
Abstract
Decellularized cardiac extracellular matrix (ECM) has been introduced as a template for cardiac tissue engineering, providing the advantages of a prevascularized scaffold that mimics native micro- and macroarchitecture to a degree difficult to achieve with synthetic materials. Nonetheless, the decellularization protocols used to create acellular myocardial scaffolds vary widely throughout the literature. In this study we performed a direct comparison of three previously described protocols while introducing and evaluating a novel, specifically developed fourth protocol, by decellularizing whole rat hearts through software-controlled automatic coronary perfusion. Although all protocols preserved the macroarchitecture of the hearts and all resulting scaffolds could successfully be reseeded with C2C12 myoblasts, assessing their biocompatibility for three-dimensional in vitro studies, we found striking differences concerning the microcomposition of the ECM scaffolds on a histological and biochemical level. While laminin could still be detected in all groups, other crucial ECM components, like elastin and collagen IV, were completely removed by at least one of the protocols. Further, only three protocols maintained a glycosaminoglycan content comparable to native tissue, whereas the remaining DNA content within the ECM varied highly throughout all four tested protocols. This study showed that the degree of acellularity and resulting ECM composition of decellularized myocardial scaffolds strongly differs depending on the decellularization protocol.
Introduction
The aim of this study was to evaluate three previously tested protocols together with a novel fourth, specifically developed protocol, which differ substantially in the nature of the used chemicals and their concentrations, and the total treatment duration, to elucidate the comparative appropriateness of the analyzed protocols with respect to specific characteristics of the resulting biomaterial.
Materials and Methods
All experiments involving laboratory animals were performed in compliance with the Guide for the Care and Use of Laboratory Animals as published by the U.S. National Institutes of Health (NIH Publication 85–23, revised 1996) and were approved by the local animal care committees (registr.nr. 35-9185.81/T47/08). The authors had full access to the data and take full responsibility for their integrity. All authors have read and agreed to the article as written.
Whole-heart explantation
Adult male LEW/Crl rats (Charles River Laboratories, Wilmington, MA) of 250–350 g were anesthetized by intraperitoneal injection of ketanest and midazolam and euthanized in a CO2 chamber. Through an immediately following median sternotomy we gained access to the retroperitoneum, where we opened the pericardium, removed the retrosternal fat body, and dissected the ascending thoracic aorta ligating its branches. After transection of all afferent and efferent vessels, we removed the heart from the chest. A prefilled 14G cannula (B. Braun, Melsungen, Germany) inserted into the thoracic aorta was used to rinse the coronary vasculature with lactated Ringer's solution (B. Braun) containing heparin 10 IU/mL (Ratiopharm, Ulm, Germany) and allowed for subsequent decellularization through coronary perfusion.
Decellularization of whole rat hearts through coronary perfusion
We developed a fully automatic software-operated control system for organ decellularization through long-term perfusion that was used for an automatic sterile decellularization of the explanted whole rat hearts through coronary perfusion allowing for a standardized constant perfusion pressure.
A closed perfusion circulation was installed, where a modified 100 mL glass flask (Schott-Duran, Mainz, Germany) served as perfusion chamber connected to a silicon tube system (Carl Roth, Karlsruhe, Germany). Coronary perfusion was enabled through connection of the thoracic aortic cannula to the tube system (perfusion inlet) with a right atrial incision allowing coronary drainage (perfusion outlet). During the decellularization process the explanted heart was kept floating freely in the perfusate. Perfusion circulation was driven by a peristaltic pump (Stöckert; Sorin Group Germany, Munich) that was automatically operated online by a specifically engineered software-based process control system (Fig. 1).
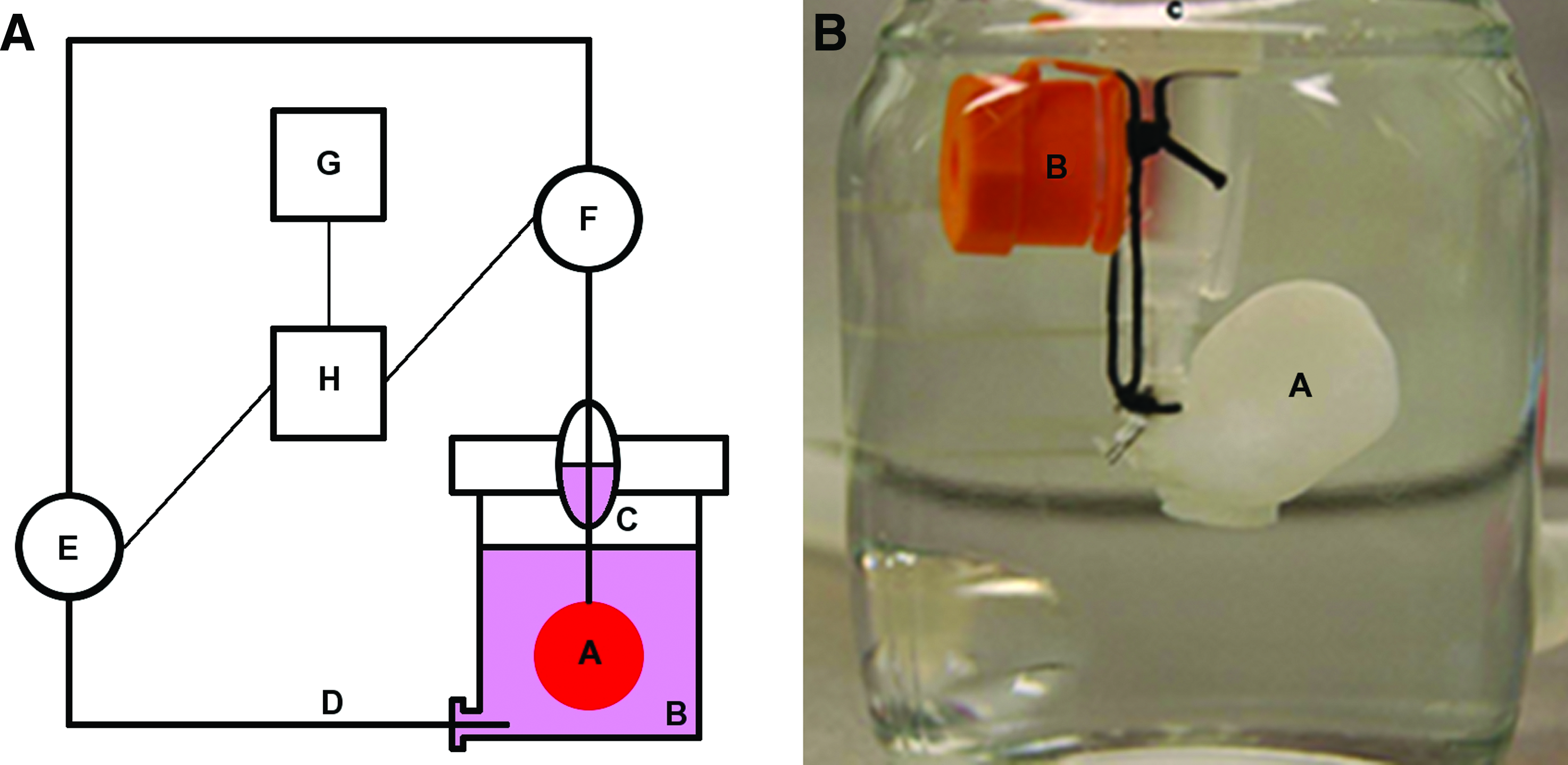
Automatic software-controlled system for whole-heart decellularization through coronary perfusion.
