
Abstract
Autologous heart valve tissue engineering relies on extracellular matrix production by cells seeded into a degrading scaffold material. The cells naturally exert traction forces to their surroundings, and due to an imbalance between scaffold, tissue, and these traction forces, stress is generated within the tissue. This stress results in compaction during culture and retraction of the leaflets at release of constraints, causing shape loss of the heart valve leaflets. In the present study, an in vitro model system has been developed to quantify stress generation, compaction, and retraction during culture and after release of constraints. Tissue-engineered (TE) constructs based on polyglycolic acid/poly-4-hydroxybutyrate scaffolds seeded with human vascular-derived cells were cultured for 4 weeks. Compaction in width was measured during culture, stress generation was measured during culture and after release of constraints at week 4, and contraction was measured after release of constraints at week 4. Both compaction and stress generation started after 2 weeks of culture and continued up to week 4. TE constructs compacted up to half of their original width and reached an internal stress of 6–8 kPa at week 4, which resulted in a retraction of 36%. The model system has provided a useful tool to unravel and optimize the balance between the different aspects of TE constructs to develop functional TE leaflets.
Introduction
Autologous TE heart valves are fabricated by seeding extracellular matrix (ECM)-producing cells on a degradable scaffold material and culture this in vitro. During the culture period the scaffold material degrades and the ECM produced by the cells takes over. In vivo results of autologous heart valves have shown promising results up to 8 months.4–6 Arterial- or venous-derived cells are commonly used as cell sources for this approach. 7 These cells are associated with high ECM production, but their contractile properties may be a source of problems. In our group, TE heart valves are grown with the leaflets attached to each other to assure constraint tissue culture, which has shown to be beneficial for tissue formation. 8 In this constrained situation, the cells are able to generate stress within the tissue, due to their contractile properties. Just before implantation, the leaflets are separated to enable opening and closing of the valve in vivo. The stress that has been build up within the tissue during culture causes retraction of the leaflets once constraints are released (Fig. 1). If this retraction is severe, the heart valve will not be able to fully close during diastole and its functionality is lost.

An ovine tissue-engineered heart valve after 4 weeks of culturing with the leaflets attached to each other
