, revealed that hESC-derived hepatocyte-like cells are more like immature fetal hepatocytes in the aspect of the gene expression pattern and function. While initial hESC studies are based on conventional two-dimensional (2D) static culture, it had been shown that even primary hepatocytes rapidly de-differentiate in conventional 2D cell culture systems, resulting in deterioration of liver-specific function.10–12 This problem can be ameliorated by using dynamic three-dimensional (3D) perfusion culture systems for directed differentiation to better mimic the in vivo environment for progenitor development in vitro. As various authors have described, 3D culture and physical parameters, such as flow, improve survival for hepatocytes in vitro.13–15 We have previously shown that hollow fiber based dynamic 3D perfusion culture systems prolong hepatic functions of primary adult hepatocytes.15,16 Moreover, the 3D culture system successfully enhanced hepatic maturation in fetal hepatocytes.14,17
To our knowledge, the influence of physical parameters on hepatic differentiation (HD) of hESC, such as perfusion, has not been investigated so far. We applied a novel approach to achieve enhanced HD of hESC by combining the use of cytokines with the culture in a dynamic 3D perfusion system. To demonstrate the technology independently of the growth factor application regimen, we used an established medium and growth factor protocol, published by Cai et al. with minor modifications. 6 In this study we also developed a twin-bioreactor system in which two bioreactors are operated under identical culture conditions except for the medium components.
Materials and Methods
Bioreactor, tubing, and perfusion units
The bioreactor model used in this study was previously described in detail.14,17,18 The system was developed as a culture model based on a four-compartment hollow fiber capillary membrane culture technology platform. 19 Briefly, each bioreactor contains two bundles of hydrophilic hollow fiber microfiltration membranes (400 kDa cut-off) to transport culture medium in a decentralized pattern mimicking arterio-venous perfusion, interwoven with one bundle of hydrophobic hollow fiber oxygenation membranes for the decentralized transport of O2 and CO220,21 in the cell compartment. Cells were evenly injected into the extracapillary space via 16 silicon tubes. Before the hESC inoculation, the bioreactor was primed with 1% Matrigel in Dulbecco's modified Eagle's medium/F12 to support survival of undifferentiated hESC in the bioreactor. The medium was pumped through the two-microfiltration fiber bundles in opposing directions (counter-current flow). 22 Flow of compressed air and CO2 through the oxygenation fibers was controlled by rotameters and adjusted according to daily blood gas analyzer readings of the medium pH. The bioreactor is integrated into a processor-controlled perfusion device (Stem Cell Systems GmbH), which was previously described in detail.17,18 The device contains two modular pump units, a heating unit, and a gas-mixing unit. The perfusion tubing with bubble traps was made of standard medical-grade dialysis PVC (B. Braun, Melsungen, Germany). Sterilization is performed with ethylene oxide according to clinical standards.
Expansion of undifferentiated hESCs
The hESCs cell line, SA002 (Cellartis AB), was used in this study. 23 The cell line was one of the NIH-registered hESC lines when the experiment was conducted. The project was approved by the University of Pittsburgh Human Stem Cell Research Oversight committee (hSCRO, ES-07-011-R). The hESC were cultured in center-well organ culture dishes (BD Biosciences) on mouse embryonic fibroblasts (CF1; Chemicon) in a humidified incubator at 37°C and 5% CO2. The culture medium VitroHES™ (Vitrolife AB) is supplemented with basic fibroblast growth factor (FGF) (Invitrogen) to a final concentration of 4 ng/mL. The medium was changed every 2–3 days (50%). Cells were mechanically passaged every 4–5 days under an Inverted Stereomicroscope (SMZ1000) (Nikon) using a stem cell cutting tool (0.29–0.31 μm) (Swemed Lab AB).
For undifferentiated expansion, cells were passaged using single-cell enzymatic dissociation. 24 Briefly, cells were dissociated using TripLE (Invitrogen) and cultured on human foreskin fibroblasts obtained from the American Type Culture Collection (CRL-2429) in a humidified incubator at 37°C and 5% CO2. The supplemented basic FGF concentration was increased to a final concentration of 10 ng/mL.
Twin-bioreactor 3D cultures
Two bioreactors were operated on one perfusion unit using two channel medium recirculation and -feed pumps under identical external conditions (Fig. 1). Only the medium composition varied to directly compare directed to spontaneous differentiation (Fig. 2). A total of 3 × 107 hESC cells were inoculated into each bioreactor. Three independent bioreactor cultures were performed for 20 days each. For an initial adaption and expansion period, the cells were cultured with mTeSR™ medium (STEMCELL Technologies) supplemented with 1% BD Matrigel™ (BD Biosciences) for 2 days. This prephase period was essential to support survival of hESC in the bioreactor.

Twin-bioreactor setup.
