
Abstract
Human skin equivalents (HSEs) are three-dimensional culture models that are used as a model for native human skin. In this study the barrier properties of two novel HSEs, the fibroblast-derived matrix model (FDM) and the Leiden epidermal model (LEM), were compared with the full-thickness collagen model (FTM) and human skin. Since the main skin barrier is located in the lipid regions of the upper layer of the skin, the stratum corneum (SC), we investigated the epidermal morphology, expression of differentiation markers, SC permeability, lipid composition, and lipid organization of all HSEs and native human skin. Our results demonstrate that the barrier function of the FDM and LEM improved compared with that of the FTM, but all HSEs are more permeable than human skin. Further, the FDM and LEM have a relatively lower free fatty acid content than the FTM and human skin. Several similarities between the FDM, LEM and FTM were observed: (1) the morphology and the expression of the investigated differentiation markers were similar to those observed in native human skin, except for the observed expression of keratin 16 and premature expression of involucrin that were detected in all HSEs, (2) the lipids in the SC of all HSEs were arranged in lipid lamellae, similar to human skin, but show an increase in the number of lipid lamellae in the intercellular regions and (3) the SC lipids of all HSEs show a less densely packed lateral lipid organization compared with human SC. These findings indicate that the HSEs mimic many aspects of native human skin, but differ in their barrier properties.
Introduction
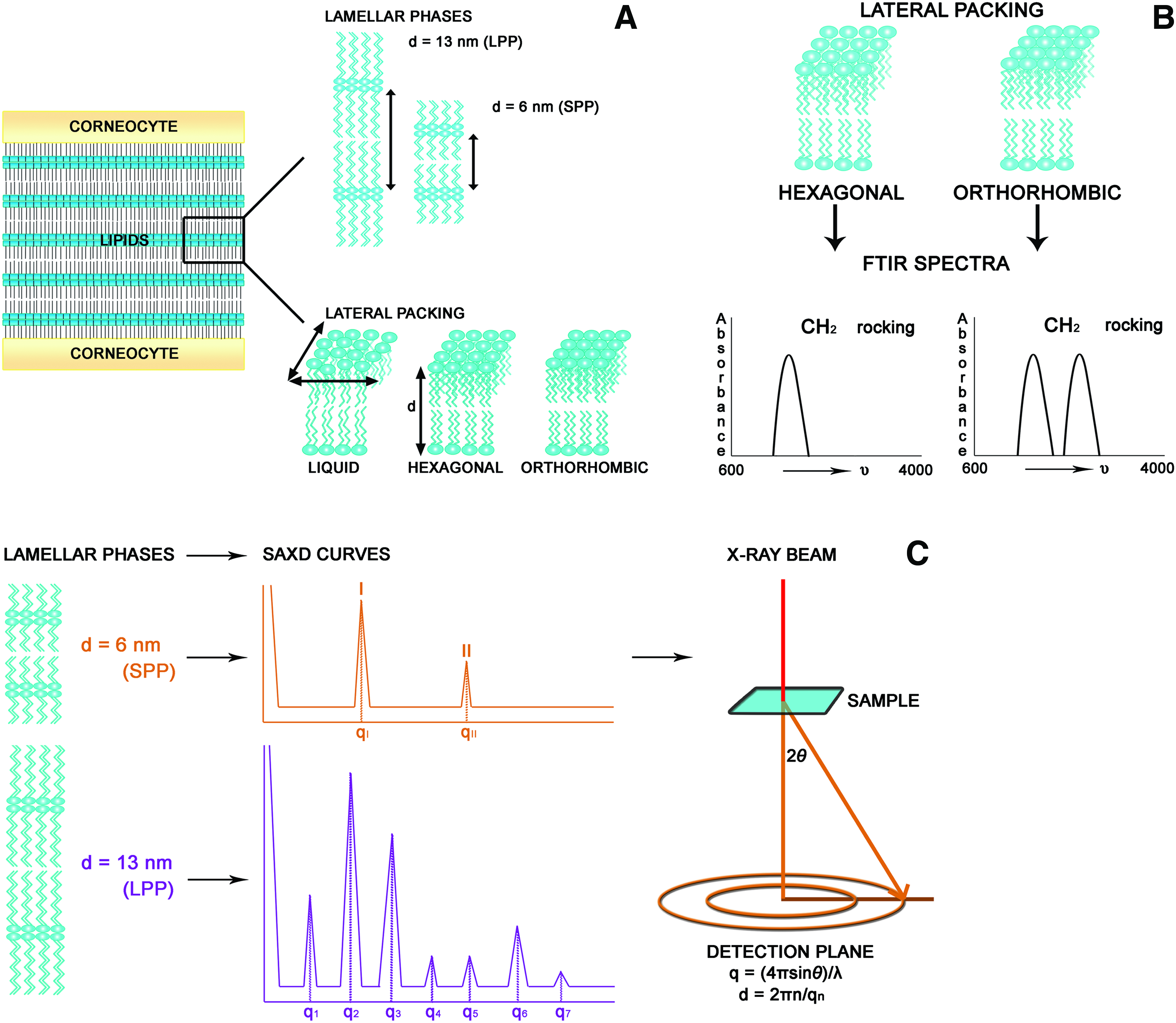
Human SC Lipid Organization.