Tissue retraction is a serious issue within heart valve tissue engineering as dysfunctional closing of the valve leaflets leads to regurgitation. Valvular regurgitation, related to decreasing leaflet length, has been observed in both natural and synthetic scaffold-based autologous heart valves.6,9 Tissue retraction results from an imbalance among the three major components of engineered heart valve tissues: the cells, the scaffold, and the newly formed tissue. The cells within the tissue will always aim to develop a certain internal stress 10 and exert traction forces to the ECM and scaffold surrounding to achieve this. In the early phase of tissue culture, the scaffold is stiff enough to withstand the contractile forces of the cells. However, after approximately 2 weeks of culture, the scaffold rapidly degrades, whereas the newly formed tissue is not capable yet to withstand the contractile forces. The changes in cell traction, scaffold, and tissue during culture are illustrated in Figure 2. As a result of these changes, two processes of tissue shrinkage can be distinguished. During culture, tissue compaction occurs by cell-traction-mediated remodeling of the construct. Upon release of the constraints, retraction occurs immediately due to release of the stress generated within the matrix by the cells during culture and slowly continues due to retraction of the matrix by the cells. 11
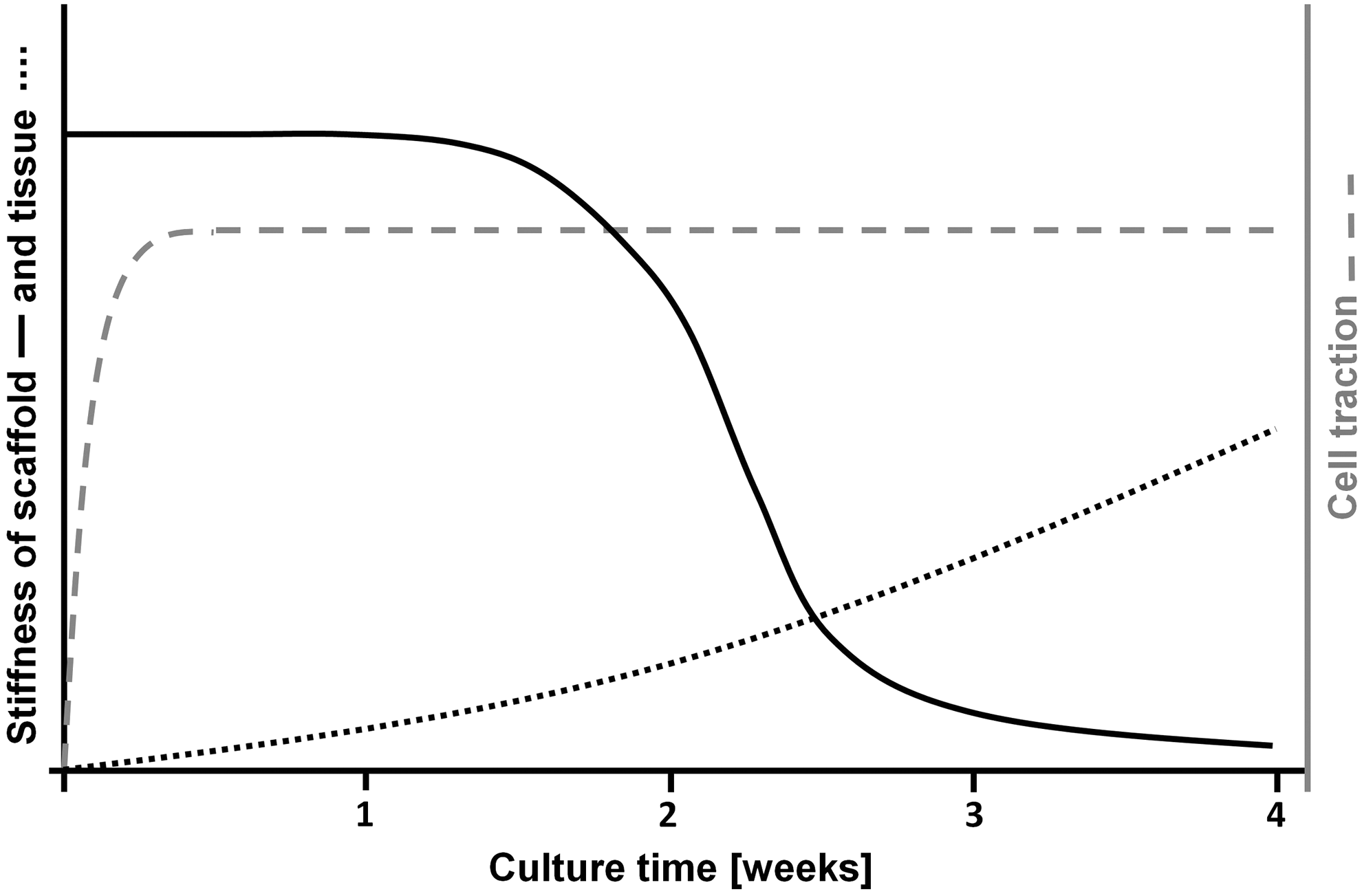
Illustration of the changes in scaffold and tissue stiffness, and cell traction during 4 weeks of culture of tissue-engineered constructs. The scaffold degrades rapidly after 2 weeks, tissue stiffness gradually increases over time, whereas total traction force of the cells will quickly reach its maximum and will stay at that level.
To solve the problem of tissue shrinkage, it is important to know how the imbalance between scaffold, tissue, and cell traction results in stress generation, compaction, and tissue retraction. Therefore, the aim of the present study was to quantify and correlate tissue compaction, retraction, and stress generation in engineered heart valve tissue during a culture period of 4 weeks. We developed an in vitro model system in which stress generation, compaction, and retraction can be measured in one single engineered tissue. Proof of principle of this model system is studied by culturing engineered heart valve tissues based on human vascular-derived cells seeded onto a rapid degrading scaffold within this model system for 4 weeks. Stress generation, compaction, and retraction were quantified during culture and after release of constraints at week 4. Using this model system, we are able to gain insight in the development of stress, needed to unravel the balance between tissue, scaffold, and cell traction. Therefore, this model system is of great value for investigating the applicability of possible solutions to prevent or compensate for tissue retraction toward a functional TE heart valve.
