
Abstract
In cartilage tissue engineering an optimized culture system, maintaining an appropriate extracellular environment (e.g., pH of media), can increase cell proliferation and extracellular matrix (ECM) accumulation. We have previously reported on a continuous-flow bioreactor that improves tissue growth by supplying the cells with a near infinite supply of medium. Previous studies have observed that acidic environments reduce ECM synthesis and chondrocyte proliferation. Hence, in this study we investigated the combined effects of a continuous culture system (bioreactor) together with additional buffering agents (e.g., sodium bicarbonate [NaHCO3]) on cartilaginous tissue growth in vitro. Isolated bovine chondrocytes were grown in three-dimensional cultures, either in static conditions or in a continuous-flow bioreactor, in media with or without NaHCO3. Tissue constructs cultivated in the bioreactor with NaHCO3-supplemented media were characterized with significantly increased (p<0.05) ECM accumulation (glycosaminoglycans a 98-fold increase; collagen a 25-fold increase) and a 13-fold increase in cell proliferation, in comparison with static cultures. Additionally, constructs grown in the bioreactor with NaHCO3-supplemented media were significantly thicker than all other constructs (p<0.05). Further, the chondrocytes from the primary construct expanded and synthesized ECM, forming a secondary construct without a separate expansion phase, with a diameter and thickness of 4 mm and 0.72 mm respectively. Tissue outgrowth was negligible in all other culturing conditions. Thus this study demonstrates the advantage of employing a continuous flow bioreactor coupled with NaHCO3 supplemented media for articular cartilage tissue engineering.
Introduction
One important factor which could be better controlled in continuous flow than in batch bioreactors, particularly for cartilage tissue engineering, is extracellular pH. Chondrocytes rely virtually entirely on glycolysis as a source of energy and hence use glucose and produce lactic acid at a high rate.9,10 Lactic acid if not removed can rapidly acidify the culture medium.8,11,12 It has long been known that chondrocyte matrix synthesis rates and rates of cell proliferation are very sensitive to extracellular pH and steeply fall in a pH-dependent manner under acidic conditions.13–20 It has been shown by Ohshima and Urban 14 and Wilkins and Hall 18 that, in regard to extracellular pH, matrix synthesis follows a bell shaped curve with optimum matrix synthesis occurring between extracellular pH 6.9 and 7.1. Acidic extracellular environments influence the intracellular pH (pHi) of chondrocytes.17,18 The pHi of articular chondrocytes is mainly regulated by the amiloride-sensitive Na+/H+ exchanger;17,21 however the pHi of chondrocytes within the superficial zone of bovine cartilage is exclusively regulated by HCO3− dependent transporter(s). 22 Bovine articular chondrocytes have a resting pHi of pH 7.1 but can only maintain this level of pHi over a narrow range of external pH values. Since changes to pHi affect ECM synthesis, 17 efficient buffering systems are important if pHi is to remain at desirable levels. It is therefore surprising that little attention has been paid to buffering systems and maintenance of an optimal extracellular pH in studies of bioreactors; indeed in many cartilage bioreactor studies the type of buffering system employed is not even reported in the methods.3,23–28 Thus the effect of changes in extracellular pH on matrix synthesis during chondrocyte culture in bioreactors is still unknown.
In our previous study, we found that chondrocytes cultured in a continuous flow bioreactor considerably accumulated more matrix than those cultured under static conditions when both systems were buffered only with 20 mM HEPES. 8 Hence, the objective of this study was to determine whether the use of sodium bicarbonate (NaHCO3) as an additional buffering agent would help maintain an extracellular pH between 6.9 and 7.2 in the bioreactor, and thus further enhance ECM accumulation.