Therefore, a data acquisition device was connected to a pressure transducer (Maquet, Hirrlingen, Germany) connected in series to the perfusion inlet via an analog input module and to the peristaltic pump driving perfusion circulation via an analog output module. Over a netbook connected to the data acquisition device, the TissueController software from engineo GmbH (
Additionally, the control software visualized all data online and recorded them as an Excel file (Microsoft) for documentation and further analysis purposes. This allowed to exclude hearts from the analysis after decellularization with inadequate perfusion pressure, for example, because of a vascular rupture. According to the nature of the different decellularization protocols, the perfusion solutions and perfusion times varied (see the next section).
Decellularization protocols
Four different decellularization protocols with previous successful application for soft tissue decellularization were comparatively evaluated. Two of them were taken from recent literature on cardiac tissue engineering, having been used for the creation of cardiac tissue derived acellular scaffolds.8,9 The third protocol was adapted from a protocol that we and others have successfully used for the decellularization of cardiac valve-bearing tissue. 7 Based on literature review as well as own previous experience, we developed an additional novel protocol specially intended for the decellularization of myocardial tissue.
Protocol I
The concept of whole-heart decellularization combined with subsequent repopulation of the matrix with murine myoblasts (C2C12 cells) has been demonstrated to result in macroscopically contractile structures. 8 Following this concept, heparinized phosphate-buffered saline solution (PBS; Ratiopharm; Biochrom, Berlin, Germany) containing 10 mM adenosine (Sanofi-Aventis, Frankfurt, Germany) was perfused for 15 min, followed by 1% sodium dodecyl sulfate (Gerbu, Wieblingen, Germany) in deionized water for 12 h, deionized water for 15 min, and 1% Triton-X-100 (Merck, Darmstadt, Germany) in deionized water for another 30 min. Finally, the heart was perfused with PBS (Biochrom) containing 100 UI/mL penicillin-G/streptomycin and amphotericin B (PAA, Coelbe, Germany) for 124 h.
Protocol II
The second protocol was adopted from recent experiments on porcine whole-heart decellularization, reported by of one of the pioneering groups in the field of tissue decellularization. 9 As described by the group of Badylak and colleagues, hearts were initially perfused with 0.025% trypsin and 0.05% ethylenediaminetetraacetic acid (EDTA) (PAA) in PBS at 37°C for 1 h, followed by 15 min with deionized water, 1 h with 3% Triton-X-100 (Merck) in deionized water, 15 min with deionized water only, 1 h with 4% deoxycholic acid (DCA; Sigma) in deionized water, 15 min with deionized water, and 1 h with 0.1% acetic acid (Fluka, Munich, Germany) in deionized water.
Protocol III
A protocol, initially introduced by Haase et al. for the decellularization of peripheral nerves 10 was proven successful for the creation of acellular scaffolds derived from the skeletal musculature. 11 In previous experiments we have applied this protocol as a potent decellularization treatment for heart valves with remarkable degree of cell removal and good preservation of the ECM. 7 Here, we applied this method to decellularize whole-hearts. After a priming perfusion with 20% glycerol (AppliChem, Darmstadt, Germany), 0.05% NaN3 (Carl Roth), and 25 mM EDTA (AppliChem) in 0.9% NaCl for 3 days (d), perfusion was continued with 4.2% DCA (Sigma) and 0.05% NaN3 (AppliChem) in deionized water for 3 days, another solution containing 20% glycerol, 0.05% NaN3, and 25 mM EDTA in 0.9% NaCl solution for 2 days, and 1% SDS (Gerbu) and 0.05% NaN3 for 2 days. Continuing perfusion, a solution containing 3% Triton-X-100 (Merck) and 0.05% NaN3 in de-ionized water was applied for 2 days, and, finally, PBS (Biochrom) solution containing 100 IU/mL penicillin-G/streptomycin (PAA) was perfused for 12 h.