When HSEs are used for in vitro permeation studies, they generally show a higher permeability compared with human skin indicative of an impaired barrier function.25–32 Several studies have been performed in which the barrier properties of various HSEs were examined. However, these studies mainly focused on the SC lipid composition. They showed that HSEs are prone to have a reduced free fatty acid content.14,25,33–36 The few studies in which the SC lateral and lamellar organization of some HSEs have been examined were performed more than a decade ago. These studies mostly showed a predominantly hexagonal packing34,37 in HSEs and the presence of the LPP.33,35,37 However, not all models showed a continuous presence of lipid lamellae in the SC or the presence of the LPP. 14
In this study we evaluated the barrier properties of two recently developed HSEs, the fibroblasts-derived matrix model (FDM) and the Leiden epidermal model (LEM), and compared their barrier properties, which have not been investigated before, with the full-thickness collagen model (FTM). The FDM model consists of a dermal compartment based on a human fibroblast-derived matrix, 38 whereas the dermal equivalent of FTM consists of rat-tail collagen populated with fibroblasts. 39 The LEM is an epidermal equivalent generated on an inert filter. To evaluate to which extent these three HSEs mimic the barrier properties of native human skin and to assess their suitability, for example, permeability testing, we have examined their SC lipid composition, lipid organization and permeability and compared these with native human skin.
Materials and Methods
Cell culture
Normal human keratinocytes (NHKs) and human dermal fibroblasts were obtained from adult donors undergoing mammary or abdomen surgery and were established as previously described. 39 NHKs used to create HSEs with only an epidermal compartment were generated with the Dermalife K medium complete kit (Lifeline Cell Technology). The NHKs were grown to a maximum confluency of 80% before trypsin digestion. First and second passage NHKs were used to generate HSEs.
Dermal equivalents
Collagen-type I containing dermal equivalents
Collagen-type I containing dermal equivalents were generated as described earlier. 39
Fully human dermal equivalents
The dermal compartment of the FDMs were generated as described earlier.
38
Briefly, 0.4×106 fibroblasts were seeded onto filter inserts (Corning Transwell culture inserts, membrane diameter 24 mm, pore size 0.4 μm; Corning Life Sciences) and were allowed to develop a dermal matrix for 3 weeks. A more detailed description of the generation of the dermal equivalents is provided as Supplementary Data; Supplementary Data are available online at
Generation of HSEs
HSEs generated on collagen dermal equivalents and fully human dermal equivalents, referred to as FTM and FDM respectively
FTM and FDM were generated as described by Bouwstra et al. and El Ghalbzouri et al.,38,39 respectively, with slight modifications: the HSEs were kept submerged for 2 days with the medium containing 5% FBS and for an additional 2 days with the medium containing 1% FBS. Hereafter, the HSEs were lifted to the air-liquid interface and were nourished with serum-free medium. The HSEs were grown for 16 days after seeding of the NHKs. A more detailed description of the generation of FTMs and FDMs is provided as Supplementary Data.
HSEs generated on inert filter, referred to as LEM
LEM was generated with minor modifications of the culture conditions described by El Ghalbzouri et al. 7 NHKs (0.2×106 cells/filter) were seeded directly onto cell culture inserts (Corning Transwell cell culture inserts, membrane diameter 12 mm, pore size 0.4 μm; Corning Life Sciences) and were kept submerged in Dermalife medium until confluency. Then, the HSEs were kept submerged in CnT medium (CellnTec), which was supplemented according to the manufacturer's protocol and 1% penicillin/streptomycin solution (Sigma), 1 μM α-tocopherol acetate (Sigma), 25 mM vitamin C (Sigma) and a lipid mixture of 7 μM arachidonic acid (Sigma), 30 μM linoleic acid (Sigma) and 25 μM palmitic acid (Sigma). After 2 days the HSEs were lifted to the air-liquid interface. The LEMs were cultured at the air–liquid interface for 12 days before harvesting.
Morphology and immunohistochemistry
Harvested HSEs were fixed in 4% (w/v) paraformaldehyde (Lommerse Pharma), dehydrated and subsequently embedded in paraffin. 5 μm sections were cut, deparaffinized, rehydrated, and stained with hematoxylin and eosin for analysis by light microscopy. Immunohistochemical staining of keratin 10, keratin 16, filaggrin, loricrin and involucrin was performed on paraffin sections as described earlier.39,40 Immunofluorescent staining of aquaporin 3 (Santa Cruz Biotechnology) was performed on frozen sections that were fixed in acetone. The aquaporin 3 antibody (1:50 dilution) was applied for 60 min followed by incubation with donkey anti-goat FITC-labeled secondary antibody (1:100 dilution) (Jackson ImmunoResearch Europe) for 60 min. Sections were enclosed with Vectashield mounting medium with DAPI (Vector Laboratories).
SC isolation and diffusion study
The SC of HSEs and human skin were isolated as described by De Jager et al. 41 The diffusion study with benzocaine and further analysis with HPLC to determine the flux of benzocaine through each SC sample was performed as described earlier 41 with modification of the mobile phase to methanol: water (50:50 v/v). The diffusion study was performed with at least 4 SC sheets from each HSE type or human skin.