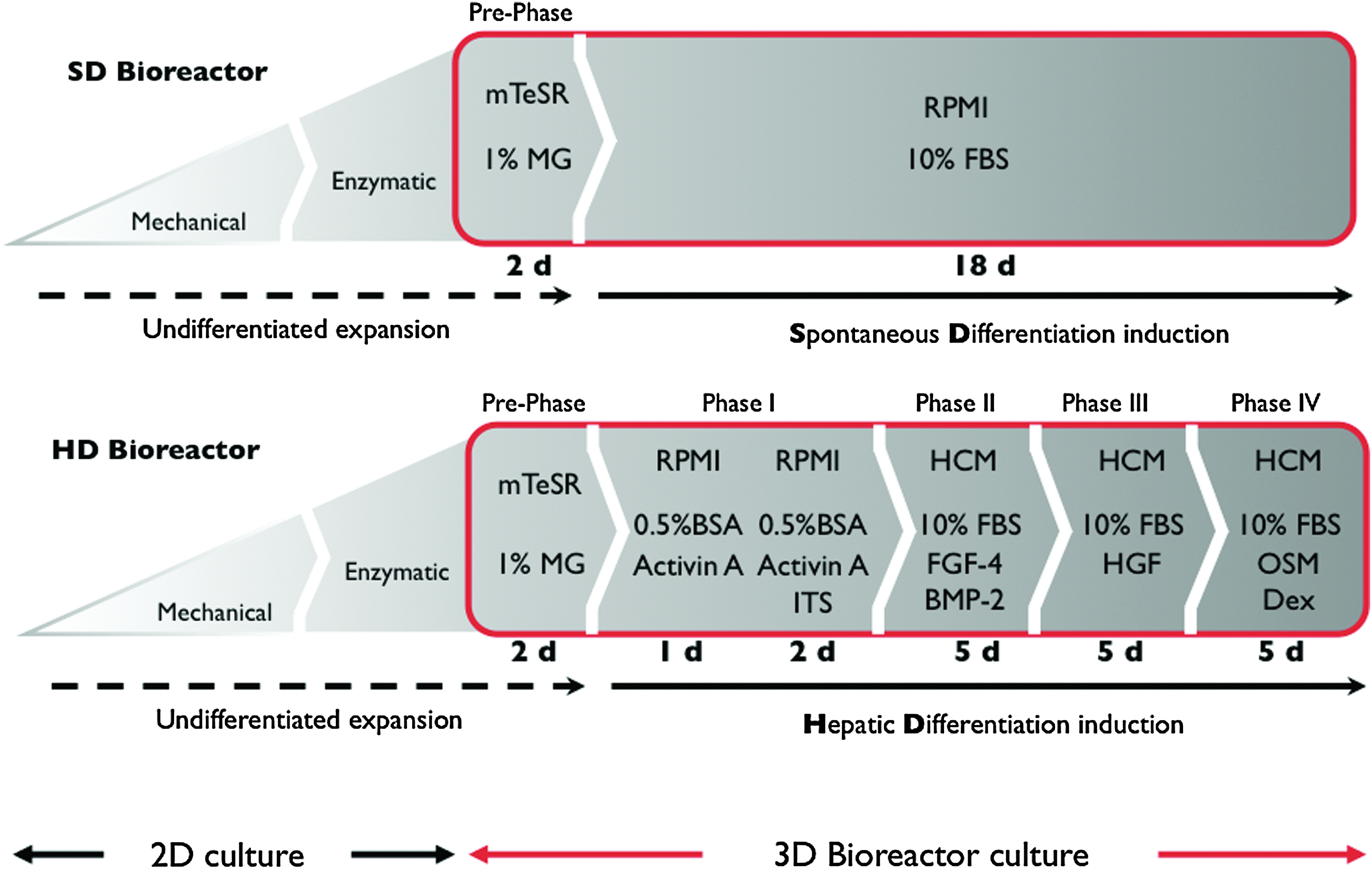
Stepwise hepatic differentiation (HD) strategy. For an initial endodermal differentiation, the cells were perfused with RPMI 1640 supplemented with 100 ng/mL activin A on days 3–5. For all following stages, the basal medium was switched to hepatocyte culture medium (HCM) (Lonza Inc.) supplemented with 1% insulin-transferring-selenium (ITS) and stage-specific cytokines and growth factors: during days 6–10, HCM was supplemented with 30 ng/mL fibroblast growth factor-4 (FGF-4) and 20 ng/mL bone morphogenetic protein-2 (BMP-2). For the next 5 days, the medium was changed to HCM and 10 ng/mL hepatocyte growth factor (HGF). For the last 5 days of culture and final maturation, HCM was supplemented with 10 ng/mL oncostatin M (OSM) and 0.1 μmol dexamethasone. For spontaneous differentiation (SD), RPMI 1640 supplemented with 10% fetal bovine serum (FBS) was used for the duration of the experiment after the 2-day initiation period (Pre-Phase). Color images available online at
For the following 18 days, the culture medium composition and supplementation conformed to the protocol published by Cai et al. with some modifications 6 (Fig. 2). Growth factors and cytokines were purchased from Peprotech: Activin A, FGF-4, bone morphogenetic protein-2, hepatocyte growth factor, and oncostatin M. To support hESC survival and efficient HD 10% fetal bovine serum was supplemented after Phase II of the HD protocol. For the spontaneous differentiation all conditions, except for the medium composition, were identical to the 3D directed HD, including the inoculated cell number, perfusion conditions, and culture period. For spontaneous differentiation the culture medium was changed after the 2-day initiation period (Pre-Phase). RPMI 1640 supplemented with 10% fetal bovine serum was used for the remaining duration of the experiment (Fig. 2).