Protocol IV
We developed protocol IV by combining different, independently acting decellularization steps. 12 Saponin, a mild nonionic detergent that so far has rarely been used for the decellularization of tissues and organs, was added because of theoretic considerations on its additive advantage in the disruption of lipid–lipid and lipid–protein interactions and solubilization of cytoplasmatic and nuclear cellular membranes in form of micelles in combination with commonly used ionic detergents like SDS and DCA. 13
For this novel protocol we used heparinized PBS (Ratiopharm; Biochrom) containing 10 mM adenosine (Sanofi-Aventis) as perfusate for 15 min, after 12 h perfusions with 1% SDS (Gerbu), 1% DCA (Sigma), and 0.05% NaN3 (AppliChem) in deionized water; 20% glycerol, 0.05% NaN3, and 25 mM EDTA in 0.9% NaCl solution; 1% Saponin (Sigma) and 0.05% NaN3 (Carl Roth) in deionized water; and then again 20% glycerol, 0,05% NaN3, and 25 mM EDTA in 0.9% NaCl solution. These steps were alternated with 15 min perfusions of deionized water between them. To complete the decellularization process, we finally perfused the hearts with 200 IU/mL DNase I (Roche, Ludwigshafen, Germany) and 50 mmL/mol MgCl (Fluka) in PBS (Biochrom).
Histological and immunohistochemical examination
Decellularized hearts were fixed in 4% buffered formaldehyde solution (Roth) and cross sections of three different standardized regions (apical region, mid-ventricular region, and basis-near region) were stained with hematoxylin and eosin (H&E), picrosirius red (PSR), and fast green (FG), and Movat's pentachrom staining to validate the protocols' decellularization effectiveness and the integrity of the remaining ECM using standard protocols. Laminin, collagen IV, and sarcomeric α-actin were evaluated immunohistochemically following standard protocols for basement membrane integrity and observation of cellular remnants, respectively (for details please refer to the Supplementary Material).
Biochemical examination
For a comprehensive analysis of the decellularization success, donor cellular protein retention was further quantified by comparing the amounts of noncollagenous protein of the scaffold matrices as an estimate of the total protein content relative to the amounts found in the native myocardial tissue. Therefore, we adapted a method previously described by Joseph et al. 14 allowing the quantification of the noncollagenous protein fraction of the ECM scaffold using a PSR and FG colorimetric analysis (for details please refer to the Supplementary Material).
ECM specimens from the apex, septum, and left and right ventricle from each individual decellularized heart were assessed for residual DNA content and overall glycosaminoglycan (GAG) composition for further assessment of the decellularization success and ECM scaffold characterization, respectively. Furthermore, the specific ECM proteins elastin, laminin, and collagen type I were quantitatively analyzed for each specimen by western blot (for details please refer to the Supplementary Material).
Biocompatibility and myogenic cellular repopulation of myocardial ECM
To determine the biocompatibility of the decellularized ECM scaffolds, a direct and two indirect assays using differentiated C2C12 myoblasts were applied. For indirect biocompatibility testing, cardiac ECM scaffolds were perfused after the decellularization process with PBS and C2C12 culture media for 12 h each in the described automatic software-controlled perfusion system to yield decellularized ECM-conditioned medium. Myoblasts were then incubated with preconditioned media for 24 h and cell viability was assessed via a metabolic activity assay and a live/dead assay.
Direct biocompatibility of the decellularized scaffolds was evaluated by surface reseeding with myoblasts onto scaffold specimens from the left and right ventricles of the hearts. After 24 h, cell viability and morphology was evaluated with a standard live/dead-staining of the recellularized scaffolds and an LSM 510 confocal microscop (Carl Zeiss, Göttingen, Germany) for examination (for details please refer to the Supplementary Material).
Statistical analysis
Tests were performed as triplicate specimens and results are presented as mean±standard error. Variations among the study groups were evaluated by one-way analysis of variance with a Bonferroni post-hoc test using SPSS statistical software package. A value of p<0.05 was considered as statistically significant.
Results
Macroscopic preservation of native myocardial ECM
For each protocol we decellularized and analyzed 15 rat hearts under software-controlled coronary perfusion as previously described (Fig. 1). After decellularization whole-heart templates were examined macroscopically for structural integrity and preservation of native myocardial ECM. All produced scaffolds showed intact 3D geometry, maintaining their basic atrial, ventricular, and valvular architecture as well as a preserved vascular coronary system (Fig. 2). On macroscopic observation, the myocardial tissue changed to a complete translucent tissue in all of the cardiac ECMs generated by protocols I, III, and IV. Only in scaffolds produced by protocol II fractions of higher optical density of up to 50 mm2 were still visible (Fig. 2). These areas of increased optical density were most frequently located at the apex of the hearts, suggesting that donor cellular remnants possibly failed to be washed out in this region. On those same scaffolds generated by protocol II, an increased amount of interstitial air bubbles accumulating during the decellularization process was visible. This finding was located exclusively to the matrix vicinity (as opposed to the cavity of the ventricular chambers) in group II. A slight increase of interstitial air entrapment was observed in those individual hearts that were exposed to an increased flow rate during the automatically adjusted coronary perfusion system (constant parameter: perfusion pressure of 77.5 mm Hg). Several technical measures were taken to exclude the possibility of embolization of macrobubbles via perfusion (e.g., installation of a collecting reservoir). However, over the entire decellularization process, a gradual loss of vascular resistance, which became evident through a need for an increase of perfusion rate to maintain a constant perfusion pressure, led to the gradual accumulation and entrapment of air bubbles and could not be completely prevented.

Macroscopic preservation of native myocardial ECM after whole-heart decellularization through software-controlled automatic coronary perfusion. Representative macroscopic images of decellularized whole rat hearts of all 4 protocols show transluminent myocardial tissue with intact three-dimensional heart geometry, maintaining their basic atrial, ventricular, and valvular architecture as well as a preserved vascular coronary system. Occasionally, macroscopically identifiable cellular debris was seen as an area of higher optical density at the apical region. Ao, aorta; LA, left atrium; LV, left ventricle; RV, right ventricle; cv, coronary vessel; cd, cellular debris; ECM, extracellular matrix. Color images available online at
Histological and immunohistological examination of decellularized myocardial ECM scaffolds
We examined three whole-heart scaffolds produced by each protocol through histology and immunohistology to determine major ECM constitutes. From each heart three cross sections of different standardized regions (apical region, mid-ventricular region, and basis-near region) were examined individually, revealing striking differences in the microarchitecture and in the constitution of the ECM.