Lipid extraction and analysis
SC samples were consecutively extracted according to a modified Bligh and Dyer procedure with the addition of 0.25M KCl to extract polar lipids as described earlier.40,42 The extracted lipids were analyzed by means of one-dimensional high performance thin layer chromatography as described earlier. 40 The ceramides are named according to the nomenclature defined by Motta et al. 43 and Masukawa et al. 44 Briefly, ceramides with a sphingosine (S), phytosphingosine (P) or 6-hydroxysphingosine (H) are linked via an amide to a fatty acid chain, which can be either an estrified ω-hydroxy (EO), α-hydroxy (A) or nonhydroxy (N) fatty acid.
Fourier transform infrared spectroscopy and small angle X-ray diffraction
The SC samples for Fourier transform infrared spectroscopy (FTIR) and small angle X-ray diffraction (SAXD) measurements were hydrated at room temperature for 24 h over a 27% (w/v) NaBr solution. The hydrated SC samples for FTIR were measured as described earlier. 45 The phase transitions of the SC lateral lipid organization were examined as a function of temperature from 0°C to 90°C with a heating rate of 1°C/4 min. At least three SC sheets of each HSE type or human SC were measured.
SAXD measurements were performed at the European Synchrotron Radiation Facility in Grenoble as described earlier. 45 SC samples were clamped in specially designed holders and measured at room temperature for 10 min. The scattered intensity was measured as a function of q. q is defined as q=4πsinθ/λ, in which λ is the wavelength of the X-rays and θ is the scattering angle. Repeat distances of lamellar phases are calculated from the peak positions; d=2πn/qn, in which n is the order of the peak and qn is its position. At least two samples of each HSE type or human SC were measured. A schematic overview of the FTIR and SAXD measurements is depicted in Figure 1B and C, respectively.
Transmission electron microscopy
HSEs were fixed in 2% paraformaldehyde −2.5% glutaraldehyde in 0.1M sodium cacodylate buffer (pH 7.4) followed by a first postfixation in 1% osmium tetroxide in cacodylate buffer and a second postfixation in 0.5% ruthenium tetroxide. Then, the samples were dehydrated in 70% ethanol and subsequently processed in a series of 70% ethanol/epoxy resin LX112 (LADD Research Industries) dilutions and finally in 100% epoxy resin. Ultrathin sections were stained with uranyl-acetate and lead hydroxide and visualized with a Fei Tecnai 12 Twin (Spirit) (Fei Europa) electron microscope. At least 2 cultures per HSE type were examined and at least 20 images per culture were made.
Results
The morphology and differentiation process show similarities between HSEs and native human skin
The FDM, LEM and FTM show a fairly similar morphology and expression of the investigated differentiation markers, except for two markers that are indicative of an activated epithelium, when compared with human skin (Fig. 2). All HSEs show the presence of all epidermal strata, including an SC. The keratinocytes become flat and elongated as they migrate and differentiate from the basal layer toward the upper spinous and granular layer. In the latter layer keratohyalin granules can be detected. Occasionally the stratum granulosum in the LEM consisted of more cell layers than observed for the FDM, FTM and human skin. Further, all HSEs show a similar expression of the early differentiation marker keratin 10, the water/glycerol transporting channel aquaporin 3 and the two terminal differentiation markers filaggrin and loricrin as human skin (Fig. 2). Filaggrin and loricrin are only detected in the stratum granulosum, whereas keratin 10 is detected in the suprabasal layers and aquaporin 3 in the cell membranes of all cells in the viable epidermis. In native human skin involucrin expression is confined to the granular layer and keratin 16 expression is absent. In all HSEs however, involucrin and keratin 16 are expressed throughout the suprabasal layers of the viable epidermis, which are signs of an activated epithelium.