Materials and Methods
Experimental setup
An in vitro model system is developed in which generated stress, compaction, and retraction of one TE construct can be quantified during culture and after release of constraints (Fig. 3A). The model system consists of a stainless steel frame in which two ultra-high-molecular-weight polyethylene sliding blocks are positioned opposite from each other. In between the two sliding blocks, the TE constructs are cultured. One sliding block can be either fixed (during culture) or move freely (to measure retraction after culture, Fig. 3B). The other is connected to the metal frame via two leaf springs and the displacement of this sliding block is related to the generated force within the tissue (Fig. 3C). The sliding blocks can be fixed with clamps (Fig. 3A-1, 2). To measure force generation during culture, the sliding block of the leaf springs is not fixed. Because the force that is generated by the cells will deform the leaf springs, this configuration is called semi-constrained. To measure generated force after release of constraint, both sides are fixed and this configuration is called constrained.

Photographs (ABC-1) and a schematic overview (ABC-2) of model system in which retraction (RL, B1-2) and generated force (F, C1-2) can be measured through the displacement (ν) of two sliding blocks positioned opposite from each other.
Both generated force and retraction are measured through the displacement of the sliding blocks by comparing the displacement of a black dot on the sliding block to a reference dot on the metal frame. To quantify the displacement during culture (generated force), photographs were made twice a week and analyzed with ImageJ (WCIF ImageJ, National Institutes of Health). Displacements at week 4 (retraction and generated force after release of constraints) were assessed from pictures taken underneath a stereomicroscope (Zeiss Observer; Zeiss) and quantified with Matlab (The MathWorks).
Quantification of the generated stress
Theoretically, the force F generated by the TE construct (mN) can be calculated from the displacement ν of the sliding block (mm) and the properties of the leaf springs from the equation for bending of a straight beam
12
:
where E is the Young's modulus of the beam material (mN/mm2), l is the moment of inertia determined from the width b (mm) and height h (mm) of the leaf spring (
This was done by measuring the displacement of the sliding block during three cycles of subjection to known applied forces ranging from 0 to 20 mN. Loading was within the linear region of the leaf springs verified by the linearity of the force–displacement curves.
To translate the generated force to stress σ (kPa), the cross-sectional area A (mm2) of the TE constructs was measured. Width was assessed during culture from photographs and underneath a stereomicroscope at week 4. The initial thickness at day 0 was 1 mm, representing the thickness of the scaffold. Thickness at week 4 was assessed from histological sections (see under Tissue formation section), assuming a linear shrinkage factor of 1.043 during processing of the tissue for histology.
13
Thickness during culture could not be measured and, therefore, the thickness at earlier time-point was interpolated assuming a change over time with a course similar to that of the width. The calculated cross-sectional area was used to determine the generated stress via:
Tissue culture
Vascular-derived cells were harvested from the human vena saphena magna obtained according to the Dutch guidelines for secondary used materials. These cells have previously been characterized as myofibroblasts. 14 They show expression of vimentin, but not desmin and a subpopulation of the cells express α-smooth muscle actin. Cells were expanded using regular cell culture methods in a humidified atmosphere containing 5% CO2 at 37°C. Culture medium consisted of advanced Dulbecco's modified Eagle's medium (Invitrogen), supplemented with 10% fetal bovine serum (Greiner Bio one), 1% GlutaMax (Invitrogen), and 1% penicillin/streptomycin (Lonza). The cells were seeded onto rectangular scaffolds (18×5×1 mm) of rapid degrading nonwoven polyglycolic acid (Concordia Manufacturing Inc.) coated with poly-4-hydroxybutyrate (Tepha; received through collaboration with University Hospital Zurich). The scaffolds were attached to the sliding blocks with polyurethane-tetrahydrofuran glue (15% wt/vol). Sterilization was achieved by 70% ethanol incubation for 30 min. Cells were seeded at passage 7 with a seeding density of 15 million cells/cm3 using fibrin as a cell carrier. The TE constructs were cultured in rectangular well plates for 4 weeks in a humidified atmosphere containing 5% CO2 at 37°C. During tissue culture, the medium was supplemented with L-ascorbic acid 2-phosphate to promote ECM production (0.25 mg/ml; Sigma), and replaced twice a week.
