
Abstract
A method to produce clinically useful platelets in vitro would help overcome the frequent shortages, donor deferrals, disease transmission, and alloimmunization with volunteer donor-derived platelets. Using CD34 positively selected cord blood cells, we investigated ways to increase platelet quality and yield in a three-dimensional modular perfusion bioreactor system. We found a two- to threefold increase in platelet numbers produced only when the early phases of the culture process were carried out at 5% oxygen, versus when 20% oxygen was used throughout the culture period (p<0.05), and much more than when 5% oxygen was used throughout. When the medium was routed through the cell–scaffold construct, versus when it flowed under and over the construct, or just intermittent feeding was used, the number of platelets increased two- to threefold (p<0.05), and enhanced collagen–induced aggregation. The 5% oxygen early in the culture process mimics the marrow adjacent to the bone where early progenitors proliferate. Flow through the cell–scaffold construct creates shear forces that mimic the flow in central venous sinuses of the marrow and enhances platelet production from proplatelets. The use of altered oxygen levels and cross flow enhanced platelet numbers and quality, and will contribute to eventual in vitro platelet production for clinical use.
Introduction
To date, there is no method of producing platelets in vitro in numbers that are clinically useful. Although a great deal of information has been obtained about platelet production in vivo, and it is possible to grow platelets in an in vitro system, it is not clear which steps are limiting production of platelets using an in vitro system. We have shown that we can expand cord blood hematopoietic cells in a three-dimensional (3D) scaffold to a much larger extent than with a two-dimensional (2D) control. 14 Production of platelets in a 3D scaffold and in a 3D bioreactor is a novel approach to modeling in vivo production that was not described before our recent preliminary article. 15 We showed that platelet production using a knitted polyester 3D scaffold was increased and prolonged versus using a 2D control, and is greatly increased and prolonged when the 3D cell–scaffold construct is supported in a modular perfusion bioreactor system. 15
The present article describes our work to increase platelet yield and improve platelet function in our 3D perfusion bioreactor system. To do this, we have experimented with changing oxygenation during the culture period and with changing flow-induced shear stress by modifying medium flow within the modular bioreactor system.
We explored decreased oxygen level during culture to enhance early progenitor self-renewal and hence eventual platelet production. Our reasoning like that of others was that there is a lower level of oxygen in the marrow space near the surrounding bone where early hematopoietic cells are found.16–18 We compared 20% O2 versus 5% O2 during different stages of our 3-stage platelet production method.
We postulated that instituting continuous flow across the cell–scaffold construct during incubation in the bioreactor would increase platelet production. We based this expectation on descriptions in the literature of up to a 20-fold increase in platelet production in a 2D flow system, presumably by promoting platelet release from proplatelets.19,20 We utilized the modular 3D perfusion bioreactor to compare 3 modes of medium change and platelet harvest to test this hypothesis.
Methods
Cord blood cell isolation and CD34 positive cell separation
Umbilical cord blood units were obtained from the Cleveland Cord Blood Center, Cleveland, Ohio, from normal full-term deliveries after institutional review board approval and informed consent. Light-density cells were isolated from citrated cord blood using discontinuous density centrifugation over Ficoll-Paque Plus (GE Healthcare BioSciences). CD34-positive cell selection was conducted using MACS Direct CD34 Progenitor Cell Isolation Kits (Miltenyi Biotec). Each experiment was performed using a unit of cord blood; ∼2 million CD34 positively selected cells were used for each of the conditions in each experiment.
3D scaffold and bioreactor
We used a modular 3D perfusion bioreactor system that we developed and reported previously (Fig. 2A). 15 Knitted polyester surgical fabric (Bard; C.R. Bard Inc.) served as the 3D scaffold. The bioreactor modules are designed to allow medium to flow over and under and/or through the cell–scaffold construct, with collection of nonadherent cells in the lower medium space from the expelled medium. Gas is exchanged with the medium gas spaces above and below the medium spaces. The gas spaces are separated from the medium compartments by gas-permeable smooth-finish fluorinated ethylene propylene membranes (McMaster Carr). Racetrack-shaped pieces of the fabric, 48 mm in length, 12 mm in width, and 1 mm in depth, were inserted into the bioreactor modules.