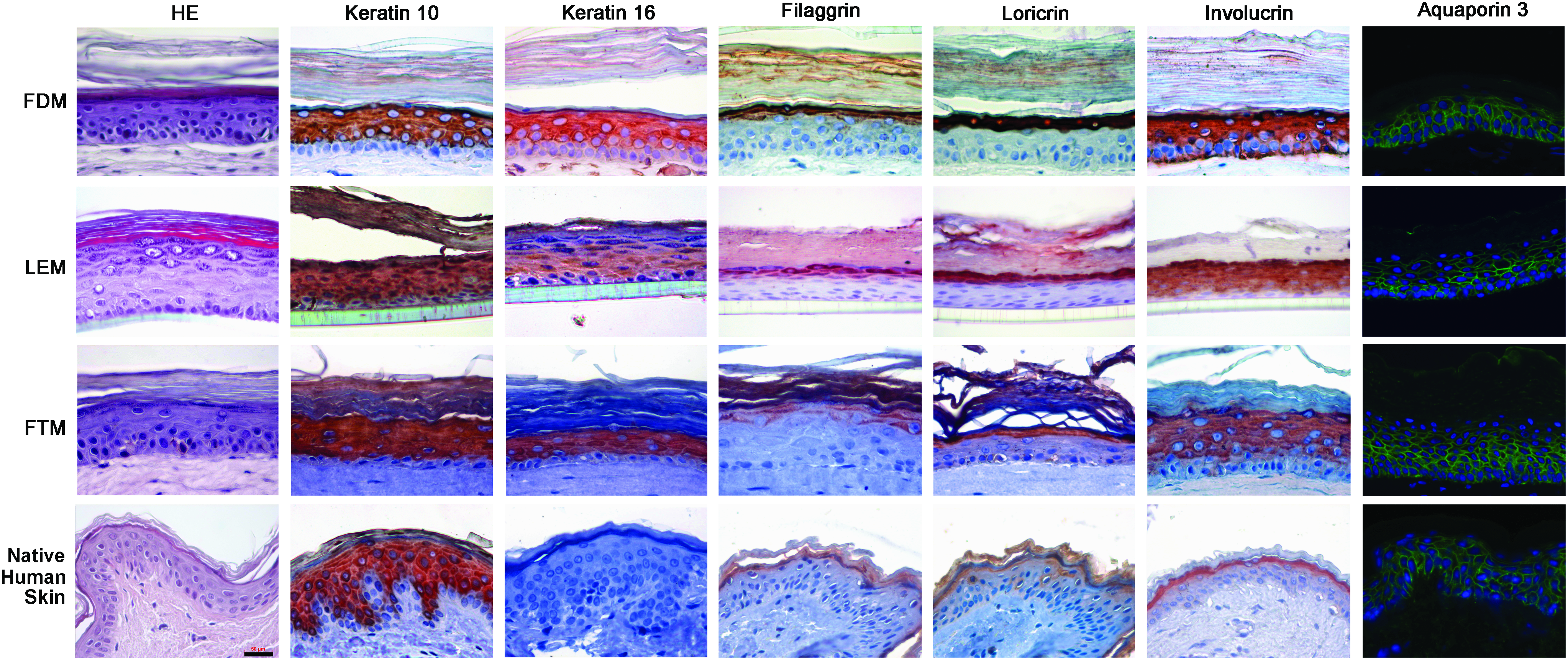
Morphology and expression of differentiation markers. Hematoxylin and eosin (HE) staining and immunohistochemical staining of the expression pattern of keratin 10, keratin 16, filaggrin, loricrin, involucrin, and aquaporin 3 (green pattern in the image) in FDM, FTM, LEM, and native human skin are shown. Scale bar represents 50 μm. FDM, fibroblast-derived matrix model; FTM, full thickness collagen model; LEM, Leiden epidermal model. Color images available online at
The SC of the HSEs contains all barrier lipid classes that are present in human skin
The three lipid classes present in human SC, namely cholesterol, free fatty acids and ceramides, are also present in the SC of the HSEs (Fig. 3). In addition, the HSEs show the presence of all ceramide subclasses that are present in native human SC. However, some differences can be noticed in the relative abundance of free fatty acids and ceramides when the different lipid profiles are compared. The FDM and LEM exhibit a lower relative level of free fatty acids compared with human SC. However, the lipid profile of the FTM indicates that in this model the free fatty acid level is higher than in the LEM and FDM and appears to be comparable to native human SC. Furthermore, all HSEs seem to have a higher relative content of ceramides EOS and EOP. The LEM and FTM have an additional ceramide subclass, with a retention factor between that of ceramide NP and AS (Fig. 3; indicated by the *), that is not present in the FDM or human SC.

Lipid analysis. The SC barrier lipid profile of each HSE and human SC are shown. The HSEs contain all lipid classes that are also present in human SC, namely cholesterol, free fatty acids and ceramides. The FTM and LEM show the presence of an unidentified lipid subclass that is not detected in human SC or the FDM. * indicates unidentified lipid. CHOL, cholesterol; FFA, free fatty acids; A, α-hydroxy fatty acids; E, ester-linked fatty acids; H, 6-hydroxy sphingosine; N, nonhydroxy fatty acids; O, ω- hydroxy fatty acids; P, phytosphingosine; S, sphingosine; HSE, human skin equivalent.
Permeability assessment of HSEs and native human skin
To investigate the barrier function of the HSEs, a diffusion study was performed. Figure 4 shows the diffusion profile of benzocaine through the SC of the three HSEs and native human skin. These results show that the FDM and LEM have an improved barrier function compared with the FTM. The calculated steady-state flux (Fig. 4) indicates that the SC of the FDM and LEM is approximately three times more permeable than native human SC, whereas the SC of the FTM is about five times more permeable. The lag time of the benzocaine flux through the SC of the FDM and FTM is slightly lower compared with native human SC.