Scaffolds from protocols I and II showed areas with large amounts of diffuse basophilic structures in the H&E staining (Fig. 3), between which blurry nuclei-like shadows as well as scattered intact nuclei could be clearly identified. The impression of incomplete decellularization was hardened by the Movat and PSR/FG staining (Fig. 3) showing insular regions with increased cellular residua, which accumulated in scaffolds of protocol II preferentially in the apical region, confirming the macroscopic impression of inconsistent cell removal in this region by protocol II. Further, in the specimens of protocol II sarcomeric α-actin immunostaining (Fig. 3) identified myocardial cytoplasmatic protein remnants throughout the ECM in the apical as well as mid-ventricular and basis-near region. Movat staining showed a very heterogenous ECM of scaffolds produced by protocols I and II, with the hearts decellularized by protocol II having a fissured and slightly fractioned appearance, which was not observed to the same degree in the scaffolds produced by the other protocols. On the contrary, the scaffolds generated by protocols III and IV showed a more uniform and homogenous ECM, with no nuclei or diffuse basophilic structures showing in the H&E staining (Fig. 3). Scaffolds of protocol III showed an intact ECM, comparable to the native ECM with a strong proportion of collagen fibers and GAG in the Movat staining. No cytoplasmatic remnants could be identified in those scaffolds, neither in the PSG/FG nor in the α-actin staining. While the ECM of the hearts decellularized by protocol IV was comparable to the ones of protocol III as far as ECM microarchitecture is concerned, on a number of cross sections of ECM IV insulated small cellular remnants were identified upon PSG/FG staining and α-actin immunostaining (Fig. 3).
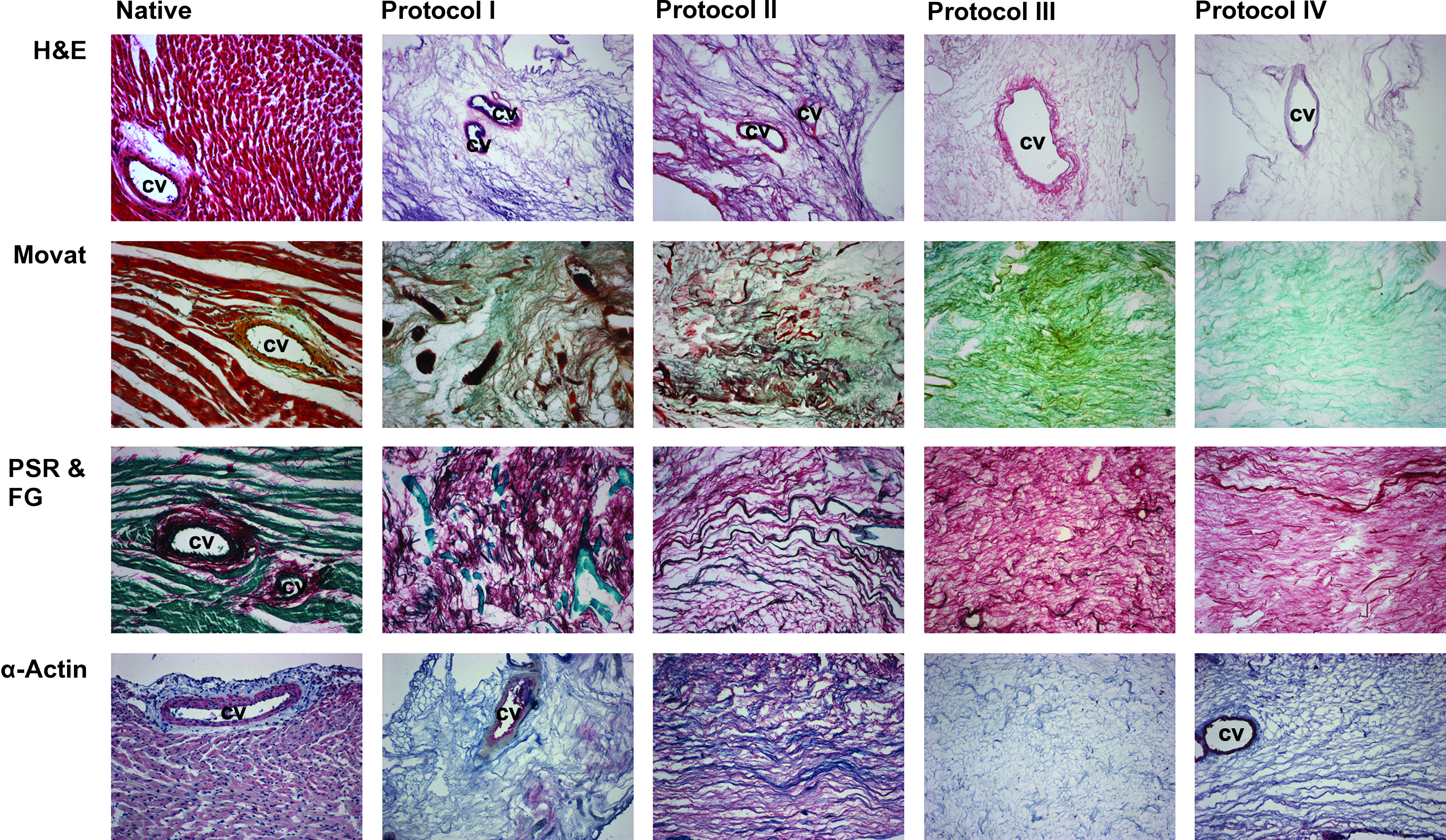
Myocardial ECM integrity and constitution after whole-heart decellularization through software-controlled automatic coronary perfusion. Representative images of native myocardium and decellularized myocardial ECM scaffolds stained with H&E, Movat, PSR/FG, and α-actin. All scaffolds show integrity and preservation of the native coronary vascular system. H&E, hematoxylin and eosin; PSR & FG, picrosirius red and fast green; cv, coronary vessel. All pictures were taken on 100× magnification.