NaHCO3 was added to the media as a second buffering agent for three distinct reasons: (i) bicarbonate is a physiological buffer in the body29–32 ; (ii) NaHCO3 has the capacity to effectively neutralize acids by the carbonic acid (H2CO3) dissociation process (HCO3−+H+→H2CO3→CO2+H2O)29,31,33; (iii) bicarbonate ions (HCO3−), can be actively transported into the cell to buffer the intracellular space22,29 and increase ECM synthesis. So addition of NaHCO3 to the culturing media is an effective way to maintain extracellular and hence pHi that in turn helps stimulate ECM synthesis.15,20,34,35 It is important to note that bicarbonate is a base and the presence of another nonbicarbonate buffering agent (e.g., HEPES) is essential.30,33,34
The media used in this study was Ham's F-12 which was further supplemented with a 14 mM concentration of NaHCO3. The addition of 14 mM NaHCO3 was based on a study conducted in 1965 by Ham. 36 Additionally, this concentration of NaHCO3 (14 mM) with Ham's F-12 media is recommended by Sigma Aldrich and Gibco, for cell culture under 5% CO2.30,34,36,37 As addition of bicarbonate also increased the osmolarity of the medium, which could itself perhaps influence cellular activity, we also investigated whether bicarbonate buffering or change in osmolarity was responsible for any changes seen.
Material and Methods
Cell isolation and high-density three-dimensional culture (static and continuous flow)
Cartilaginous tissue constructs were generated using isolated chondrocytes harvested from calf (12–18 months old) metacarpal-phalangeal articular cartilage by sequential enzymatic digestion, as previously described. 38 Tissue was obtained from several joints (up to four per experiment) and pooled together to get a sufficient cell number. The cells were seeded on the surface of type II collagen-coated Millicell™ filters in high-density, three-dimensional culture (2×106 cells/filter or 33,000 cells/mm2) and maintained in Ham's F-12 media (HyClone) containing 10 mM glucose which was supplemented with 20% fetal bovine serum, 100 mg/mL ascorbate. Ham's F-12 medium without added bicarbonate, but supplemented with 20 mM HEPES (N-2-hydroxyethylpiperazine-N′-ethanesulfonic acid; Sigma Aldrich Ltd.), had a pH of 7.4 in air and a pH of 6.95 under 5% CO2. The cultures were grown in an incubator maintained at 37°C and 95% relative humidity supplied with 5% CO2: 95% atmospheric air. The culture medium (1 mL per filter) was changed every 48–72 h and fresh ascorbic acid was added with each change. Cultures were grown under static (no-flow) conditions for a week to generate a layer of cartilaginous tissue before being cultured in the continuous flow bioreactor, as previously described. 8 In the bioreactor, cultures were supplied with a 10 μL/min continuous flow of medium with or without a supplement of 14 mM NaHCO3. The 10 μL/min flow rate corresponded to an average residence time of ∼7 h. Additionally, cultures were also grown under static conditions in medium with or without supplementary 14 mM NaHCO3. The above-mentioned culturing conditions were repeated at least 3 times using bovine chondrocytes isolated at various time points and all the cultures were grown for 5 weeks.
Osmolarity control experiments were also conducted in the reactor as described above for a period of 3 weeks. In these experiments the osmolarity of the media without NaHCO3 was adjusted to the same osmolarity as the media containing 14 mM NaHCO3 by the addition of ∼24 mM mannitol. After the addition of mannitol the osmolarity of both the solutions was remeasured to confirm that it was the same.