Experimental design
TE constructs were cultured for 4 weeks in constrained (n=5) and semi-constrained (n=5) configuration. During culture, the compacted width Wcomp (μm) of both constrained and semi-constrained TE constructs (n=10) was assessed twice a week. Compaction CW (%) was defined as the percentage of shrinkage compared with the original width W0:
Stress generation during culture was measured in the semi-constrained TE constructs twice a week. After 4 weeks of culturing, generated stress and retraction were measured in the constrained samples after release of the constraints. First, generated stress was assessed during 30 min with time intervals of 3 min, after which retraction in length RL (%) was measured during 24 h as the percentage of shrinkage compared with the original length L0 (μm):
in which Lretract is the retracted length (μm) of the TE construct. Time intervals between retraction measurements were 6 min in the first half hour, followed by measurements after 1, 2, 4, 6, 16, and 24 h. An overview of the experimental design is given in Figure 4.

Schematic overview of the experimental design. Compaction during culture was measured in both constrained and semi-constrained samples. Retraction and generated stress were measured at week 4 after release of constraints in the constrained samples. Generated stress during culture was measured in the semi-constrained samples. PGA, polyglycolic acid; P4HB, poly-4-hydroxybutyrate.
Tissue formation
Tissue quality was evaluated by histological staining. The TE constructs were fixed in 3.7% formaldehyde (Merck) and embedded in paraffin. Tissue sections of 10 μm were cut and studied by hematoxylin and eosin staining for general tissue morphology and Masson-Trichrome staining for deposition of collagen. The stained sections were evaluated using light microscopy (Axio Observer; Zeiss). Additional sections were stained with Picrosirius red to observe mature collagen fibers and evaluated using polarized light microscopy.
Statistical analyses
All data are presented as mean and their standard error of mean. Linear regression analysis was performed to fit the calibration curves using GraphPad Prism software (GraphPad Software Inc.).
Results
Calibration of the model systems
Given the constants of the materials and the model system (see quantification of generated stress section), the theoretical correlation between force (F in mN) and displacement (ν in μm) becomes F=0.018·ν. Figure 5 shows a representative fit and its 95% confidence interval for one of the 20 model systems that were calibrated. The average slope of all model systems was 0.021±0.003, indicating that bending of the leaf springs is slightly stiffer than theoretically determined. The averaged 95% confidence interval of all model systems comprised 8.7% of the generated force.

Representative force–displacement calibration curve. The solid line indicates the fit through the data points (squares) and the dotted lines indicate the 95% confidence intervals.
Tissue culture in the model system
TE constructs were successfully cultured in the model system for 4 weeks in both constrained and semi-constrained configuration (Fig. 6A–C). Newly formed tissue and compaction in width was clearly visible at week 4 (Fig. 6B, C) compared with day 0 (Fig. 6A). In Figure 6C it can be seen that the generated stress has displaced the sliding block attached to the leaf springs. The hematoxylin and eosin staining showed homogeneous tissue formation (Fig. 6D). The Masson-Trichrome and Picrosirius red staining indicate a dense collagen network with mature collagen fibers (Fig. 6D–F) after 4 weeks.

Tissue-engineered constructs just after seeding
Tissue compaction and generated stress during culture
The TE constructs started to compact after 2 weeks and continued to compact to approximately half of their original width (CW=51.2%±4.8%) at week 4 (Fig. 7A). At day 0, tissue thickness was 1000 μm and at week 4 a thickness of 1067 μm was assessed from histology. It was assumed that tissue thickness increase over time changed to a pattern similar to that of width decrease, starting after 2 weeks (Fig. 7B). Using this assumption, the total cross-sectional area of the TE constructs was determined, which compacted from 5.44±0.08 mm2 at start to 2.57±0.17 mm2 after 4 weeks (Fig. 7C). Force and stress generation showed the same time-lapse as compaction, starting after 2 weeks and gradually increasing up to, respectively, 20.3±2.9 mN and 8.0±1.3 kPa at week 4 (Fig. 8).