2D HD control culture
One million hESC were cultured on 60 mm culture dishes (BD Biosciences) coated with 1% BD Matrigel. Cells were cultured in a humidified incubator at 37°C and 5% CO2. The culture medium was changed daily, and the stage-specific basal medium supplied with cytokines was added according to the differentiation protocol (Fig. 2). Control 2D cultures were repeated four times.
Metabolic activity monitoring
Metabolic activity of the cells inside the bioreactors was quantified on a daily basis by measuring the concentrations of glucose and lactate in the culture medium with an i-STAT beside analyzer (Abbott Point of Care). The concentrations were normalized by inoculated cell number and feed rate using the formula below to compare each individual bioreactor experiment.
P: metabolite production rate (μg/[million cells]) F: medium feed rate (mL/h) Cn: metabolite concentration on day n (μg/mL) V: total medium volume of bioreactor including tubing (mL)
Liver-specific gene expression
Cells were harvested before inoculation into the bioreactor (Pre-BR), and after bioreactor culture (Post-BR) (d20) and control dish culture (2D Cont). Total RNA was isolated from the cell samples with mRNA isolation kit (QIAGEN) and RNA quality was verified with SYBR green on a 2% agarose gel. Complimentary DNA was synthesized with a conventional reverse transcription process (Promega). Quantitative real-time polymerase chain reaction (qRT-PCR) analysis was performed using an ABI PRISM 7000 and predesigned TaqMan probes and primer sets for albumin (Hs00609411_m1), alpha 1-antitrypsin (A1AT, Hs00165475_m1), cytochrome P450 (CYP)3A4 (Hs00430021_m1), CYP3A7 (Hs00426361_m1), hepatocyte nuclear factor 4 alpha (HNF4a, Hs00230853_m1), glucose-6-phosphatase (Hs00609178_m1), arginase 1 (ARG1, Hs00968978_m1), argininosuccinate lyase (ASL, Hs00902699_m1), argininosuccinate synthetase 1 (ASS1, Hs00540723_m1), carbamoyl-phosphate synthetase 1 (CPS1, Hs00157048_m1), ornithine carbamoyltransferase (OTC, Hs00166892_m1), and β-actin (Hs99999903_m1) from TaqMan Gene Expression Assays (Applied Biosystems). Ct- values were processed using Relative Expression Software Tool 200825,26 and normalized to β-actin to account for total amount of RNA in each individual sample.
To elucidate the induction of drug-metabolizing enzymes of the CYP family in hESC-derived hepatocytes, rifampicin was added to the culture medium of one system to a final concentration of 25 μmol/mL for the last 5 days of culture. Gene expression of CYP3A4 and the fetal form CYP3A7 were evaluated by qRT-PCR analysis.
Flow cytometry
Before differentiation the undifferentiated state of the hESC was validated by the expression of the pluripotency markers SSEA-4, TRA-1-60, TRA-1-81, and IGFR-1a (data not shown). To determine HD of hESC, flow cytometric analyses were performed with an antibody against asialoglycoprotein receptor (ASGPR) (Cell Sciences) and Alexa 488–conjugated goat anti-mouse IgG secondary antibody (Invitrogen) before and after the culture. Isotype control antibody (mouse IgG1 isotype control, HI1001; Cell Sciences) was used as negative control for the measurement of nonspecific binding of mouse monoclonal antibodies of IgG1 isotype. The control samples were prepared and processed along with the ASGPR Ab staining for each experiment. Analyses were performed on a FACS-Calibur using the Cell Quest software (Becton Dickinson). The forward scatter and side scatter gates were set to include all viable cells. Postacquisition analysis and desktop publishing were accomplished using the flow cytometry analysis software FlowJo (Tree Star, Inc.). The percentage of positive cells was measured from a cut-off set according to the isotype-matched nonspecific control.