Determination of ECM accumulation and cellularity
At the end of the culture periods (static and continuous), tissue constructs were harvested, lightly patted dry, and weighed (wet weight). Tissues were lyophilized overnight and the dry weight of the tissue determined. Tissue samples were then digested with papain (80 μg/mL in 20 mM ammonia acetate, 1 mM ethylenediaminetetraacetic acid and 2 mM dithiothreitol [DTT]) for 72 h at 65°C and stored at −20°C until analysis. Aliquots of the digest were analyzed for glycosaminoglycans (GAGs), collagen, and DNA content. Sulfated GAGs were quantified by using the dimethylmethylene blue dye binding assay (Polysciences Inc.).39,40 Collagen content was estimated by assuming that hydroxyproline comprises ∼10% the weight of collagen. 41 Aliquots of the papain digest were hydrolyzed in 6 N HCl at 110°C for 18 h. Hydroxyproline content of the hydrolyzate was determined using chloramine-T/Ehrlich's reagent assay. 42 DNA content was determined using the Hoechst dye 33258 assay (Sigma Aldrich Ltd.). 43
Histological and immunohistochemical evaluation
Cultures from each of the culturing conditions were subjected to histological and immunohistochemical evaluation. Upon harvest, tissue constructs were fixed overnight in 4% paraformaldehyde (Sigma Aldrich Ltd.) and embedded in paraffin. Thin sections (5 μm thick) were cut and stained with hemotoxylin-eosin (H&E) and toluidine blue (TB). All sections were examined by light microscopy. Cryofixed sections were immunohistochemically tested with antibodies against type I and II collagen. 44 Before incubation with the primary antibody, sections were predigested with 0.25% trypsin (Sigma Aldrich Ltd.) only for collagen II and 0.5 units/mL chondroitinase ABC and 0.5% hyaluronidase (Sigma Aldrich Ltd.) to facilitate immunostaining. The following primary antibodies and dilutions were used: rabbit polyclonal type I collagen antibody (Fitzgerald), 1:200 dilution; mouse monoclonal type II collagen antibody (Developmental Studies Hybridoma Bank, University of Iowa, IA), 1:10 dilution. Fluorescently labeled secondary antibodies were used, with 2% (v/v) fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit Fab fragment (Vector Laboratories) for collagen I and FITC-conjugated horse anti-mouse Fab fragment (Vector Laboratories) for collagen II. Sections were washed and mounted with Vectashield hardest mounting medium with 4′,6 diamidino-2-phenylindole (DAPI) (Vector Laboratories).
Tissue thickness measurements
The thicknesses of the tissue constructs were determined via two methods: (i) bright-field images of the histology sections and (ii) needle probe method as reported by Hoch et al. 45 Bright-field images of histological sections for the various culturing conditions were calibrated with an online scale bar, from which the thicknesses of the tissue samples were determined; thickness measurements were taken at nine random locations and the average value recorded. In the needle probe method a 25-gauge needle (Becton-Dickinson) was attached to a 1 kg load cell of a Mach-1 mechanical tester (Biosyntech) which was then displaced into the tissue at a rate of 5 μm/s. Resistive compressive forces and needle displacement were recorded at a frequency of 10 Hz. Abrupt changes in force were interpreted as needle contact with the tissue and the underlying support surface to determine the tissue thickness. Thickness measurements were taken at nine random locations and the average value was recorded.
Indentation testing
Elastic moduli of the developed tissue constructs were determined by indentation testing. Harvested tissues were placed flush on the surface of a porous steel stage of a Mach-1™ micromechanical tester (Biosyntech) equipped with a 1 kg load cell and a plane-ended indenter (2.1 mm diameter). Samples were preloaded to 10 mN and then subjected to step indentations of 2% strain to a maximum of 20% strain. At each step, the resulting force decay was recorded until equilibrium was reached (≤2 mN/min). The elastic modulus at 20% strain was then determined using the expression derived by Hayes et al. 46 for indentation testing of cartilage.
Statistical analyses
All numerical results were expressed as the mean±standard error of the mean (SEM). Three independent experiments were conducted with 3 independent cell extractions. Data was compared between each of the experimental groups using a two-way analysis of variance and the Fisher's least significant difference (LSD) post hoc test (SPSS 18.0; SPSS Inc.). Significance was associated with p-values <0.05.
Results
Combined effect of NaHCO3 and continuous flow bioreactor on ECM accumulation and cellularity
To determine the combined effects of NaHCO3 and a continuous flow of medium on cartilaginous tissue growth, tissue constructs after 1 week of preculture were cultivated in a continuous-flow bioreactor with a medium flowrate of 10 μL/min. The medium used in the bioreactor was supplemented with 14 mM NaHCO3, and the constructs were cultured for a duration of 5 weeks (Fig. 1). Tissue constructs cultivated in the continuous flow bioreactor with medium supplemented with 14 mM NaHCO3 accumulated significantly more (p<0.05) ECM in comparison with all other conditions (Fig. 2). Thus, (i) dry weight of tissue: reactor with NaHCO3: 20.5±0.98 mg, reactor without NaHCO3: 3.7±0.13 mg, static with NaHCO3: 2.9±0.22 mg, and static without NaHCO3: 2.1±0.1 mg (p<0.05; n=9), (ii) wet weight of tissue: reactor with NaHCO3: 292.4±16.3 mg, reactor without NaHCO3: 33.2±1.2 mg, static with NaHCO3: 23.1±1.4 mg, and static without NaHCO3: 12.8±1.0 mg (p<0.05; n=9). For the GAG and collagen results, see Figure 3. Additionally, the tissue constructs cultured in the bioreactor with NaHCO3-supplemented medium demonstrated a significant increase in cellularity in comparison with all other culturing conditions. Thus in the reactor, DNA was increased by 6-fold when the medium contained supplementary bicarbonate, while in the static (no flow) controls, the corresponding increase in DNA was 13-fold (p<0.05; Fig. 4). Although, there was a significant increase (p<0.05) in ECM macromolecules accumulated per microgram of DNA in NaHCO3-supplemented static cultures (no flow) in comparison with static controls (no flow) without NaHCO3 supplementation, no significant difference was observed between the two continuous-flow bioreactor conditions (i.e., with NaHCO3, or without NaHCO3; Table 1).