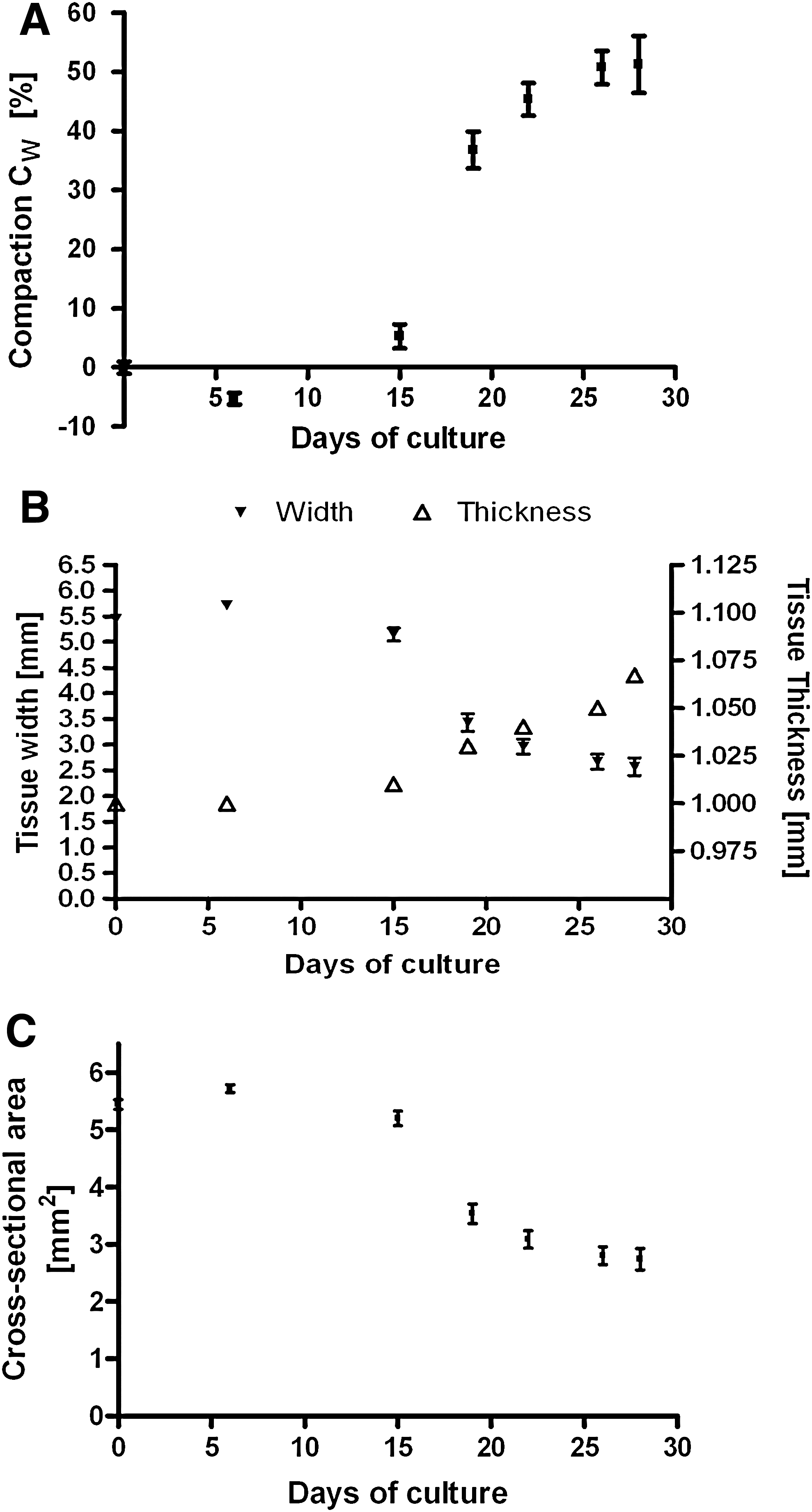
Tissue compaction

Force
Tissue retraction and generated stress after release of constraints
Within 5 s after release of constraints, the measured generated force and stress were, respectively, 8.7±0.5 mN and 3.3±0.3 kPa. After 30 min, generated force had gradually increased to 15.1±0.7 mN, which correlated to a generated stress of 5.6±0.6 kPa (Fig. 9A, B). Next, retraction was measured, which occurred very fast in the first 20 min after release of constraints. This resulted in a retraction of 16.4%±1.3%, which was almost half of the total retraction after 24 h being 35.8%±3.3% (Fig. 9C).