Donor cellular protein retention of the decellularized myocardial ECM scaffolds was further quantified by comparing the amounts of noncollagenous protein of the scaffold matrixes as an estimate of the total protein content relative to the amounts found in the native myocardial tissue, adapting a method previously described by Joseph et al. 14 As already seen in the histological examination, donor cellular protein retention varied according to the applied decellularization protocol. While the relative amount of noncollagenous protein in the ECM was substantially reduced after decellularization by all four protocols, protocol I yielded scaffolds with significantly higher retention of donor cellular protein within the ECM, as compared to the other protocols (p<0.05), with still 13.90%±2.11% of the native noncollagenous protein content remaining (Fig. 4). In contrast, scaffolds generated by protocol II only retained 4.25%±2.29%, protocol III 5.67%±2.53%, and protocol IV 7.00%±1.14% of the native noncollagenous protein content, with no statistical difference between those three protocols.
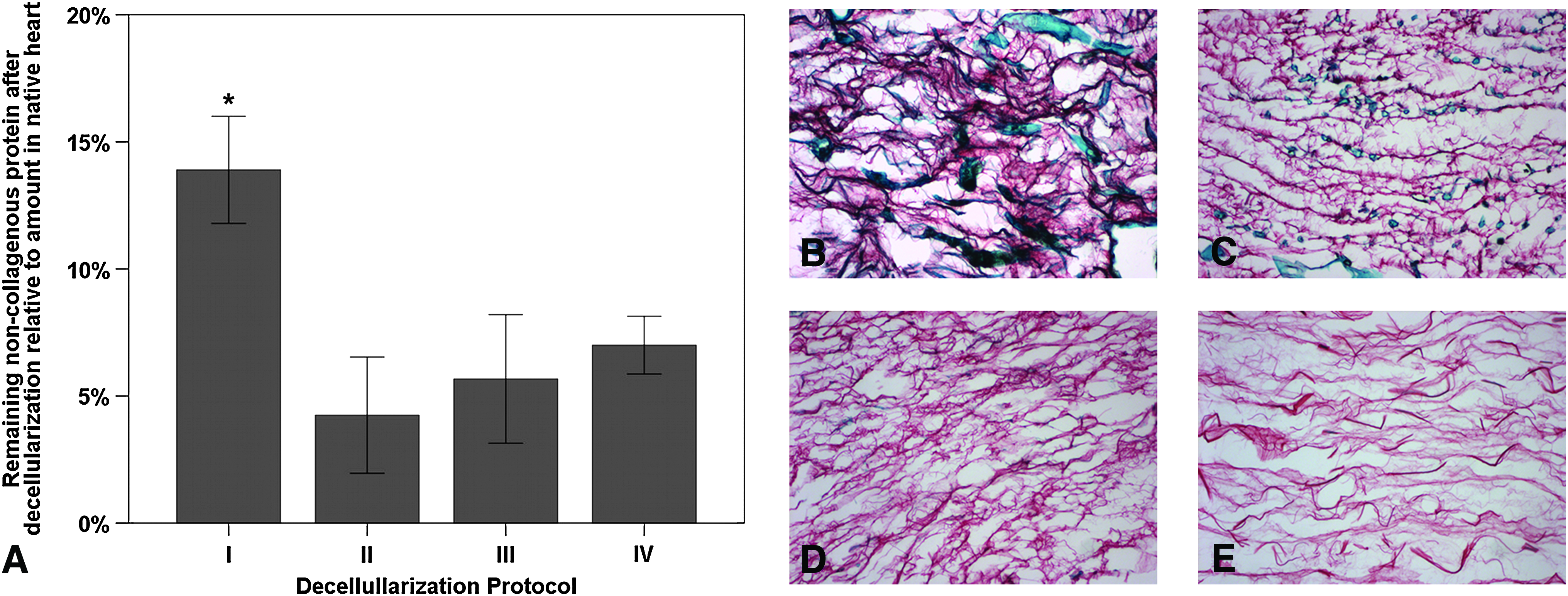
Donor cellular protein retention of the myocardial ECM after whole-heart decellularization through software-controlled automatic coronary perfusion.
Upon Immunostaining, collagen type IV could only be identified in scaffolds of protocols I, III, and IV with varying intensity throughout all of the examined cardiac regions, with larger presence evident in the apical part (Fig. 5E). In contrast, immunostaining for laminin showed heterogenous amounts of laminin in decellularized scaffolds of all protocols. However, based on the qualitative evaluation by immunohistochemistry, the most homogenous distribution of laminin was observed in scaffolds obtained through protocol IV, whereas the highest staining intensity was found in scaffolds of protocol I (Fig. 5E). Laminin could also be detected in the scaffolds of all four protocols through western blot analysis (Fig. 5A); however, a quantitative analysis revealed a substantial reduction of the laminin content of the ECM for all protocols when compared to the native heart (Fig. 5B). Scaffolds generated by protocol I retained the highest laminin amount with 33.68%±10.02% of the native amount (p<0.001), whereas other scaffolds only displayed between 2% and 5% of the native amount with no statistical difference between them (Fig. 5B). The cardiac ECM decellularized by protocol I also retained the highest amount of elastin with 7.58%±3.17% of the native amount compared to 3.41%±0.64% of the ECM from protocol II and 2.48%±1.19% of the ECM of protocol III (p<0.05) (Fig. 5C). With the antibody used in our laboratory, elastin could not be detected in scaffolds produced by protocol IV.