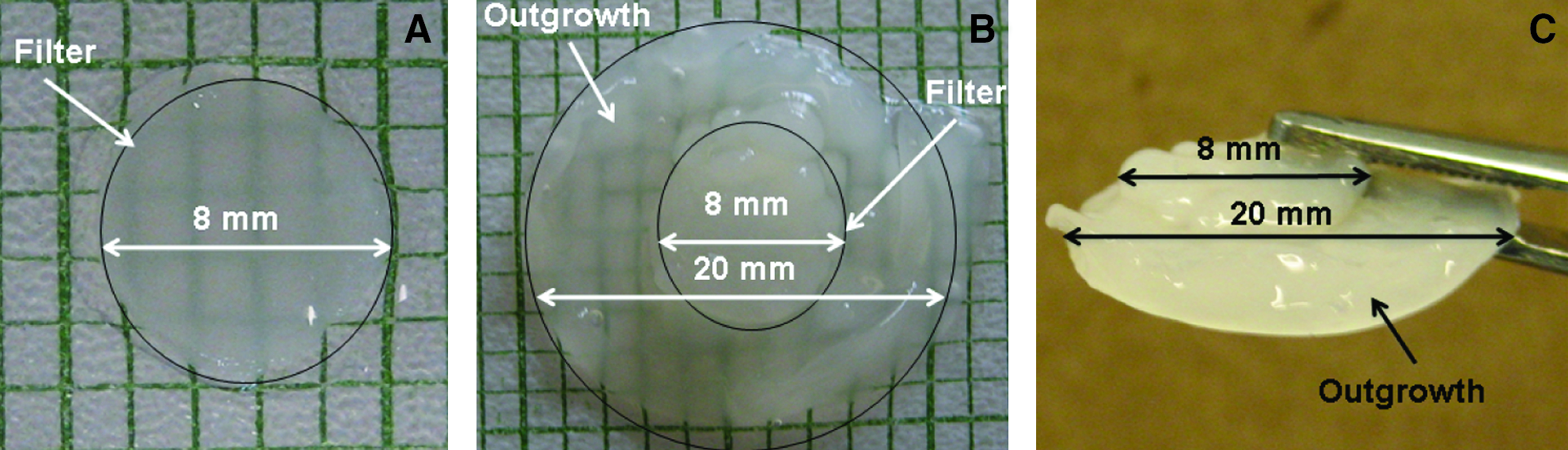
Pictorial representation of

Effect of the presence or absence of sodium bicarbonate supplementation on ECM accumulation for constructs grown in a continuous bioreactor and static conditions. Collagen and glycosaminoglycan contents of the engineered tissue constructs were biochemically determined via the dimethylmethylene blue dye binding assay and chloramine-T/Ehrlich's reagent assay, respectively. **Represents significant difference from all other groups (p<0.05). Data are presented as mean±SEM. ECM, extracellular matrix; SEM, standard error of the mean.