Generated force
Discussion
This article describes a new in vitro model system for investigating the development of stress, generated by traction forces exerted by the cells, in TE constructs. On the one hand, this cell traction is beneficial for tissue maturation and alignment in engineered tissues8,15,16; on the other hand, it causes tissue shrinkage at release of constraints (Fig. 1). Tissue shrinkage results from an imbalance between scaffold and tissue stiffness, and the traction forces of cells (Fig. 2). When traction forces exerted by the cells are larger than the forces the scaffold or newly formed tissue can resist, tissue shape will change. During tissue culture, scaffold and tissue properties change, influencing the effect of cell traction forces on tissue shape. In the present study, a model system is developed in which stress generation, compaction, and retraction can be measured in one single engineered heart valve tissue. Histology confirmed that the model system is suitable for tissue culture showing homogeneous tissue and abundant collagen formation after 4 weeks of culture.
Compaction and stress generation during culture
The TE constructs started to compact after 2 weeks of culture and continued to compact to approximately half of their original width at week 4. Generated force was quantified with an averaged error of 8.7% and divided by the cross-sectional area to obtain stress. The stress, generated by the cells, followed the same time-course as compaction reaching a value of 8.0 kPa at week 4. However, at week 4, compaction seemed to level off, whereas no signs of leveling off were observed in the stress.
Tissue compaction has been investigated extensively in all sorts of cell-populated constructs, predominantly made of collagen type I gels.17–20 Constrained culture of smooth muscle cells in collagen gels resulted in a compaction of 90% in 21 days, 19 whereas faster compaction of 98% in 6 days was observed in unconstrained disk gels. 18 Bone marrow stromal cells in constrained collagen lattices compacted 90% in 3 days time 17 and human dermal fibroblasts seeded on top of collagen gels reached a compaction in thickness of 85%, while being constrained to the surface. 20 Compaction of constrained tissues made from cell sheets of human dermal fibroblasts was less and slower, reaching a compaction of 70% after 21 days of constrained culture. 21 Contrary to our findings, compaction in most of these studies occurs within hours to days and the amount of compaction is higher, ranging from 50% up to 90%. These differences are probably caused by the type of scaffold used, its degradation profile, and the formation of new tissue. During the first 2 weeks, the traction forces of the cells are most likely counterbalanced by the stiffness of the scaffold. After 2 weeks, the scaffold rapidly loses its mechanical integrity 22 and the traction forces of the cells can compact the construct, which explains why first compaction was observed after 2 weeks. The fact that compaction does not reach extreme levels up to 90% as observed in collagen lattices implies that the developed ECM is less prone to the traction forces of the cells than a collagen gel is. It has been shown that increasing collagen concentration leads to decreased compaction.17,19 We believe that a fully developed matrix limits compaction as its collagen network is denser and stronger (due to cross-links) than the collagen network of a collagen gel.
Studies of human dermal fibroblasts in collagen lattices have shown that cells exert traction forces within hours to reach tensional homeostasis, and that these forces level off after 6–36 h.10,23,24 In the present study, 2 weeks were needed to generate stress and no plateau was reached at week 4. This phenomenon can also be explained by the degradation of the scaffold and the development of tissue over time. However, it is expected that if culture time is increased, the tissue further develops and equilibrium between traction forces and tissue stiffness would eventually be reached in the studied TE constructs as well.