Albumin secretion
Medium samples were harvested every day and stored at −150°C until analysis. The concentration of human albumin was measured using ELISA Quantitation kit (Bethyl Laboratory) under the manufacturer's instructions. Concentrations were normalized to the inoculated cell number and feed rate to compare each individual bioreactor experiment as described above.
Ammonia metabolism activity
To evaluate hepatic detoxification function, an ammonia challenge was performed on the last day of culture (d20). The cells were incubated with 5 mM ammonium chloride (NH4Cl; Sigma-Aldrich) for 4 h. During this time, the bioreactor system was operated without fresh medium supply to eliminate dilution. After incubation, samples were collected and urea concentration was measured using a colorimetric assay kit (Urea Colorimetric Assay Kit, QuantiChrom; BioAssay Systems).
Histological and immunofluorescence analysis
The upper part of the bioreactor was opened by using a circulate saw (diameter 1.5 inch). Using a scalpel, the bundled hollow fibers were cut along the inner wall of the compartment to maximize sample volume. The hollow fibers were harvested en-bloc (two to four fiber bundles) and immediately embedded in 2% low melting agarose gel to preserve structures. The specimens were fixed with 4% paraformaldehyde or 10% buffered formalin for immunofluorescence and hematoxylin and eosin (H&E) staining, respectively. H&E staining was performed according to standard laboratory procedures. Periodic Acid-Schiff (PAS) staining was carried out with PAS staining system (Sigma-Aldrich) to observe glycogen in the tissue-like structure using the manufacturer's directions.
Primary antibodies used for immunofluorescence were mouse anti-albumin (Sigma Aldrich), rabbit anti-cytokeratin (CK)-8 (Abcam), mouse anti-CK18, rabbit anti-CK7 and mouse anti-CK19, rabbit anti-HNF4a (Santa Cruz Biotechnology), and rabbit anti-CYP3A4 (Cypex Ltd.). Secondary antibodies used were goat anti-mouse Alexa Flour 488 (Invitrogen) and goat anti-rabbit Cy3 conjugated (Dianova). Nuclei were observed with 4′-6-diamidino-2-phenylindole staining. An inverted microscope (Axiovert 200M; Carl Zeiss) equipped with a CCD camera (Retiga 2000R; QImaging) was used for microscopic observation and imaging. The files were processed using the digital imaging software Image Pro Plus (Media Cybernetics). Adjacent sections served as negative controls and were incubated with appropriate isotype control antibodies and secondary antibodies.
Statistical analysis
Statistical evaluation was performed using Prism version 5.0c (GraphPad Software Inc.). Results are listed as mean ± standard deviation. One-way analysis of variance followed by Tukey–Kramer multiple comparison testing was applied where multiple comparisons were made. Differences were considered to be statistically significant at p < 0.05.
Results
Metabolic activity
After inoculation, the metabolic activity of the cells in the bioreactors, estimated by glucose consumption and lactate production, was stable throughout the culture period (Fig. 3A, B). No statistically significant difference of the metabolic activity was observed between the two bioreactors in each twin-bioreactor setup of spontaneous and directed differentiation (p = 0.3685). The metabolic activity data indicate that hESC were successfully inoculated in the space between the hollow fibers and stably cultivated within the dynamic perfusion system.

Metabolic activity of human embryonic stem cell (hESC) in three-dimensional (3D) perfusion culture.
Endodermal and hepatic marker gene expression
The expression of six liver-specific genes was analyzed by qRT-PCR in HD, spontaneous differentiation (SD) 3D perfusion bioreactor, and conventional 2D HD cultures. Gene expression levels were normalized to an internal control gene (b-actin). The relative expression to the starting materials were plotted to investigate the maturation of hESCs during 3D perfusion bioreactor and 2D static culture. As expected, all endodermal and liver-specific gene expressions were increased after 20 days in culture with HD (2D or 3D) or 3D spontaneous differentiation induction (Fig. 4A–E). One of the endodermal marker genes, HNF4a, expression was higher in the 2D static culture (2D), whereas expression of all other HD marker gene was significantly high in the 3D HD group. Interestingly, early phase HD marker, albumin, and alpha-1 antitrypsin (A1AT) genes were expressed in the 3D spontaneous differentiation group on a comparable level with the 2D HD group (Fig. 4C, D). These data indicate that some of the hESC differentiated into hepatic lineage without any exogenous humoral stimulation. These spontaneously differentiated hepatic lineage cells, however, did not express the late-phase HD marker glucose-6-phosphatase. The 2D HD culture showed considerably lower G6P gene expression, compared to 3D directed differentiation (Fig. 4E). These data showed that the 3D perfusion culture system enabled a more efficient HD in hESC.

Real-time polymerase chain reaction analysis for relative endodermal and hepatic gene expressions at day 20.