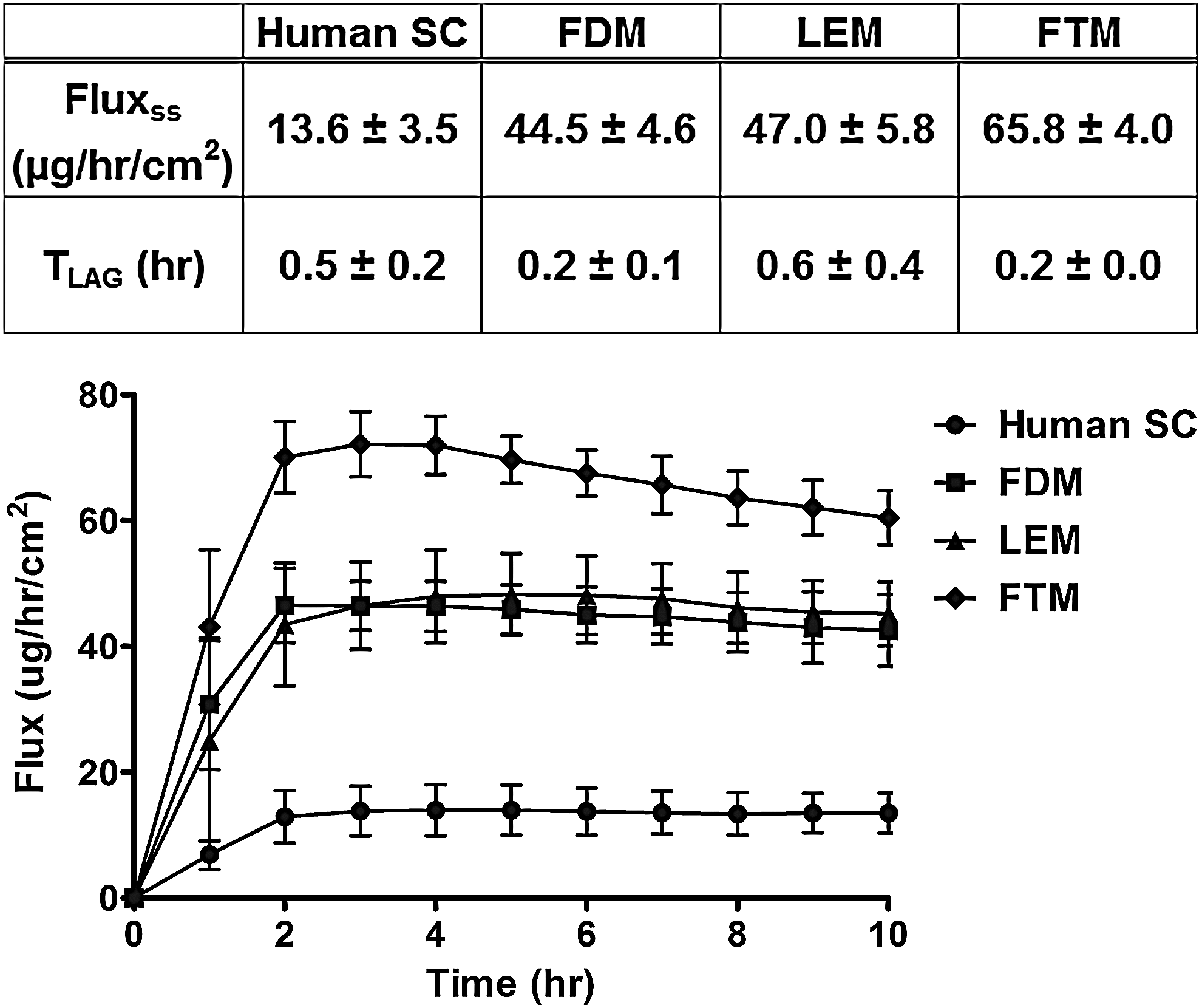
Benzocaine diffusion profile. Benzocaine was used as a model drug to investigate the permeability of the three HSEs and human SC. The data represent the mean and standard deviation of at least four measurements. The estimated steady-state flux (Fluxss) and lag time (Tlag) of benzocaine through the SC of human skin and the three HSEs are also shown.
The lateral packing in the SC of the three HSEs differs from human SC
The lateral packing in the SC of the three HSEs and human skin was examined by using FTIR. The lateral packing can be determined by monitoring the rocking bands in an FTIR spectrum. When lipids are in a crystalline orthorhombic packing the CH2 rocking band consists of two vibrations at 719 and 730 cm−1, whereas a crystalline hexagonal lateral packing results in a single vibration at 719 cm−1. An example of the thermotropic response of the CH2 rocking band as a function of temperature of the FDM, LEM, FTM, and native human SC are shown in Figure 5. In general, the HSEs show a strong peak at 719 cm−1 and a smaller peak at 730 cm−1 at lower temperatures. This indicates that in that temperature region the lipids in the SC mainly form a hexagonal lateral packing and a small population of lipids forms an orthorhombic packing. In human SC the high intensity of the 730 cm−1 peak indicates an abundant formation of the orthorhombic lateral packing. The temperature at which the orthorhombic packing transforms into the hexagonal packing, indicated by the disappearance of the 730 cm−1 peak, varies between each HSE type and also between samples of the same HSE type and native human skin. In the FDM the orthorhombic packing disappears at a temperature that varies between 8°C and 24°C, in the FTM between 14°C and 26°C, in the LEM between 24°C and 30°C and in native human SC between 40°C and 50°C. At which temperature the lipids finally form a liquid phase can be determined from the CH2 symmetric stretching mode, which provides information about the conformational disorder. In a crystalline phase (hexagonal or orthorhombic packing) the conformational disorder is low, resulting in symmetric stretching frequencies below 2850 cm−1. In a liquid phase the conformational disordering is high, resulting in symmetric stretching vibrations of around 2852–2854 cm−1. The transition from a crystalline to a conformational disordered liquid phase can, therefore, be distinguished by a steep increase in wavenumber. In human SC, the CH2 symmetric stretching is around 2848.5 cm−1 at 0°C and increases slightly when the temperature is raised. Between 30°C and 40°C there is a steeper shift indicating the orthorhombic to hexagonal phase transition. A further increase to 66°C results in a gradual shift in wavenumber. At 66°C a steep shift is noticed from 2850.9 to 2854.0 cm−1 at 86°C, which corresponds to the formation of a liquid phase (Fig. 5e). When focusing on the HSEs, between 0°C and 56°C a gradual increase in wavenumber from around 2849.5 to 2850.4 cm−1 is observed. The symmetric stretching is located at higher wavenumbers than observed for native human SC. Above 56°C a steep increase in wavenumber is observed until 82°C–84°C, from 2853.2 to 2853.6 cm−1 (Fig. 5e) demonstrating the formation of the liquid phase. The liquid phase is formed at a lower temperature in the SC of the HSEs compared with human SC.