Cell culture was carried out in three stages. In Stage 1, CD34 positively selected cord blood cells were put into liquid culture in T75 tissue culture flasks at 100,000 cells per mL in 10–15 mL of medium initially, and then divided into additional T75 tissue culture flasks during the next 48 to 72 h as the cells proliferated for a final concentration of one million cells per mL. Cell expansion was carried out in a humidified tissue culture incubator at 37°C with 5% CO2 in either air or air mixed with nitrogen to achieve 5% oxygen. The medium consisted of Iscove's modified Dulbecco's medium (IMDM; Gibco, Invitrogen) expansion medium containing 10% fetal calf serum (FCS; JRH Bio Sciences), stem cell factor (SCF, 50 ng/mL), flt-3 ligand (FL, 10 ng/mL), thrombopoietin (TPO, 10 ng/mL), and 10 ng/mL interleukin (IL)-6. All growth factors were from R & D Systems.
After this liquid expansion phase, the cells were infused into the upper chamber of the bioreactor modules, with flow into the bottom medium chamber and flow out of the top medium chamber blocked, so that the cell–medium mixture circulated through rather than parallel to the surface of the scaffold material (Fig. 2A); this resulted in trapping of cells within the scaffold. 21 Then, Stage 2 was accomplished by incubation for 7 days at 37°C using medium consisting of IMDM, 10% FCS, 10% horse serum (JRH Bio Sciences), 0.25 μM hydrocortisone, 10 ng/mL TPO, and 25 ng/mL FL. All additional medium additives and growth factors were from R & D Systems.
The third stage, differentiation, followed. After 7 days in the bioreactor modules, the medium was changed to an Src differentiation medium that consisted of IMDM, 10% FBS, 30 ng/mL TPO, 1 ng/mL SCF, 7.5 ng/mL IL-6, 13.5 ng/mL IL-9, and 2.5 μM SU6656 (Sigma), an Src kinase inhibitor. 22 On day 14, the medium was switched to IMDM with 10% FBS, 1% bovine serum albumin, 25 ng/mL TPO, 25 ng/mL IL-3, 50 ng/mL IL-6, and 10 ng IL-11.
For Stages 2 and 3, performed in the bioreactor system, 3 mL of medium (the approximate void volume of each bioreactor module) was changed daily by closing the bottom medium entrance port and top exit port and forcing fresh medium through the cell–scaffold construct and allowing collection of the expelled medium and nonadherent cells through the bottom exit port. Each bioreactor module has a separate seeding port allowing each module to be manipulated or removed independently of the other modules despite connections through the tubing network.
Hypoxia experiments
To determine the effect of reduced oxygen levels, a portion or all of the three stage process was carried out at either 5% oxygen or 20% oxygen. All oxygen-regulated experiments were performed in a Thermo Scientific Model 3130 water-jacketed CO2 incubator maintained at 37°C using compressed nitrogen to control oxygen content. Three sets of conditions were compared. Then, 20% oxygen was used for all three stages, 5% oxygen was used for stages 1 and 2, and 20% oxygen for stage 3, or all 3 stages were carried out in 5% oxygen.
Medium flow variations (using hypoxia in stages 1 and 2)
To measure the effect of constant flow-induced shear on platelet production, we compared flow through the cell–scaffold construct (cross flow), achieved by blocking the lower medium input port and the upper output port, with flow parallel to the cell–scaffold construct (parallel flow; all ports open; Fig. 2). Medium passed through the module continuously at 3 mL/24 h. A 3 mL bolus was used for harvest in both the cross and parallel flow modes, and for the intermittent flow bolus alone method used as a control. 15
Platelet collection and analysis
Cells and medium were collected continuously in Fenwal PL732 plastic storage bags (Baxter Healthcare) designed for platelet storage. In addition, daily medium exchange and cell harvests were accomplished simultaneously; medium (3 mL) containing nonadherent cells was withdrawn from the bioreactor as fresh medium entered the module from the medium supply bottle. For platelet function studies, a syringe containing ∼30 mL glucose-based platelet storage medium 23 was attached to the harvest port of the bioreactor module and the 3 mL of expelled medium and cells were collected into the module. For experimental controls, neonatal platelets were isolated from citrated cord blood by 120 g centrifugation for 15 min at room temperature. Neonatal platelets were then washed and labeled as described for experimentally produced platelets. Platelet concentrates from the local American Red Cross were also used as controls.