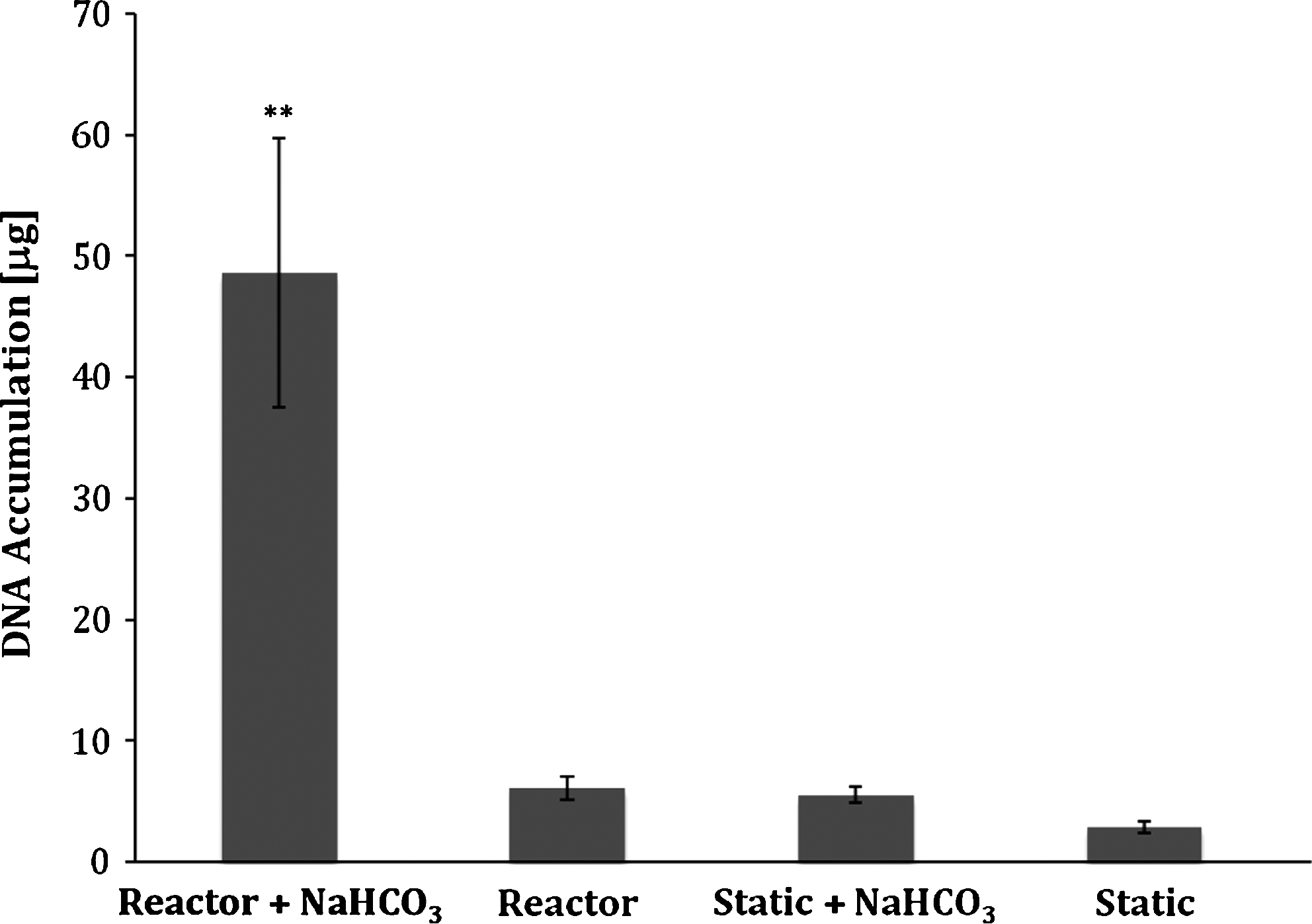
Effect of the presence or absence of sodium bicarbonate supplementation on chondrocyte proliferation. DNA contents of the engineered tissue constructs were biochemically determined via Hoechst dye 33258 assay. **Represents significant different from all other groups (p<0.05). Data are presented as mean±SEM.
n=9 per group.
Data are either expressed relative to bioreactor without NaHCO3 or static without NaHCO3.
Data are presented as mean±SEM.
Represents statistical difference with respect to bioreactor without NaHCO3 and static without NaHCO3 (p<0.05).
Represents statistical difference with respect to static without NaHCO3 (p<0.05).