Lateral lipid organization. Examples of the lateral lipid organization as a function of temperature are shown for the FDM, LEM, FTM, and native human SC
The SC intercellular lipid regions of HSEs are composed of more lipid lamellae than native human skin
Transmission electron microscopy images taken at the stratum garnulosum/SC interface show the lamellar body extrusion process in the HSEs, similarly as observed in native human skin (Fig. 6a, c, e, g). The extruded lipids are neatly arranged into lipid lamellae (Fig. 6b, d, f, h), which is characteristic for the SC lipid organization. When compared with human SC, it is evident that the HSEs contain a higher number of lipid lamellae between corneocytes. This finding indicates that the intercellular lipid regions in the SC of the HSEs are more pronounced than in human SC.
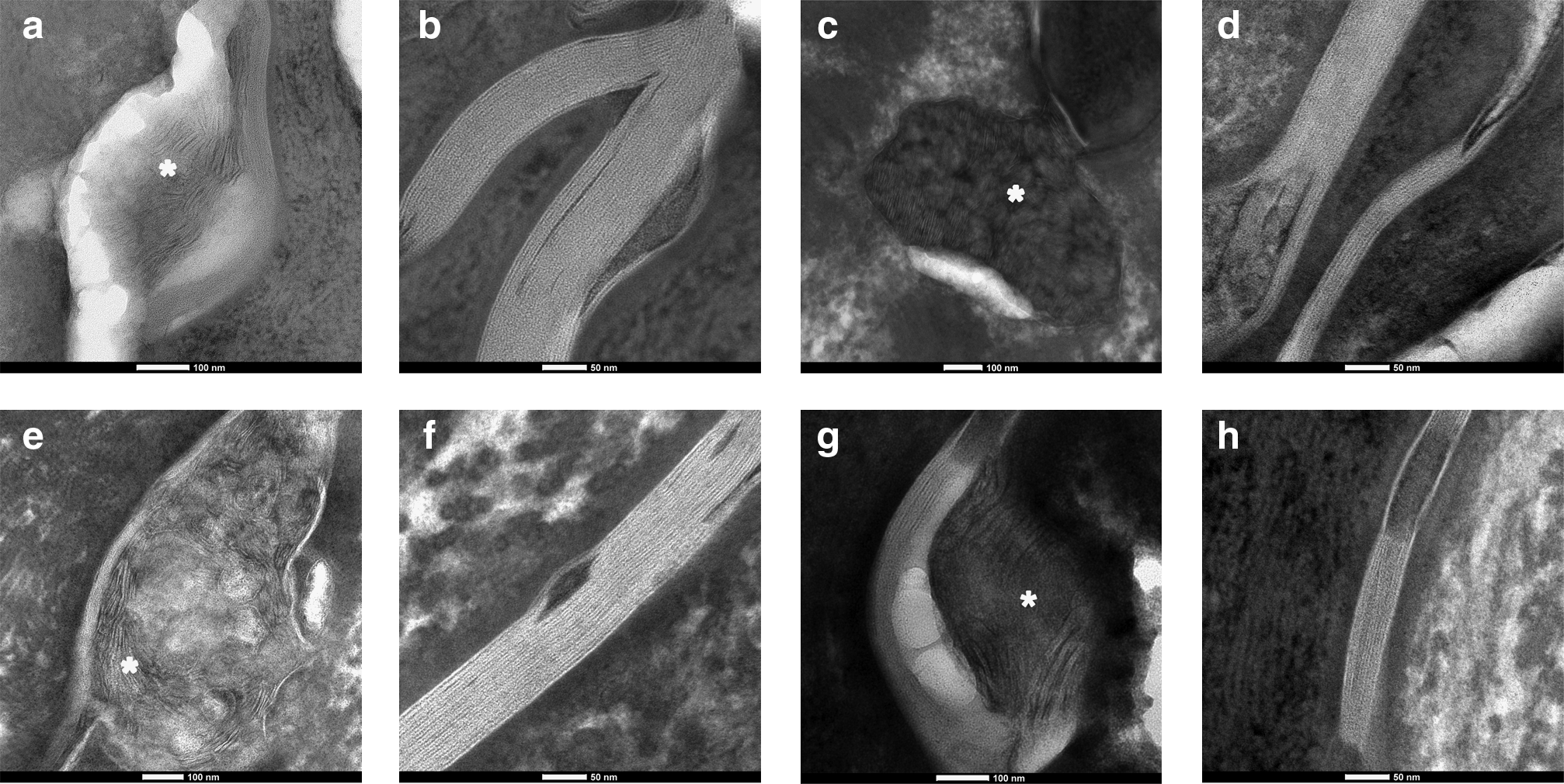
Visualization of the intercellular SC lipids. The lipid lamellae in the SC of the three HSEs and human SC were visualized with transmission electron microscopy. Representative images of each HSE and human SC are shown.