Platelet aggregation
To determine the functional properties of platelets harvested from 3D scaffolds in wells and from 3D perfusion bioreactor growth conditions, harvested platelets were collected and washed in glucose storage buffer. The platelets were then centrifuged at 1000 g and resuspended in human plasma to create a platelet-rich concentrate at a minimum concentration of 10 million platelets per mL. The platelets were then analyzed using an AggRam aggregometer (Helena Laboratories). Aggregation was studied after addition of an agonist. Agonists used included collagen at a final concentration of 10 μg/mL, ristocetin at 1.5 mg/mL, ADP at 20 μM, and epinephrine at 200 μM.
Flow cytometry
Cells were washed in Ca++/Mg++ free Dulbecco's PBS to remove culture medium and labeled with FITC- or phycoerythrin-conjugated antibodies (Immunotech) for surface antigens, including those for CD34, CD41, CD62, and CD63. CD41 is part of the GPIIb/IIIa complex on the surface of megakaryocytes and platelets. CD62 (p-selectin) is a constituent of alpha granules that moves to the surface of platelets with activation. 24 CD63 (granulophysin), found in dense granules, is also moved to the surface with platelet activation. Excess antibody was removed by washing, and the fluorescent cell analysis was performed on a BD FACSCalibur System flow cytometer (BD Biosciences). Isotype-labeled and unstained cells were used as controls. CD62 and CD63 analysis was done before and after addition of thrombin at 1 U/mL for ten minutes at 37°C.
Data analysis
Flow cytometry populations and comparisons between them were analyzed with the Kolmogorov-Smirnov algorithm using FlowJo flow cytometry analysis software Version 7.6.3 (Tree Star, Inc.). Paired t-tests were calculated using Microsoft Excel (Microsoft).
Results
Reduced oxygen experiments
Using flow cytometry following the first stage liquid culture, we studied cells incubated at 5% versus 20% oxygen (Table 1). We found that the lower oxygen tension increased the number of CD34-positive cells, as well as the early hematopoietic cells that were CD34 positive lineage negative and CD34 positive CD38 negative. There was also an increased proportion and number of CD34-positive CD41-positive cells that include committed megakaryocytic lineage progenitors.
We found that if the entire platelet production scheme outlined above was performed at 5% O2, there was a marked diminution in platelet production, compared to using 20% oxygen throughout the incubation. On the other hand, if Stages 1 and 2 (initial 2–3 day expansion in liquid culture and 7 day expansion in the 3D scaffold) were performed at 5% and the differentiation stage (Stage 3) was carried out at 20% O2, we found an approximate doubling of the number of platelets produced in the 3D scaffolds in wells and in the 3D bioreactor system (Table 2 and Fig. 1).