CYP3A4 and 3A7 expression
CYP3A4 is one of the most important drug metabolism enzymes in human liver, whereas CYP3A7 plays this role in the fetal liver. The expression pattern and the level of these CYP genes are therefore important to evaluate hepatic maturation. 27 To evaluate the functional CYP genes expression, cells in the 3D HD group were treated with rifampicin and gene induction was determined (Fig. 4F). Both CYP3A4 and 3A7 expression was increased up to twofold by the rifampicin induction. Further, CYP3A4 gene expression was relatively higher than CYP3A7 gene expression. The results showed that although some of the hESC-derived hepatic cells in 3D culture were still in the developmental state of fetal hepatocyte, some of the cells responded to rifampicin induction with CYP3A4 gene upregulation like mature hepatocytes. The CYP3A4/3A7 ratio was 1.3 after the rifampicin induction. Although there is no statistical significance between 2D and 3D culture system due to the wide range of deviation and insufficient sample number (n = 3), the CYP3A4/3A7 ratio at the endpoint of the experiments (d20) was tentatively higher in 3D culture group (235.4% ± 436.4%) than that in 2D culture control group (2.688% ± 1.817%). Uneven maturation of hES-derived hepatic cell and uneven localization of the mature hepatic cells in the cell compartment of the bioreactor might cause the variation of the gene expression data.
Flow cytometry analysis shows hepatic maturation of the hESC
To quantify how many hESC were differentiated to mature hepatocytes in each culture condition, flow cytometric analyses were conducted with anti-ASGPR1 antibody. ASGPR1 is a liver-specific membrane receptor and has been utilized as a marker for mature hepatocytes.28–31 While the cells from 2D culture contained only a small fraction of ASGPR1-positive cells (2.36% ± 0.54%), 28.56% ± 1.81% of the cells in 3D HD culture were positive for ASGPR1. Under 3D spontaneous differentiation conditions, 13.46% ± 0.03% of cells expressed ASGPR1 (Fig. 5). Compared to static 2D culture, the 3D perfusion culture system increased the mature hepatocyte marker positive cells for 11.9-fold. ASGPR1 expression was not detected on the hESC before differentiation.

Flow cytometry analysis of hepatic marker-positive cells of cultured hESC in 2D HD, 3D SD, and 3D HD cultures (n = 3). Dotted line represents isotype control, and bar represents percentage (means and SD) of positive cells compared to isotype control.
Functional analysis of hESC-derived hepatocyte-like cells
Hepatic synthetic function of hESC-derived hepatocyte-like cells was confirmed by the detection of albumin secretion by ELISA analysis of culture medium from daily samples (Fig. 6A). After 10 days in culture (HD phase III), albumin secretion was detected in the medium of both 2D and 3D culture groups but not in the 3D spontaneous differentiation group. The normalized data showed that albumin production and secretion was significantly active in the 3D culture system.
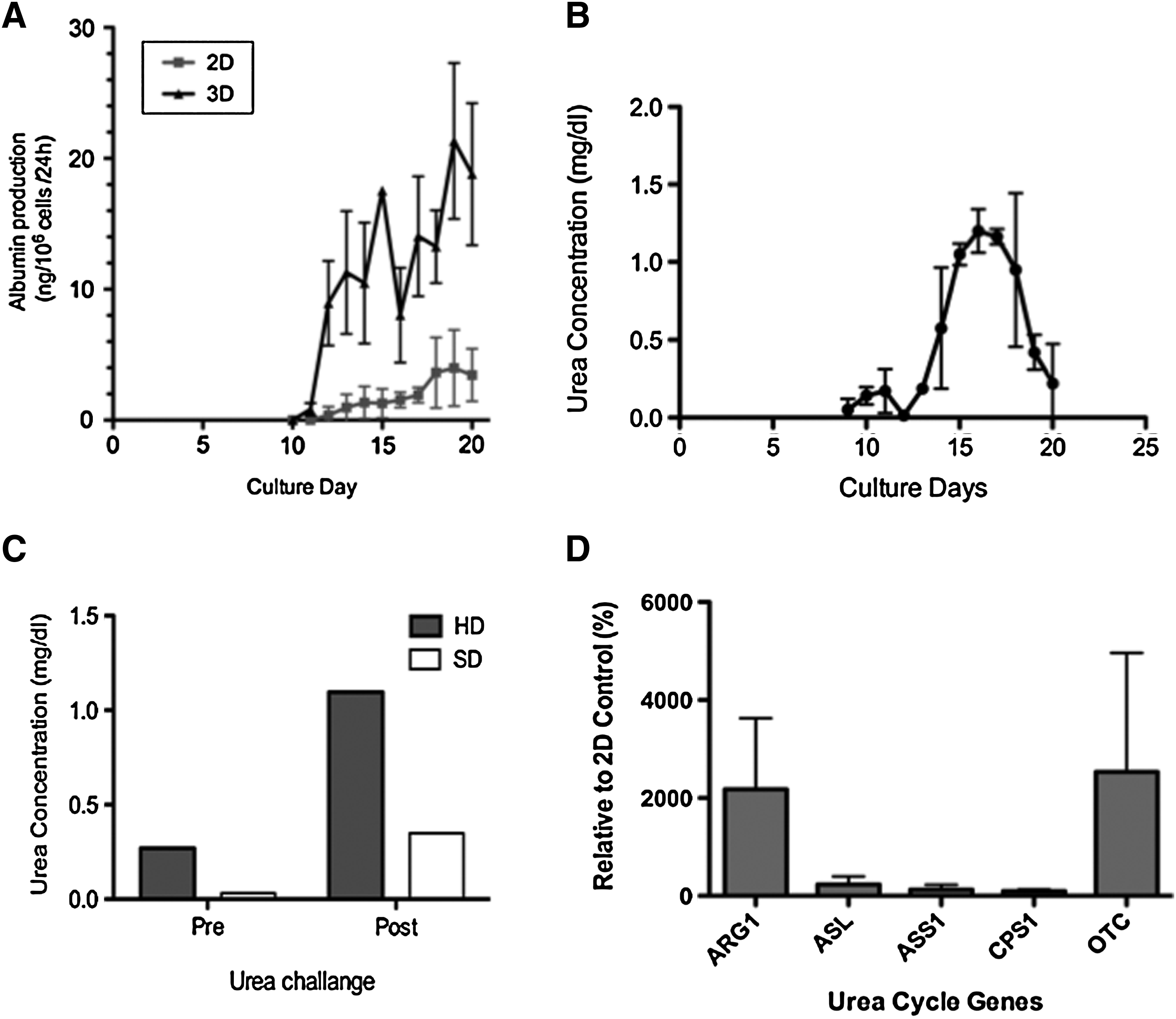
Hepatic function of hESC-derived hepatocyte-like cells.