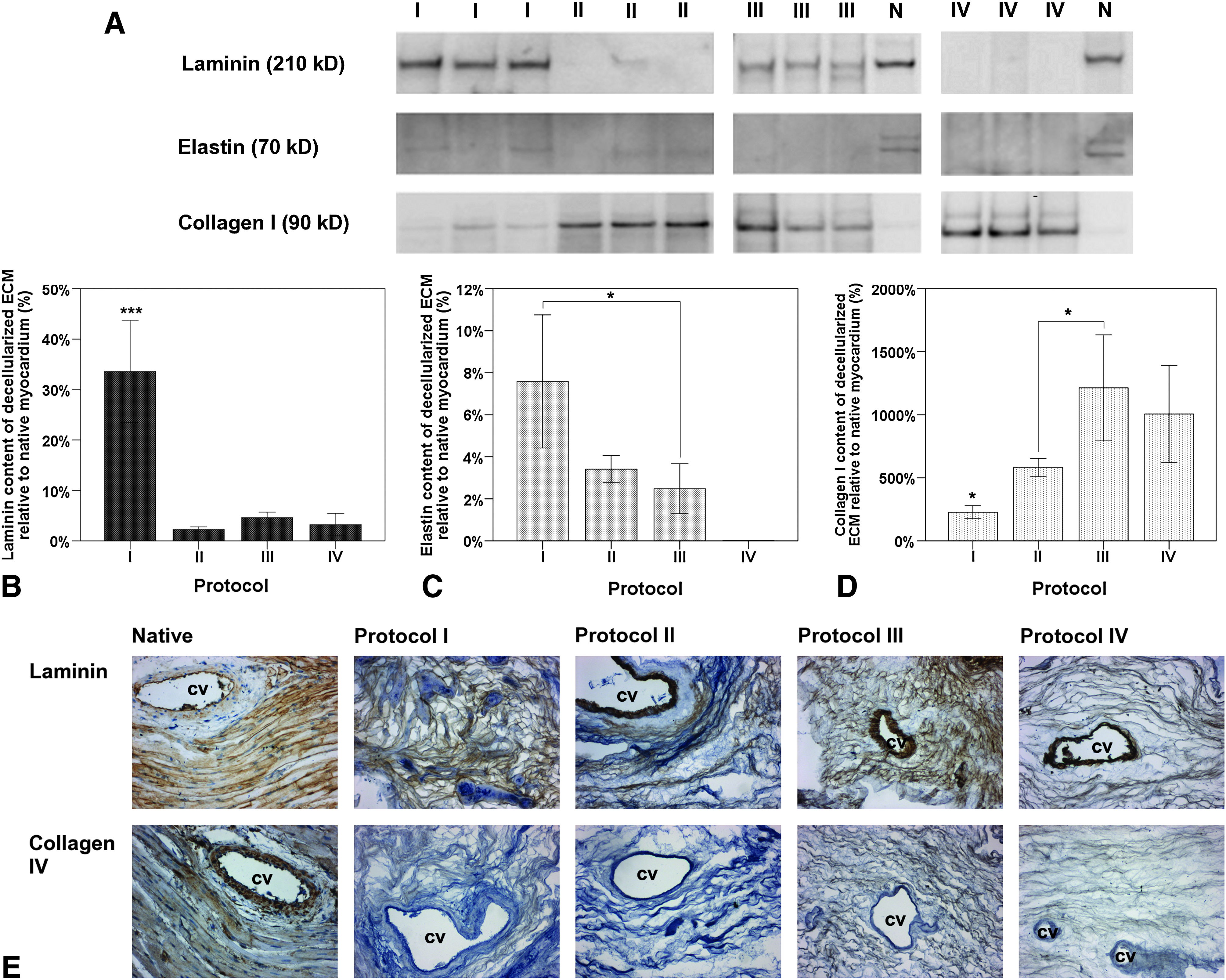
Immunohistological and molecular biological examination of major ECM constitutes of the myocardial ECM after whole-heart decellularization through software-controlled automatic coronary perfusion. Laminin, elastin, and collagen type IV in the myocardial ECM were significantly reduced after the decellularization process when compared to the amounts in the native myocardium, with scaffolds generated by protocol I retaining the highest amounts of laminin and elastin. However, relative content of collagen type I in overall ECM composition substantially increased after decellularization as compared to the native myocardium, with the ECM from scaffolds generated by protocols III and IV consisting of the highest amounts.
The amounts of collagen type I of the decellularized hearts as an estimate of the total ECM protein compared to the relative amount in the native heart could also be determined. Due to the removal of cellular protein during the decellularization process the relative amounts of collagen type I of the generated scaffolds substantially increased, however, varying widely from protocol to protocol with protocol I and II increasing the relative amount up to 2 and 5 times, respectively, and protocol III and IV up to more then 10 times when compared the to relative amount found in the native myocardium (Fig. 5D). Therefore, analysis comparing the different protocols between themselves revealed that the relative ECM composition of the scaffolds generated by protocols III and IV consisted of almost five times more collagen type I than scaffolds from protocol I and almost twice as much more collagen type I than as scaffolds from protocol II (p<0.05).
Western blot analysis of these three ECM components was also done individually for four standardized regions of the decellularized hearts (apical, septal, left ventricular, and right ventricular region), however, revealing no statistical difference in regional retention or distribution within the decellularized hearts of individual protocols.
Preservation of GAG and residual DNA content in decellularized myocardial ECM scaffolds
In a detailed analysis, we further evaluated ECM specimens decellularized by each protocol and obtained from following myocardial regions for their ECM and cellular content: apex, septal wall, and left and right ventricular free wall. Three different hearts were used for each group to determine the residual GAG and DNA content. The determined amounts of GAG in the cardiac ECM after decellularization according to protocol I (255.53±0.37 μg/g), protocol II (232.13±55.42 μg/g), and protocol III (202.90±64.65 μg/g) showed no statistical difference when compared to native myocardial tissue (219.33±31.28 μg/g) (Fig. 6F). In contrast, decellularized ECM scaffolds generated by protocol IV exhibited a statistically significant decrease of almost 70% (66.87±4.21 μg/g) of the native GAG amount (p<0.001). Upon regional analysis, residual GAG content remained homogenously distributed between the apex, septal wall, and left or right ventricular free wall, regardless of the used decellularization protocol (Fig. 6G–J).