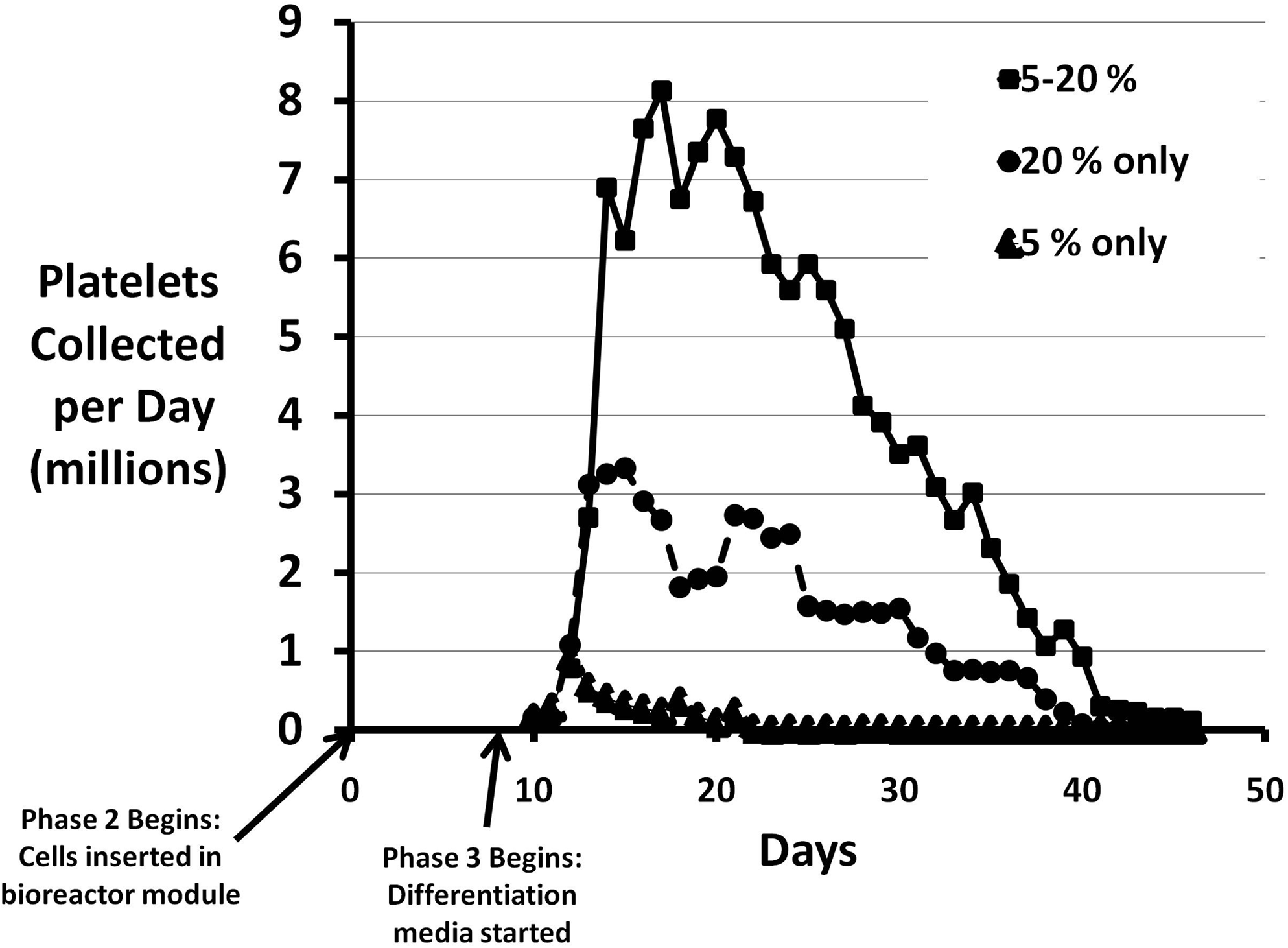
Effect of low oxygen tension on platelet production in the bioreactor system. The solid squares represent daily platelet collection numbers when 5% oxygen was used through day 10, followed by 20% oxygen thereafter. The solid circles show the effect of 20% oxygen throughout the culture period. The triangles indicate daily platelet numbers when 5% oxygen was used throughout the culture period. Time zero refers to the point at which the cells are added to the scaffold in the bioreactor modules, the beginning of stage 2.
ND, not done, NS, not significant.
Based on the results from the hypoxic condition studies, we routinely incorporated the use of 5% oxygen concentration for the first two phases of platelet production followed by 20% oxygen during the differentiation phase.
Cross flow experiments
In the modular perfusion bioreactor system, we next compared intermittent bolus feeding (rapid injection of 3 mL new medium every 24 h with collection of cells and platelets in the expelled medium) with the addition of continuous flow of 3 mL over 24 h. This additional medium was directed either in parallel with the surface of the cell–scaffold construct (parallel flow) or through the cell–scaffold construct from the top of the module to the bottom (cross flow) (Fig. 2A). We found up to a threefold increase in platelet production, from day 17 onward, with cross flow, compared to intermittent bolus feeding alone (Table 2, Fig. 2B). We found no significant effect of using continuous parallel flow (medium along the surface but not through the cell–scaffold construct) on platelet production, compared to intermittent bolus feeding alone.