Urea metabolism and secretion capability of hESC-derived hepatic cells were also investigated. Intrinsic basal urea production was measured in daily culture medium samples from all experiments (n = 3 each). The linear detection range of the colorimetric assay kit is 0.006 mg/dL (1 μM) to 100 mg/dL (17 mM) urea in 96-well plate assay. Only samples from 3D HD culture showed a detectable level of urea production around day 10 of culture (Fig. 6B). The urea production increased in a time-dependent manner and interestingly peaked at day 16 and declined after day 17, whereas all other metabolic activity indicated the cell cultures were stable. Considering the lag in response time, this data indicates the phase III HD media containing hepatocyte growth factor enhances the urea metabolism function, and phase IV media containing oncostatin M and dexamethasone did not contribute to an increase urea production. The ammonia challenge data on day 20, however, indicates that the ESC-derived hepatic cells maintained their urea production and detoxification capability. The ammonia challenge test was performed in all experiments. In the 3D HD culture group, strong activity was detected as 4.1-fold urea production after the 4-h incubation with ammonia. Although the baseline was relatively low, the 3D spontaneous differentiation group also showed some increases of urea production (Fig. 6C).
The acquisition of urea metabolism function is confirmed by the gene expression of major urea cycle-related enzymes, ARG1, ASL, ASS1, CPS1, and OTC. All studied key urea cycle genes were expressed in both 2D and 3D HD culture samples at the endpoint of the experiment (day 20). As shown in Figure 6D, ARG1 and OTC gene expressions in 3D culture system were significantly higher (21.76-fold and 25.32-fold, respectively) than that of in 2D culture system. Other genes' expression, including the rate-limiting enzyme, ASS1, in the 3D culture were in comparable range with 2D culture system.
Histology and immunofluorescence show hepatic structures in 3D culture
In the 3D HD culture, cell clusters with typical hepatocyte morphology were identified in H&E staining (Fig. 7A). The hepatocyte-like clusters were also positive for PAS staining (Fig. 7B), confirming glycogene storage function of the cells. Immunofluorescence analysis showed that most of the cells were positive for CK18 and CK8, indicating hepatic lineage cells (Fig. 8A–D). Large cell aggregates were stained positive for albumin accompanied with nuclear HNF4a staining (Fig. 8E–H). Some of the albumin-positive cells were also positive for CYP3A4 (Fig. 8I–M). The albumin-positive cells were clearly distinguished from biliary cell marker CK19-positive cells. The CK19-positive cells were coexpressed with mature biliary marker CK7. These CK19 and CK7 double-positive cells formed bile duct-like tubular structures in the 3D culture system. The CK19 and CK7 double-positive cells formed a single-layered ring of cells without adjacent endothelial cells. Further, there was no vessel-like structure, including portal vein, hepatic vein, and hepatic artery. Also, there is no significant morphological difference that indicates the hilum or the periphery of liver structure or zonal variations. These data suggest that the differentiated hESC in 3D HD culture form structures that resemble primitive in vivo-like tissue structures of parenchymal and nonparenchymal hepatic cells. These formations were not observed under conventional 2D culture.