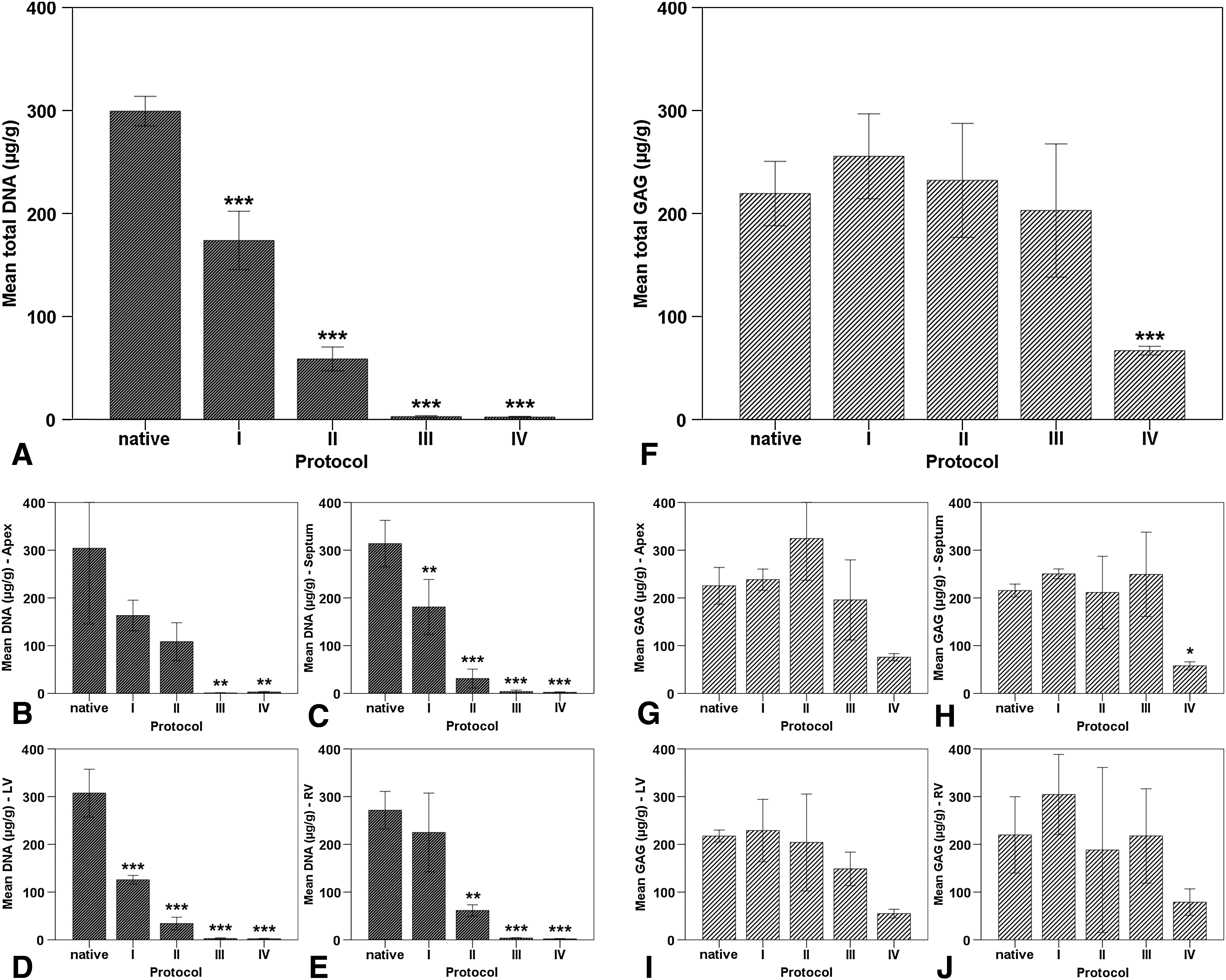
DNA and GAG content in myocardial ECM after whole-heart decellularization through software-controlled automatic coronary perfusion. All scaffolds showed a significant reduction of the DNA content after decellularization. However, the residual DNA content varied widely depending on the decellularization protocol and the myocardial region, with protocol I retaining the highest DNA amount. GAG content after decellularization was only significantly reduced in scaffolds generated by protocol IV, maintaining a homogenous regional distribution through all protocols.
Residual DNA was quantified as a further measure of decellularization quality using a standard DNA quantification kit (Qiagen, Hilden, Germany). Though scaffolds of all protocols showed a statistical reduction of the residual DNA content as compared to the native heart, differences between the individual protocols were remarkable (Fig. 6A). While protocol I was able to remove around 43% of the native DNA (173.77±0.37 μg/g) and protocol II resulted in a reduction of ca. 80% of native DNA (58.80±11.59 μg/g), only protocol III (2.80±1.02 μg/g) and protocol IV (2.40±0.69 μg/g) generated scaffolds with less than 0.5% of the native DNA content with this difference being highly significant (p<0.001). It was also noticed that the DNA amounts of the different specimens taken from hearts decellularized by protocol II fluctuated remarkably, depending on the myocardial region of origin. Interestingly, scaffolds generated by protocol I retained the highest amount of residual DNA in the right ventricular free wall and scaffolds generated by protocol II retained the highest amount of residual DNA in the apex, whereas in scaffolds from protocols III and IV, DNA removal was homogenous in all regions (Fig. 6B–E). Overall, the left ventricular free wall seemed to be the region with the most efficient DNA removal independently from the decellularization protocol.
Cytotoxicity of decellularized myocardial ECM and its suitability for a de novo cellular repopulation
In order to determine the cytotoxicity of the decellularized myocardial ECM, first an indirect biocompatibility study was conducted exposing differentiated C2C12 myoblasts to conditioned culture media after perfusion through the decellularized myocardial ECM for 12 hours. Cell viability was subsequently evaluated by a cell viability assay via live/dead staining with sybr green and propidiumiodid (Fig. 7A) and by a proliferation assay based on 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt (MTS) assessing the metabolic activity (Fig. 7B) to further validate the viability results. Based on these assays, no evidence for an in vitro cytotoxicity of any of the produced scaffolds could be detected. After 24 h of culture in conditioned media, a high cell viability of ca. 95% was observed independently from the respective decellularization protocol showing no significant differences to cells incubated in control standard culture media. Metabolic activity levels that were comparable for cells incubated either in conditioned media or in control standard media confirmed these results.