Using flow cytometry, we found that platelets made using cross flow had far fewer microparticles than platelets made using intermittent bolus feeding alone. In Cross Flow Experiment 1 (Table 2), forward scatter analysis indicated that, for platelets collected early in the differentiation phase, cross flow culture produced 4.0% microparticles versus 10.6% for bolus feeding only. For platelets collected late in the differentiation phase, cross flow produced 3.3% microparticles, whereas bolus feeding produced 17% microparticles.
While we were previously able to demonstrate platelet aggregation in response to thrombin with platelets from intermittent bolus feeding alone, we had been unable to demonstrate aggregation to other agonists. With cross flow, we found that the platelets produced aggregated well to collagen (Fig. 3A), but not to ristocetin, ADP, or epinephrine. For comparison purposes in this experiment we used donor platelets collected and processed by the local Red Cross blood center and stored for 2 days. These platelets aggregated in response to collagen and ristocetin, but not ADP or epinephrine.
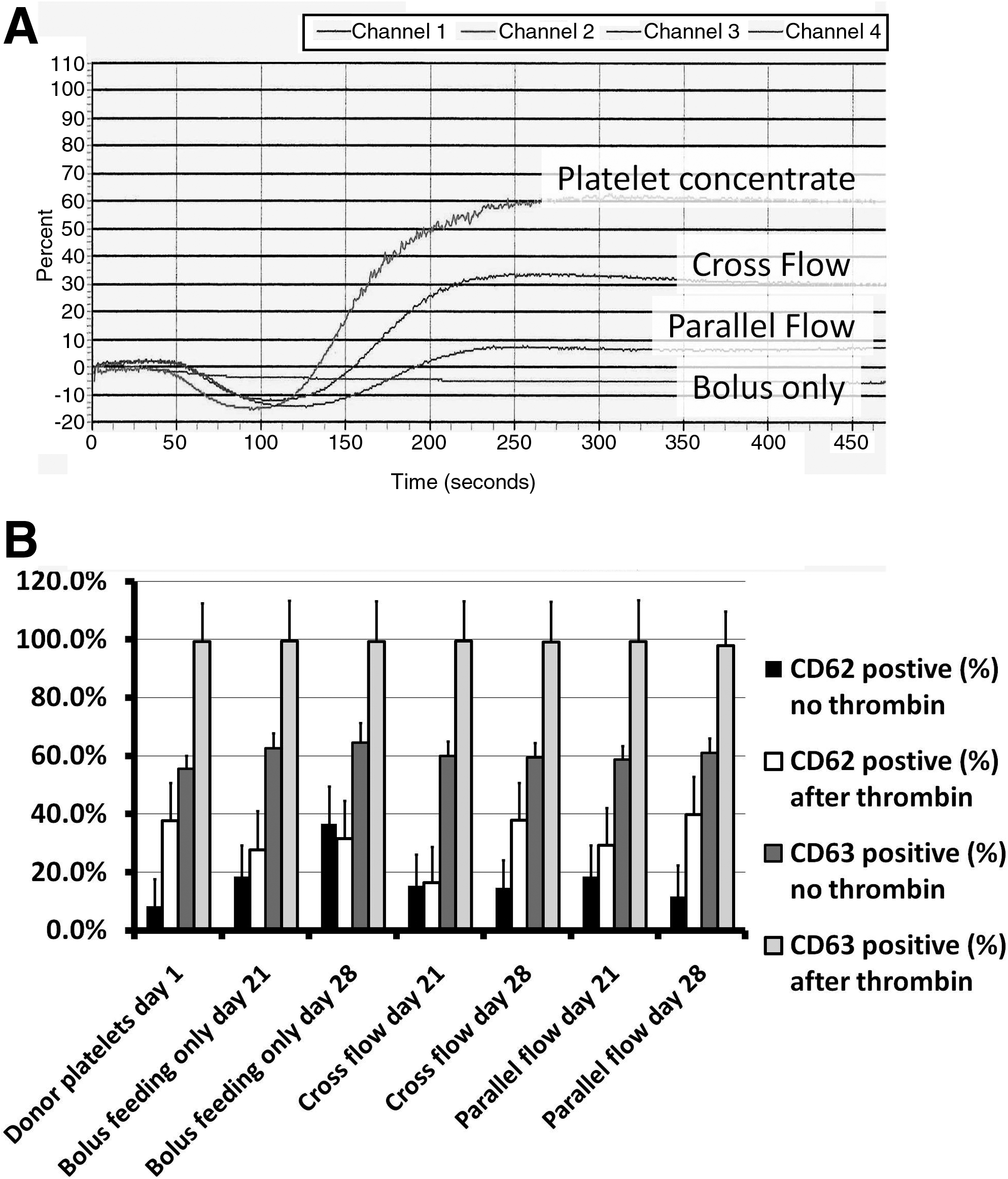
We examined expression of CD62 and CD63 before and after added thrombin to evaluate the platelets for activation as harvested and after stimulation (Fig. 3B). In this typical experiment, we found that cross flow at day 21 and 28 and parallel flow at day 28 had relatively less CD62 activation at harvest than with bolus feeding/harvest alone (not statistically significant [NS] for day 21 and p<0.001 for day 28). Cross flow and parallel flow at day 28 had more activation in response to thrombin than bolus feeding/harvest alone (NS). Parallel flow had even lower activation at harvest and higher CD62 expression after thrombin than cross flow at day 28 (NS before thrombin, p<0.001 after). At days 21 and 28 CD63 activation was somewhat but not statistically significantly higher at harvest for bolus feeding/harvest alone, but all conditions produced platelets that responded with high CD63 positivity on exposure to thrombin (no statistically significant differences).
Discussion