Represents statistical difference with respect to static without NaHCO3 and static with NaHCO3 (p<0.05).
GAG, glycosaminoglycans; SEM, standard error of the mean.
Combined effect of NaHCO3 and continuous flow bioreactor on tissue thickness and tissue modulus
The thicknesses of the tissue constructs were measured via histology, as the results obtained were reliable and consistent. The thickness measurements obtained from histology demonstrated that the constructs cultured in the bioreactor with NaHCO3-supplemented media were significantly thicker (3380±124 μm) in comparison with all other culturing conditions, results from the latter being: reactor: 295±14 μm, static with NaHCO3: 87±4 μm; static control (no-flow): 115±4 μm (p<0.05; n=9). Further, the constructs cultured in the bioreactor with NaHCO3-supplemented medium were characterized by tissue outgrowth to average diameter 4 mm and average thickness 720±40 μm. Tissue outgrowth was negligible in all other culturing conditions. The moduli of the tissue constructs cultured in the bioreactor with NaHCO3-supplemented medium were significantly higher (p<0.05) than the bioreactor constructs without NaHCO3-supplemented medium (Table 1).
Influence of extracellular osmolarity on cell growth and matrix accumulation
To determine whether osmolarity was the underlying cause of the increased cell proliferation and ECM accumulation, the osmolarity of the medium without bicarbonate supplementation was adjusted to be equal to the osmolarity of the medium supplemented with bicarbonate. The osmolarity was adjusted by adding mannitol. Results showed that there was considerably more cell proliferation and matrix accumulation in the constructs cultured with bicarbonate than in those cultured without bicarbonate under isoosmotic conditions (Table 2). Cell proliferation appeared to be the factor that was most stimulated by bicarbonate addition, while GAG and collagen accumulation is mainly a result of an increase in cellular proliferation (Table 2). From the results obtained at week 3, it is evident, that osmotic adjustment alone could not explain increased cell proliferation and ECM accumulation, in the bicarbonate constructs (Table 2).
All under 5% CO2.
n=9 per group.
Data are presented as mean±SEM.
Represents statistical differene with respect to without NaHCO3 condition (p<0.05).
NaHCO3, sodium bicarbonate.
Combined effect of NaHCO3 and continuous flow bioreactor on extracellular pH
The pH of the medium from inside the wells was measured at the end of a 48 h media replacement cycle just before harvesting the tissue (Tables 2 and 3). A significant difference was observed between the pH of NaHCO3-supplemented medium from static (no-flow; pH: 6.91) and from static control (no-flow) without NaHCO3 supplementation (pH: 6.72). At week 5, there was no significant difference (p<0.05) observed between the bioreactor with NaHCO3-supplemented medium (pH: 6.89) and the reactor with unsupplemented medium (pH: 6.81; Table 2). During the 5 week culture period the pH of the NaHCO3-supplemented medium in the bioreactor was for the most part 6.9 or higher, while the media without NaHCO3 was mostly lower than 6.9 (Table 3). The pH of the medium-only condition with no cells or tissue, 48 h post the medium replacement cycle, was as follows: (i) medium with NaHCO3 supplementation: 7.41 and (ii) medium without NaHCO3 supplementation: 6.95 (Table 2).
Media collected from each well.
n=at least 4 per group.
Data are presented as mean±SEM.
Histological and immunohistochemical assessment
Histological assessment of the developed tissues via H&E and TB staining revealed that the cartilaginous constructs cultured in the continuous bioreactor with NaHCO3-supplemented media was significantly thicker (p<0.05) in comparison with all the other culturing conditions (Figs. 5 and 6). Tissue sections from all of the culturing conditions stained positive for sulfated proteoglycans (Figs. 5 and 6). All of the culturing conditions, except the static control (no-flow), displayed depth-dependent staining with increasingly more intense staining at the bottom of the tissue section and less staining at the tissue surface (Figs. 5 and 6). Tissues were also immunostained to detect the presence of collagen types II (cartilage-specific) and I (de-differentiation marker). All conditions stained intensely throughout the tissue thickness for collagen type II and less intensely for collagen type I (Fig. 7).