Representative section for hematoxylin and eosin and immunohistochemical staining.

Immunofluorescence staining for hepatic and biliary marker proteins in 3D HD group.
Discussion
In this study a dynamic 3D perfusion culture system was investigated for its ability to facilitate HD in hESC compared to the standard static 2D culture, when using the same media and growth factor regimen. The limitation of hepatocyte culture on 2D flat culture surface is well documented.32,33 In vivo, hepatocytes are organized into a polarized epithelium with distinct apical and basal domains. This hepatic polarity and the response of cells in the liver cannot be reproduced in 2D monolayer culture systems. 3D perfusion culture enhances physiological cell-to-cell interactions that should provide crucial physical stimuli to hESC, in addition to the humoral stimuli with cytokines and growth factors in the culture medium. Dynamic 3D perfusion with constant slow medium feed and faster medium recirculation also provides a more in vivo-like homeostatic environment by constantly supplying humoral factors and nutrition with low gradients, and removing cell products avoiding gradient built-up, whereas the integral oxygenator serves for decentralized oxygen supply and CO2 removal also with low gradients. 17 It has already been shown that static 3D culture conditions improve survival for hepatocytes in vitro.13,34,35 These biochemical and topological elements of 3D perfusion culture also affect the differentiation and maturation process of stem cells. 36 3D culture systems have been applied to induce HD in several studies.37–40 These studies presented static 3D culture systems using biodegradable polymer scaffolds or extracellular matrixes enable to provide effective HD of ESCs. Previously, our group showed that the dynamic 3D perfusion culture system used can prolong hepatic functions of primary adult hepatocytes 15 and enhance functional maturation of fetal hepatocytes.14,17 This study further supports existing data showing advantages of 3D perfusion culture conditions.
To test the influence of media factors in 3D culture on the hESC, two identical bioreactors were used in one perfusion unit with the dual channel pump heads and inoculated the same number of hESC, but used different medium supplementations. The metabolism data indicate that both conditions encouraged cell proliferation. The qRT-PCR gene expression analysis data suggested that some of the hESC were spontaneously differentiated into hepatic lineage cells even without specific growth factors. The relative expression levels, however, were low, and the late-phase marker gene (G6P) was not detected. Although it was not significant, the HNF4a gene expression was relatively higher in the 2D HD control culture samples. According to the transcriptome studies on mouse liver development, the HNF4a gene is expressed in a relatively early stage of liver development (ED8.5 in mouse). Jochheim et al. demonstrated that HNF4a gene expression peaked on ED11.5 and decreased until ED13.5, then gradually increased but never again reached as high as ED11.5 in the mouse ESC HD process. 41 Although hESCs were used in the present study, the above-cited data provide a basis to speculate that the ES-derived hepatic cells in 2D culture system were at the high HNF4a expression stage (equivalent to ED11.5 of mouse liver), whereas the cells in 3D system were further differentiated.
No albumin-positive clusters or bile duct-like tubular structures were observed in the spontaneous differentiation bioreactors, indicating that the 3D perfusion conditions alone were not sufficient to induce hepatic maturation but enhanced the action of a growth factor application regimen. These advantages should be in effect when cell-to-cell interactions are established under high-density culture.
The drug inducability of CYP gene family is an important marker for mature functional hepatic cells. The P450 family is crucial for the metabolism and elimination of drugs, xenobiotics, and endogenous agents such as steroids.42–44 The induction of CYP3A4 and 3A7 by stimulating the differentiated cells with rifampicin was observed. 45 The degree of maturation of hESC-derived hepatocytes was further evaluated by flow cytometry analysis for ASGPR1 expression. 46 The hESC-derived hepatic cells from 3D dynamic perfusion culture contained up to 31.7% ASGPR-positive cells. The ASGPR-expressing hESC-derived hepatic cell is of interest for in vitro models to study the hepatotropic viruses such as hepatitis virus, which uses this receptor to infect cells. 30 In addition to the ASGPR expression, infection of hepatitis C virus (HCV) requires a tight junction to polarized the apical surface of hepatocytes. 47 The insufficient polarization of cultured hepatocytes in 2D culture prohibits establishing in vitro model systems for HCV research. Since the infection of HCV is strictly limited to human hepatocytes, suitable in vitro systems are desired to pursue HCV research, including antivirus drug discovery.48,49 Detoxification function is one of the most important hepatic functions. To evaluate the detoxification function of hESC-derived hepatic cells, an ammonia challenge was performed by overloading the each culture system (2D and 3D) with ammonium chloride (5 mM). Urea cycle-related gene induction and 4.1-fold urea production were observed in the 3D HD culture group, whereas the 3D spontaneous differentiation group also showed some increases of urea production. Although no increase of lactate dehydrogenase as a marker for cell injury was detected in the culture medium after the ammonia challenge, the 5 mM ammonium chloride could cause injury to the hESC-derived hepatic cells and lead to the release of arginase from the cells. Thus, there is a possibility of production of urea associated with nonspecific arginase activity that degrades released arginine to ornithine and urea. 50 The urea cycle consists of five enzymatically controlled steps. The gene expression of the five key enzymes was investigated on 2D and 3D culture sample at the endpoint of the HD protocol. Interestingly, 3D culture system increased ARG1 and OTC gene expression in hES-derived hepatic cells, whereas CPS1, ASL, and ASS1 gene expression levels were similar to 2D culture system. This qRT-PCR data indicate that ARG1 and OTC genes might be induced by 3D culture-specific signals. Further investigation will be required to elucidate the signals and mechanism of influence of 3D culture to these particular genes.