The SC lipids in HSEs form the LPP similar to native human skin
The observed lamellar organization in the SC of the three HSEs and native human SC (Fig. 6) was also investigated by SAXD. The SAXD profiles of all three HSEs show the presence of three sharp diffraction peaks (Fig. 7; referred to as 1, 2 and 3) that can be attributed to the LPP with a corresponding repeat distance varying between 11.8–12.6, 12.6–12.8 and 11.6–12.5 nm for the FDM, LEM and FTM, respectively. However, no peaks are observed that can be attributed to the SPP. The presence of crystalline cholesterol can also be detected in the diffraction profiles of all three HSEs (Fig. 7; indicated by *).

Lamellar lipid organization. A representative small angle X-ray diffraction profile of each HSE type is shown. The first, second, and third order diffraction peaks of the LPP (indicated by 1, 2 and 3, respectively) are located at q=0.50, q=1.01, and q=1.48 nm−1, respectively for the FDM, corresponding to a repeat distance of 12.6 nm. For the LEM the various orders of the LPP are located at q=0.50, q=1.00, and q=1.46 nm−1 indicating a repeat distance of 12.7 nm. For the FTM various orders of the LPP are located at q=0.52, q=1.01, and q=1.59 nm−1, which corresponds to a repeat distance of 11.7 nm. At q=1.84 nm−1, a reflection is detected in all profiles demonstrating the presence of phase-separated crystalline cholesterol (indicated by *).
Discussion
To obtain a suitable replacement of native human skin for penetration studies or for safety testing of compounds, we compared two novel in-house skin models with FTM and native human skin and evaluated their SC barrier properties. An overview of the SC barrier properties of the three HSEs and human skin is provided in Table 1. Both FDM and LEM show an improved SC barrier function compared with FTM, indicated by the higher flux of benzocaine through the SC of the FTM model. Although the SC barrier function of the novel HSEs has improved and mimics more closely the SC barrier function of native human skin, it is still reduced compared with native human skin. A reduced SC barrier function has also been reported for other HSE types, including the commercially available models Epiderm (Mattek), Episkin and SkinEthic.25–32 Since the lipid organization is a crucial determinant for the SC barrier function, we examined the lamellar phases and the lateral packing in all three HSEs and compared these with native human SC.
An overview of the SC barrier properties of native human skin (HS) and SC of the FDM, LEM and FTM are shown in the table.
RT, room temperature; CHOL, cholesterol; FFA, free fatty acid; CER, ceramides; Fluxss, steady state flux; FDM, fibroblast-derived matrix model; FTM, full thickness collagen model; LEM, Leiden epidermal model; SC, stratum corneum; LPP, long periodicity phase; SPP, short periodicity phase; arrows indicate an increase (↑) or decrease (↓).
Morphology and expression of differentiation markers
All three HSEs show a similar morphology and expression of several differentiation markers as native human skin, except for the expression of involucrin and keratin 16, which were detected throughout the suprabasal viable epidermis in all three HSEs. The observed expression of keratin 16 and the early start of the terminal differentiation program indicated by the premature expression of involucrin might be explained by an over-activated cell differentiation due to compounds or possible growth factors present in the commercial culture media or by the number of fibroblasts present in the dermal matrices.15,16 The over-activated differentiation program observed in the HSEs may have an effect on the barrier properties of the HSEs. An altered expression of differentiation markers has also been observed in the commercially available models. 12 These models showed a fully differentiated epidermis and an expression pattern for keratin 10 and loricrin that is mainly similar to native human skin. However, all commercial skin models showed a premature expression of involucrin compared with native human skin indicating that homeostasis was not yet achieved in these models. Cheng et al. 46 and El-Ghalbzouri et al. 13 showed that it is possible to generate HSEs that do not demonstrate the premature expression of involucrin and/or the presence of activation associated markers such as keratin 6, 16, and 17. However, it still remains to be investigated whether a normalized involucrin expression or the absence of activation associated markers lead to improved SC barrier properties.
Previous studies have shown that the SC of the FTMs is less hydrated than human SC. 39 The similar expression of the water/glycerol transporting channel aquaporin 3 in the HSEs and native human skin indicates that the reduced SC hydration in the FTM is not caused by the absence of aquaporin 3.