Histological assessment of cartilaginous tissue constructs after 5 weeks of culturing under different culturing conditions (static culture, static with NaHCO3-supplemented medium, and continuous-flow culture without NaHCO3 medium supplementation). Tissue sections were stained with toluidine blue (TB; sulfated proteoglycan stain) and hematoxylin and eosin (H&E; general connective tissue stain). Scale bar 100 and 200 μm. Color images available online at

Histological assessment of cartilaginous tissue constructs after 5 weeks of culturing in a continuous flow bioreactor with NaHCO3-supplemented medium. Tissue sections were stained with TB (sulfated proteoglycan stain) and H&E (general connective tissue stain). Scale bar 100 μm. Color images available online at

Immunohistochemical assessment of cartilaginous tissue constructs after 5 weeks of culturing under different culturing conditions (static culture, static with NaHCO3-supplemented medium, continuous flow culture without NaHCO3 medium supplementation, and continuous flow culture with NaHCO3 medium supplementation). Tissue sections were stained for collagen types I and II. Scale bar 100 μm. Color images available online at
Discussion
The purpose of this study was to determine whether the addition of an additional buffer, NaHCO3, to the culture media in a continuous flow bioreactor 8 would increase ECM accumulation and cell proliferation.
We found that both continuous culture and NaHCO3 buffering stimulated matrix accumulation and cell proliferation. After 5 weeks of continuous culture, constructs cultured with NaHCO3 supplementation produced markedly more GAG and DNA, both in the bioreactor and in static cultures, than in constructs with no NaHCO3 supplementation (Figs. 3 and 4; and Table 1). Looking at results from the bioreactor cultures, even though the GAG/DNA ratio increased by 65% (Figs. 3 and 4) in the NaHCO3-supplemented system relative to the unsupplemented system, the marked increase in the amount of GAG and collagen produced (Fig. 3) appears to arise mainly from a 6-fold increase in the number of cells (Fig. 4). In addition, as previously found, 8 chondrocytes cultured under continuous flow produced more matrix than chondrocytes cultured under static conditions (Figs. 3 and 4). Remarkably, in a bioreactor with NaHCO3 supplemented media, the harvested cartilaginous tissue constructs were significantly thicker (a 28-fold increase [p<0.05]) and contained significantly more (p<0.05) ECM macromolecules (a 98-fold increase in GAGs and a 25-fold increase in collagen content) compared to the static control (no-flow) cultures (Fig. 3). Histological assessment of the developed tissues revealed that all of the tissue sections from the bioreactor stained positive for sulfated proteoglycans. Tissues were immunostained to detect the presence of collagen types II (cartilage-specific) and I (de-differentiation marker). Constructs cultivated in the continuous-flow bioreactor with and without NaHCO3 medium supplementation stained more intensely for type II collagen than for type I collagen (Fig. 7). Measurements showed that the constructs cultured in the continuous-flow bioreactor with NaHCO3-supplemented medium had a greater elastic modulus than bioreactor constructs grown without NaHCO3 supplementation; however, in no case was the elastic modulus comparable to that of native articular cartilage.
From the results presented here, it is evident that chondrocytes cultured in a continuous-flow bioreactor where the medium is buffered by both 14 mM NaHCO3 and 20 mM HEPES proliferate more extensively and produce more cartilage matrix than chondrocytes cultured in a similar continuous flow bioreactor with medium buffered only by 20 mM HEPES (Figs. 3–5). The question then arises, are the effects mediated by improved buffering, or by the presence of NaHCO3 itself? Table 2 shows that there was a significant difference (p<0.05) between the pH levels of the media with and without NaHCO3 supplementation, with no exposure to cells or tissue; when allowed to equilibrate with 5% CO2 at 37°C. The medium containing NaHCO3 was at pH 7.4 whereas without NaHCO3 it was at pH6.9. However, there was little difference between the two media after exposure to cells. After 5 weeks of culture in the bioreactor, the pH of the effluent medium with NaHCO3 present was pH 6.89, whereas in its absence, the effluent pH was 6.81 (Tables 2 and 3). Thus, in the bioreactor with bicarbonate present, because of increased cell number and activity, the pH fell 0.54 pH units more during the 7 h residence time than it fell in the bioreactor with no bicarbonate supplementation. Could the differences in inlet (fresh medium) pH (Table 3) alone be responsible for the dramatic increase in cell number and matrix accumulation (Figs. 3–5)? Acidification of the extracellular medium from pH 7.4 to 6.8 is reported to reduce the rate of cell proliferation of other cell types47,48 and the rate of matrix synthesis by chondrocytes13,14,16,19,20 markedly. With no NaHCO3 supplementation, the extracellular pH of the medium fed to the cells was never above pH 7.0 (Tables 2 and 3); the acid pH of the medium in contact with the cells over the entire culture period could account, in part at least, for the lower cell numbers and amount of matrix produced in cultures which are not buffered by NaHCO3.