The immunohistochemistry data provided further evidence that cells supplied with specific cytokines under 3D conditions could more efficiently differentiate toward hepatic lineage. Liver mass-like PAS staining positive hESC-derived hepatocytes and duct-like structures formed in the 3D bioreactor. The culture conditions induced not only differentiation of hepatic lineage cells but also cells of biliary cell lineage. Proximity of hepatic and bile duct-like cells resembles a primitive organ-like structure.17,51 Cells after spontaneous differentiation under 3D conditions and 2D directed differentiation did not exhibit similar structures. During the liver development, intrahepatic bile duct morphogenesis progresses from the hilum of the liver to the periphery of the lobes. The earliest biliary differentiation occurs during the formation of the ductal plate (a continuous ring of cells) adjacent to the portal vein. 52 CK19 and CK7 double-positive cells that form a single-layered ring of cells were found in the 3D HD culture condition. Nevertheless, no vessel-like structures including portal vein, hepatic vein, and hepatic artery was detected. Also, there is no significant morphological differences indicating the hilum or the periphery of liver structure. Further investigation will be required to elucidate the mechanism of such bile duct-like structure formation in the 3D culture system without scaffolds of endothelial cells.
The findings in this study suggest that directed differentiation of hESC in dynamic 3D perfusion culture provides a promising approach to effectively derive functional hepatic cells. There is a possibility that physical parameters such as flow, stress forces, and pressure might play a vital role in the development and differentiation of hESC into lineage committed cells and tissue structures in addition to the 3D topological parameters. The concept of the bioreactor resembles nature's concept of capillaries providing active mass exchange, and it is likely that not only final concentrations of specific growth factors are important, but also gradual changes are crucial for the process of differentiation. While a standard Petri dish culture medium is changed abruptly, the constant perfusion and gradual substitution of medium and supplements more closely resembles the natural process of growth and development in vivo. Future studies will be aimed to elucidate the influence of the physical parameters on the differentiation to improve the differentiation protocols. Prolonged experimental conditions might increase maturation. Another enhancement might be the use of specific extracellular matrix components to support liver-specific cell fate.53–55
Previously, we have shown that the dynamic 3D perfusion culture system enabled larger-scale undifferentiated ESC expansion using mouse ESCs. 56 The dynamic 3D perfusion culture also promoted spontaneous in vitro differentiation of mouse ESCs when cultures were conducted for several weeks. 18 These results indicate a feasibility of initially expanding undifferentiated hESC cells in larger-scale, followed by a specific differentiation regimen in the same culture system. Easy scale-up of the system to larger bioreactors for potential extracorporeal liver support studies, the use of medical-grade components, and closed system conditions provide a promising tool for possible preclinical studies in the future. A clinical scale version of the here-described bioreactor technology platform was already used for extracorporeal liver support research and—using primary human liver cells—in clinical applications.57,58
In conclusion, dynamic 3D perfusion culture in bioreactors supports HD of hESC in vitro, when exposed to a specific differentiation regimen. In contrast to conventional 2D culture, the 3D perfusion culture systems induced more functional maturation to hESC-derived hepatocytes. The derivation of functional human hepatic cells from hESC may provide an unlimited cell source for analyzing the molecular mechanisms involved in liver development for toxicology, drug discovery, virology, and bioartificial liver support system development research. The twin 3D perfusion culture bioreactor application method presented could be a powerful in vitro model system for liver function and disease.
Footnotes
Acknowledgments
The authors thank Cellartis AB for provision of the SA002 cells and Dr. Petter Björquist for fruitful discussions. We also thank Patrick Over and Matthew Young for technical assistance.
Disclosure Statement
No competing financial interests exist.