Tissue stress and retraction after release of constraints
Retraction at release of constraints occurred very fast in the first 20 min, after which it gradually continued up to 36% after 24 h. The measured force also consisted of a fast and slow part similar to retraction. Immediately after release of constraints, a force of 8.7 mN was measured. After that, the measured force kept increasing and reached a value of 15.1 mN after 30 min. The fast processes of retraction at release of constraints are caused by the release of the developed stress in the ECM during culture and the release of stress in the cells. Next to this passive retraction, cells will also actively exert traction forces on their environment when their internal stress is released, seeking to reach their basal internal tension again. This process results in further retraction and measured force, but in a slower fashion. Although no new constraints are enforced to the tissue, retraction is hypothesized to stop eventually due to volume restrictions. Compared with fibroblast populated fibrin gels, retraction in the TE constructs was slow. Fibroblast-populated fibrin gels retracted 60% after release from the surface within 10 min, 11 even though cell density was 30 times lower than in the present experiment. This implies again that a fully developed ECM is better in withstanding traction forces of cells than a sole gel is.
Limits and future applications of the developed model system
Translation of force to stress required the tissue cross-sectional area. Width was measured in all samples but a histology section was used to estimate tissue thickness at week 4. A tissue shrinkage factor of 1.043 was assumed to correct for tissue dehydration during preparation and the thickness at earlier time points was determined by interpolation. A more accurate method should be developed for future experiments as both the tissue shrinkage factor and interpolation over time could be sources of errors. However, in this experiment changes in thickness appeared to be less than changes in width. Therefore, thickness changes had a minimal influence on the translation from force to stress.
The generated force and stress were slightly higher in the semi-constrained samples than in the constrained ones. In a fully constrained construct, cells will remodel their environment until an internal equilibrium in stress level is reached. In the semi-constrained samples, a certain degree of compaction (due to bending of the leaf springs) as well as generation of stress will occur as the cells exert traction forces to the tissue. It is hypothesized that this possibility of matrix remodeling in length leads to enhanced force and stress generation. Although the amount of compaction was only small in the semi-constrained samples, it does affect the position of the sliding blocks at which the cells reach their desired intracellular tension. The difference between constrained and semi-constrained can be reduced by increasing the stiffness of the leaf springs. However, increasing the stiffness of the leaf springs will reduce the displacement of the sliding block and therewith the accuracy of the force measurement.
The TE constructs are fabricated with seeding techniques, cells, and scaffold material similar to those of TE heart valves and therefore are a good representative of TE valves. The semi-constrained configuration resembles the situation within a leaflet more closely than the constrained configuration, as during valve culture leaflet flattening occurs due to cell traction. Retraction of the leaflets at release of constraints does differ from the TE constructs as it is counteracted by the constraints in the perpendicular direction. These differences should be investigated, that is, with computational models, when translating these results to TE heart valves.
The development of the currently described in vitro model system enables us to study and resolve the problem of retraction by fine-tuning the balance between scaffold, tissue, and cell traction. As shown by the time-course of stress development in this study, the scaffold is able to withstand compaction during the first 2 weeks. After that, tissue has to take over, but is not stiff enough yet. This suggests that either decreasing scaffold degradation (Fig. 10A) or increasing tissue stiffness (Fig. 10B) would change the course of compaction during culture and the resulting retraction. Other approaches could focus on reducing cell traction forces (Fig. 10C), by, for example, disruption of the actin fibers of the cells, disruption of cell–matrix interaction, or the use of a less contractile cell source (Fig. 10D). In addition to these problem-solving approaches, the developed model system is also a tool for further investigation of the role of cellular elements in compaction and retraction. Distinguishing between active and passive processes is required for understanding and controlling the process of retraction and is subject of future studies. Finally, in vivo, heart valves are subjected to continuous loading, which will counteract with the traction forces of the cells. This in vivo load could perhaps be used in finding the balance between cell traction and shape conservation. Now the force exerted by the cells is known, one can estimate the load needed to withstand retraction.