The results presented here are an important step in development of an ex vivo system to produce platelets for clinical use. The two manipulations described are based on the known physiology of the growth and production of hematopoietic cells and platelets in vivo, and each results in a several-fold increase in the number of platelets produced, compared to controls. The quality of the platelets, as measured by their aggregation response to collagen, their lower baseline activation, and their marked activation after stimulation with low dose thrombin, is also increased.
Although no one has yet reported in vitro production of platelets in numbers near that required for transfusion in humans, some beginning work has been done. Including some mentioned above, there have been a number of published studies of megakaryocyte and/or platelet production from several starting cell populations, including human cord blood; murine, human, and nonhuman primate embryonic stem cells; cancer cell lines; peripheral blood progenitor cells; and marrow, both human and murine.22,25–36 Many of these described production of megakaryocytes but not platelets. Some described production of platelets qualitatively but not quantitatively. Most were performed in suspension culture or on a 2D surface; some used feeder layers. A wide variety of added growth factors or, for earlier studies, conditioned medium was used to support the cultures. Two studies have reported formation of platelets in NOD/SCID mice after transplant with hematopoietic progenitors grown from human embryonic stem cells on OP9 feeder layers in wells.33,37 Matsunaga et al. reported platelet production using a three-stage approach in tissue culture flasks that used genetically modified human marrow stroma (carrying an inserted gene for telomerase) as a feeder layer. 25 The platelets functioned as verified using aggregation and P-selectin (CD62) expression on stimulation with ADP and fibrinogen, among other tests. When they gel-filtered to remove microparticles, they recovered 60% to 80% of the platelets. Their experiments used very small batch sizes; they noted that there were problems scaling up to larger experiments to produce clinically relevant numbers of platelets. None of these reports described using 3D scaffolds.