Lamellar organization
Our results demonstrate that in all three HSEs the LPP is formed, similar to native human skin, 21 whereas there is no indication of the formation of the SPP. The latter is present in native human SC. Previous studies demonstrated that the presence of acylceramides, such as acylceramides EOS and EOP, induce the formation of the LPP. 22 These ceramides are present in relatively higher amounts in the HSEs compared with native human skin and therefore may account for the abundant presence of the LPP. However, this most probably does not contribute to the reduced skin barrier function of the HSEs, as the LPP plays a more prominent role in the skin barrier function than the SPP. 41 In 2000 Ponec et al. 14 reported the lamellar organization of the EpiSkin and EpiDerm penetration models. In the EpiDerm model an LPP with a repeat distance of 12 nm was clearly detected, whereas the EpiSkin model showed signs of a poor lamellar organization. Although both the EpiDerm and Episkin models had approximately the same relative amount of ceramide EOS as native human skin, ceramides EOP, AS, AP and AH were minimally present or even absent.6,14
When we examine the electron microscopy (EM) images, the SC of the HSEs seem to contain a larger number of lipid lamellae than observed for human SC. The sharp peaks observed in the SAXD profiles clearly indicate that the mean number of lipid lamellae between the cells is increased in the SC of the HSEs compared with native human SC; the width of half maximum is proportional to 1/N, in which N is the number of lamellae in a stack. 47 Since the penetration of substances through the SC mainly proceeds via the lipid domains, we hypothesize that the increased number of lipid layers in the HSEs forms wider intercellular “channels” for compound penetration, which may attribute to the impaired skin barrier function of the HSEs. The SC intercellular lipid domains of the commercially available skin models have also been examined by EM. 14 All models showed the presence of lipid lamellae, however this was not continuously observed throughout the SC in all models. Additionally, some EpiSkin models showed an incomplete lamellar body extrusion process.
Lateral packing
When focusing on the lateral packing, HSEs have a hexagonal packing rather than an orthorhombic packing that is present in native human SC. Previous studies 18 have shown that long-chain free fatty acids (e.g., free fatty acids with a chain length of 24 or 26 carbon atoms) promote the formation of an orthorhombic packing in SC lipid mixtures, whereas a reduction in these fatty acids results in a more prominent formation of the hexagonal packing. These findings suggest that a reduced free fatty acid content in the FDM and LEM may contribute to the absence of an orthorhombic packing. The free fatty acid level in FTM mimics the free fatty acid content in human SC more closely, but nonetheless, the lipids also form a hexagonal packing. Therefore, other factors also play a role, such as a possible reduction in the chain length of the free fatty acids or the presence of unsaturated fatty acids. Based on previous studies, both these aberrations are expected to reduce the formation of the orthorhombic packing. 48 To gain more insight into this phenomenon the free fatty acid chain length distribution and degree of saturation in the HSEs will be the subject of future studies. Whether the hexagonal packing contributes to the increased permeability for substances such as benzocaine is not yet clear. Although the hexagonal packing is less dense than the orthorhombic packing, very recently no difference in the benzoic acid permeability was observed between SC lipid membranes forming either a hexagonal packing or an orthorhombic packing. 49
Formation of a liquid phase
The higher degree of conformational disorder observed for the HSEs at 32°C, which is the normal skin temperature, suggests an increased proportion of gauche conformations in the alkyl chains of the fatty acids or ceramides in the SC of HSE compared with human SC. This may signify that relatively more lipids may already be in the liquid phase at 32°C compared with the lipids in the SC of native human skin. This may render the HSEs more permeable.
When examining the transition from a hexagonal to a liquid phase, this transition occurs at a lower temperature in the investigated HSEs compared with human SC. Previous studies show that a reduced free fatty acid content cannot explain this phenomenon, as a reduction in the free fatty acid level increases the temperature at which a liquid phase is formed. 50 However, a reduced chain length of the free fatty acids or the presence of unsaturated chains may contribute to the decrease in transition temperature. 48
Optimization of the barrier properties of HSEs
The presented results show that the increased permeability of the investigated HSEs is likely caused by an increased conformational disordering of the lipids compared with the lipids in human SC. Furthermore, we hypothesize that a higher number of lipid lamellae between the cells may also reduce the SC barrier function. Whether the pronounced hexagonal packing also plays a role is not yet clear, but will be the subject of future research. The change in lipid profile compared with native human SC indicates that the lipid metabolism in these HSEs differs from native human skin. Each HSE is generated under the same incubator conditions, but still shows (slight) differences in its lipid composition. Additionally, our in-house HSEs have an improved SC lipid composition and lamellar organization compared with the commercially available HSEs studied so far. 14 Taken together, this implies that factors such as substrates used for HSE generation, culture media or other culture conditions can have an influence on the lipid metabolism in the viable epidermis resulting in changes in the SC barrier properties. Over the past years modulation of, for example, culture media or culture environment have improved epidermal development and consequently, the lipid profiles and SC barrier formation of HSEs.15,25,35,51 Ponec et al. 35 showed that addition of vitamin C to the culture medium improved the lamellar body formation, extrusion and thus, the intercellular lipid lamellae formation. The epidermal lipid metabolism was altered and resulted in an increased amount of ceramides AS and AP and detection of ceramide AH in the SC. Further optimization of culture media and culture conditions may lead to further improvements in the epidermal lipid metabolism in HSEs, such as an increase in the free fatty acid synthesis to induce the orthorhombic packing. Additionally, improvements made in the culture conditions may also enhance epidermal homeostasis, which in turn may lead to improved SC barrier properties. Further optimization of the SC barrier properties of the presented HSEs, to resemble the SC barrier properties of native human skin more closely, will be the subject of future research.
Footnotes
Acknowledgment
The authors would like to thank Dr. Maria Ponec for suggestions and discussions during their meetings and for proof reading the article.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.