Does NaHCO3 play a role apart from buffering medium pH? The effect of extracellular pH on cell function is in general mediated through its influence on pHi both in regard to cell proliferation49,50 and matrix synthesis. 51 Cells thus have a complex system of membrane transporters, channels, and pumps to maintain intracellular ionic homeostasis in the face of extracellular insults. 52 Freshly isolated chondrocytes appear to regulate their pHi mainly through the sodium hydrogen exchanger,52–54 and unlike other cell types55,56 do not appear to utilize Na+/HCO3− dependent membrane transport pathways. 51 However, there are some indications, that HCO3− transporters may be involved in pH regulation in chondrocytes under some circumstances; bicarbonate transporters have been reported in a chondrocyte cell line, 57 in avian chondrocytes 53 and in surface zone articular chondrocytes. 22 The osmolarity of the medium buffered by both NaHCO3 and HEPES (325 mOsm) was ∼25 mOsm higher than the medium buffered by only HEPES (300 mOsm). Previous studies have shown a 25 mOsm difference in osmolarity to have an influence on cell proliferation and matrix synthesis.50,58,59 From the results obtained from the osmolarity experiment it is evident that the dramatic cell proliferation and tissue accumulation in the NaHCO3-supplemented media, is not solely due to increased osmolarity arising from NaHCO3 addition (Table 4). It is now important to investigate whether the mechanism of increased tissue growth arises from operating the bioreactor with optimum media pH, or if the presence of bicarbonate ions per se is required.
n=4 per group.
Data are expressed relative to bioreactor without NaHCO3 at 3 weeks.
Data are presented as mean±SEM.
The results here demonstrate the importance of maintaining pH and providing adequate buffering against lactic acid induced acidification in cartilage tissue engineering. Briefly, we compared cartilage constructs engineered in our system supplemented with NaHCO3 to other dynamic bioreactor systems, namely: (i) mixed flask bioreactor,60,61 (ii) rotating vessel bioreactor, 62 and (iii) perfusion bioreactor. 26 From the data reported it is evident that our constructs accumulated significant amounts of ECM. We postulate this could be due to the continuous supply of medium (15 mL/day per construct) that might have resulted in a near infinite supply of nutrients. Additionally, NaHCO3 supplementation provided a slightly basic environment (Tables 2 and 3), thus, maintaining the pH of the medium close to pH 7 for the most part57,63 which in turn might have helped regulate the pHi of the chondrocytes to a value close to pH 7.1,18,52 thus accounting for high GAG and collagen accumulation.
In conclusion, this study highlights the advantage of employing a continuous flow bioreactor in conjunction with appropriate buffers in adequate quantities for articular cartilage tissue engineering, thus; promoting cell proliferation and increasing matrix accumulation.
Footnotes
Acknowledgments
Funding for this research was provided, in part, by the Natural Sciences and Engineering Research Council of Canada (NSERC) and Canadian Institutes of Health Research (CIHR). The authors would also like to thank Dr. Jill Urban, Dr. Robert Wilkins and Dr. Robert Lee (University of Oxford) for their expert advice on pH, osmolarity, data interpretation/presentation, and scientific critique, Dr. Richard Boyd (University of Oxford) for allowing access to his laboratory and cell culture facility, Mr. Jackie Fan (Queen's University) for his technical assistance with mechanical analyses, Mr. John DeCosta and Mrs. Judy Pang (Queen's University) for preparing the histological and immunohistochemical sections.
Disclosure Statement
No competing financial interests exist.