In the current study, we showed that the platelets produced aggregated in response to collagen but not to risticetin, ADP, or epinephrine. Conventionally collected and stored platelet concentrate aggregated to both collagen and ristocetin but not epinephrine or ADP. This is a very relevant control, since this is the currently used source of platelets for transfusion. The aggregation response to these four agonists in stored platelet concentrates has been variously reported in the literature, however. For example, Miyaji et al. found very low ADP-induced aggregation but much higher collagen-induced aggregation in stored platelet concentrates; the ADP-induced aggregation recovered after infusion into thrombocytopenic patients. 38 Apelseth et al. found no aggregation in stored platelets with ADP, and gradual decrease in response to collagen over 5 days of storage. 39 Kocazeybek et al. reported aggregation in stored platelet concentrates to ADP, ristocetin, collagen, and epinephrine, but a less rapid fall-off in response to collagen versus the other agonists over 5 days of storage. 40
We found an increase in CD34-positive cells and in early hematopoietic CD34 positive subsets, namely, lineage negative and CD38 positive, with liquid culture at 5% versus 20% oxygen. Committed megakaryocytic progenitor cells characterized by dual positivity for CD34 and CD41 were also increased. Enhancement of in vitro hematopoiesis incorporating reduced oxygen levels has been known for some time. The marrow space near bone is believed to have a low oxygen level. 16 Haylock et al. have shown that the earliest hematopoietic cells lie adjacent to the marrow endosteum in mice. 17 Hypoxia seems to decrease early hematopoietic cell-cycle entry and to promote hematopoietic cell self-renewal over differentiation when cell cycling does occur. 41 It may work through hypoxia-inducible factor 1 (HIF-1), a heterodimeric transcription factor stabilized in hypoxic conditions. HIF-1 in turn regulates many genes involved in metabolism, cell-cycle regulation, erythropoiesis, and angiogenesis.
Interestingly, extension of the induced hypoxia into the differentiation phase of platelet production markedly decreased platelet output. This would appear to contradict the speculation of Wasiluk et al. that the thrombocytopenia of small-for-gestation-age newborns is due to the effect of increased erythropoietin production observed in these children; our data suggest that the hypoxia per se without high erythropoietin levels might be responsible. 42 Mostafa et al. found that 5% oxygen decreased megakaryocyte production and ploidy compared to that observed at 20% oxygen, results that are reflected in our observations when hypoxia was used for both early and late hematopoiesis and megakaryocytopoiesis. 18
In the work presented here, we have chosen to use 5% oxygen to promote early cell self-renewal while limiting the cell-cycle quiescence in hematopoietic stem cells associated with lower oxygen levels immediately adjacent to the bone. In this application, it may not be necessary to expand the earliest hematopoietic stem cells; expansion of more committed but still early progenitors may suffice.
The prototypic 3D perfusion bioreactor modules we have developed facilitate medium and gas exchange over and under or through a cell support scaffold. 15 The design allows maturing cells with altered adherence characteristics to fall through the cell–scaffold construct to be collected in effluent medium. In our previous work, we harvested platelets from the bioreactor modules by running new medium into the top of the bioreactor modules and taking it out of the bottom (at the opposite end) at the same time. This caused a cross flow of medium through the cell–scaffold constructs. We believe that the resulting shear dislodges platelets that have not yet fallen into the bottom chamber and may increase platelet release from proplatelets, accounting for at least some of the increase in platelet output when the 3D scaffold is used in the bioreactor system versus in wells. Increased release of platelets in flowing blood or medium has been documented by others both in vivo and in vitro.19,20 On the other hand, shear has been shown to activate platelets in the circulation, especially with the elongational shear forces that also are believed to enhance platelet release from proplatelets.43,44 We also hypothesized that increased cross flow would decrease platelet dwell time in the cell–scaffold construct, perhaps balancing the activation effects of increased shear to some extent. Our results supported this hypothesis: we were able to increase the number of putative platelets produced several fold, and we improved function as measured by aggregation in response to collagen and activation of the platelets as collected from the device. The lower baseline activation with parallel flow versus cross flow was presumably because flow past the surfaces of the cell–scaffold construct was less traumatic than flow through the construct.
Footnotes
Acknowledgments
This work was supported by the Center for Stem Cell and Regenerative Medicine (Tech 06-063) through the Biomedical Research and Commercialization Program, a component of the Third Frontier Program of Ohio. Flow cytometry was performed at the University Cell Analysis and Sorting Core, a joint venture between The Comprehensive Cancer Center and The Davis Heart & Lung Research Institute, both at The Ohio State University. The authors would like to thank Helena Laboratories for providing the aggregometer and associated reagents used in this study.
Disclosure Statement
Neither author has commercial associations that might create a conflict of interest in connection with this article.