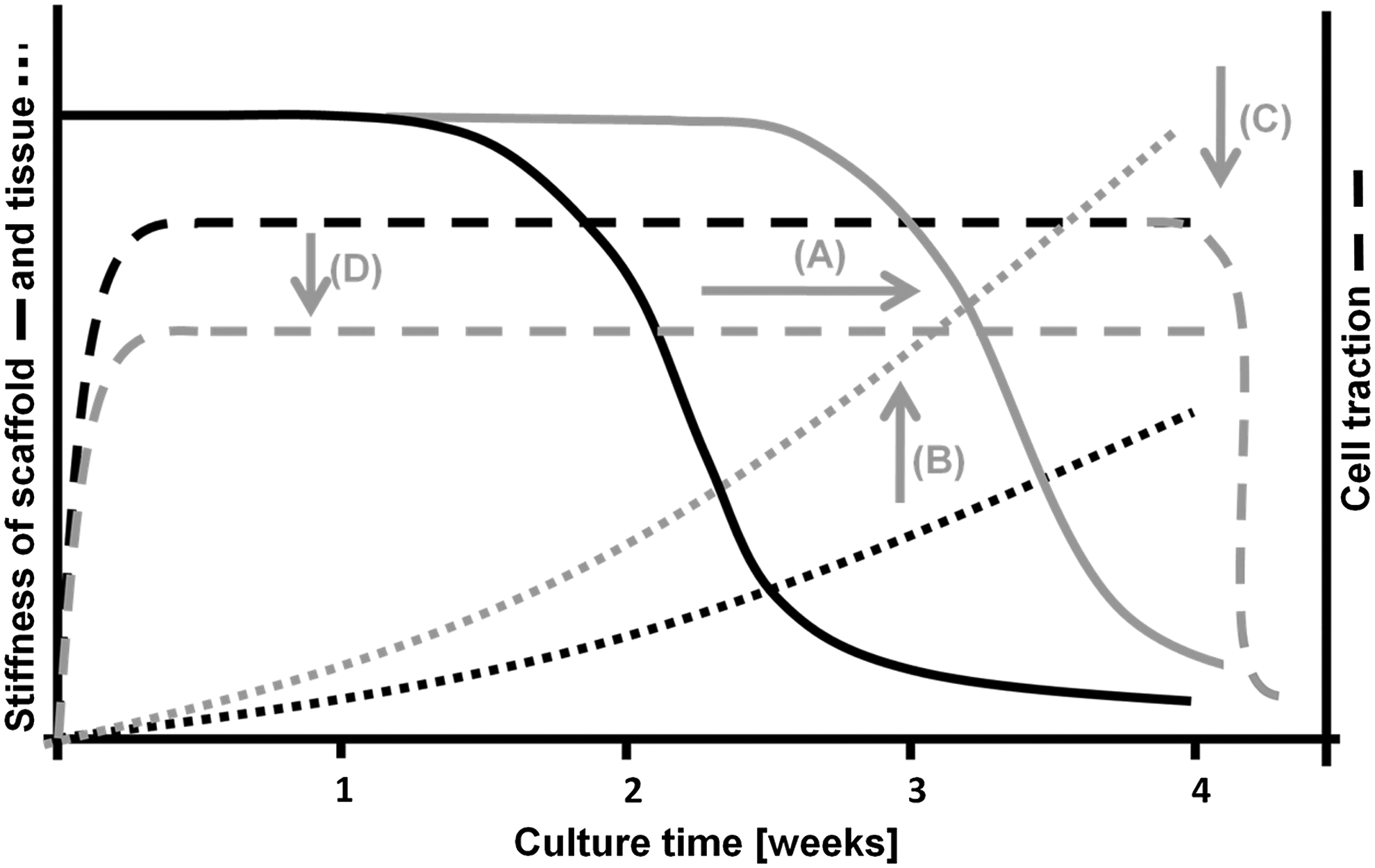
An overview of strategies (gray lines) to reduce tissue compaction and retraction. Decreasing scaffold degradation
In conclusion, with the developed model system we were able to quantify generated stress, compaction, and retraction over time and after release of constraints. In engineered heart valve tissue, both compaction and stress generation started after 2 weeks of culture and continued up to week 4. TE constructs compacted up to half of their original width and developed an internal stress of 6–8 kPa at week 4, which resulted in a retraction of 36%. The model system has provided a useful tool to unravel and optimize the balance between traction forces of the cells, tissue, and scaffold properties in engineered tissues to develop functional TE heart valve leaflets.
Footnotes
Acknowledgments
The authors gratefully acknowledge the support from the Smart Mix Program of The Netherlands Ministry of Economic Affairs and The Netherlands Ministry of Education, Culture and Science. The authors would like to thank Rob van den Berg for his contributions to the development of the model system.
Disclosure Statement
No competing financial interests exist.