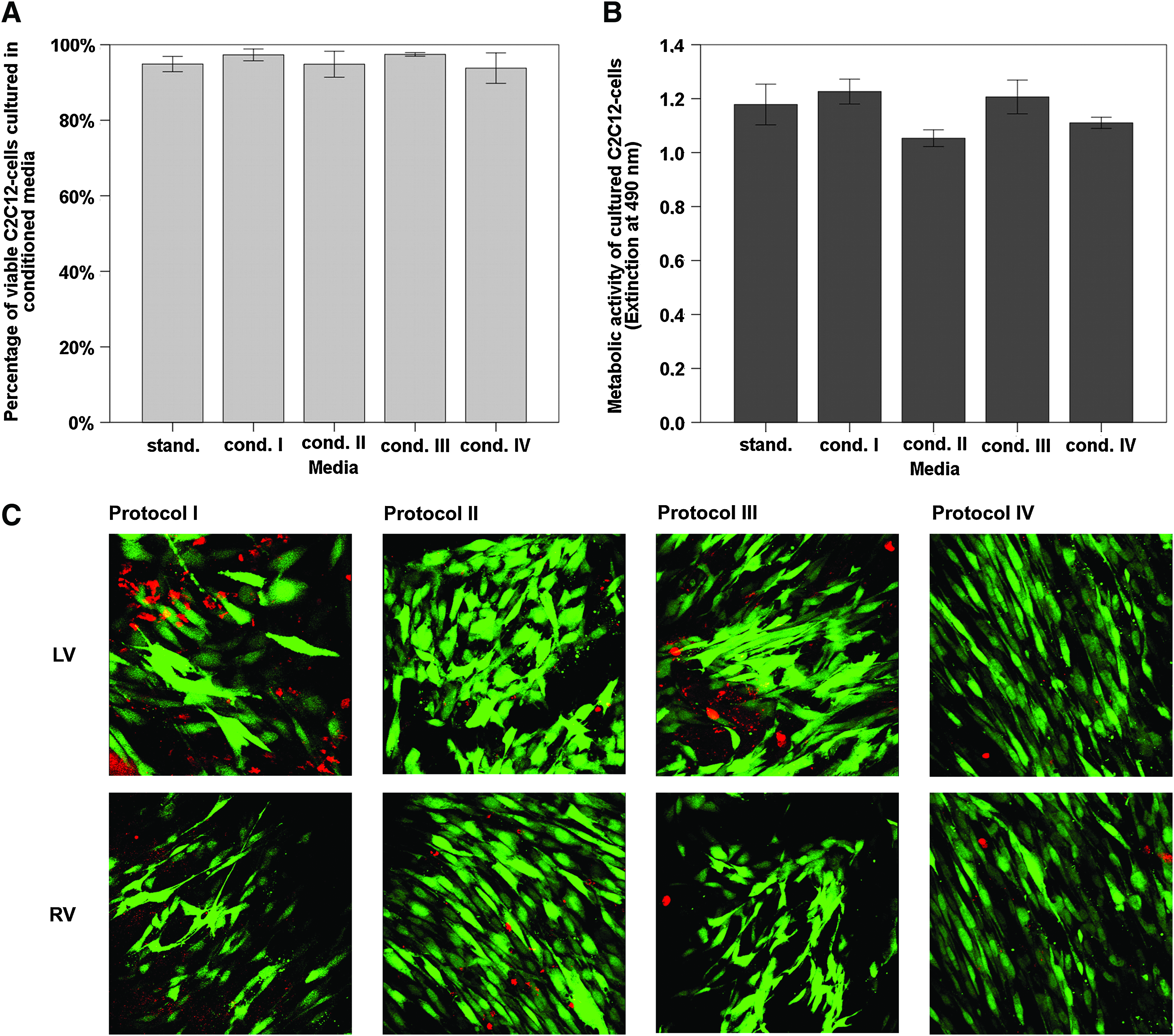
Cytotoxicity and suitability for de novo cellular repopulation of myocardial ECM after whole-heart decellularization through software-controlled automatic coronary perfusion. Differentiated myoblasts (C2C12 cells) incubated for 24 h in conditioned (12 h perfusion through decellularized heart after decellularization) and standard culture media showed no difference in cell viability and proliferation. Further, decellularized scaffold specimens from the left and right ventricular free wall could successfully be repopulated with C2C12 cells demonstrating high cell viability and organization forming intercellular networks inside the ECM after 24 h.
Finally, the suitability of the different myocardial ECM scaffolds for a de novo repopulation was assessed by a direct in vitro seeding of differentiated C2C12 myoblasts onto 100 mm2 large decellularized myocardial ECM specimens of the left and right ventricular free wall, which were obtained after whole-heart decellularization according to the individual protocols. After 24 h of culture cell, calcein-AM and ethidiumbromid staining showed high cellular viability and unremarkable cell morphology. ECM penetration and cell migration inside the scaffold specimens was observed in myocardial ECM with no differences that could be linked to the respective protocol used to produce the scaffolds (Fig. 7C). After 24 h the seeded myoblasts were already highly elongated and organized in aligned strands inside the 3D ECM of all groups, although particularly 3D cultures in ECM scaffolds of protocol IV appeared as highly aligned tissue surrogates, with almost all myoblasts found elongated and aligned in one major direction.
Discussion
Decellularization of native donor organs experiences an increasing level of attention and popularity as a procedure for the creation of an organ-specific extracellular environment for studies in the fields of regenerative medicine and stem cell biology. Recently, studies involving decellularized ECM of different organ systems have been reported, including intestine, urogenital tract, lung, liver, skeletal muscle, and myocardial tissue.8,9,11,15–17 Data on the results of the decellularization of heart valves, which have been extensively exploited as one of the first tissue types when decellularization became popular more than a decade ago, indicate that minor changes in the applied decellularization protocol may lead to major changes of the biophysical or the biochemical characteristics of the decellularized ECM product. These differences of the ECM characteristics are of greater importance when in vitro studies involving more sensitive and delicate cell types are proposed, e.g., stem or progenitor cells. Reviewing the contemporary literature, we found reports on three different protocols with a successful decellularization of myocardial or muscular tissue. In our search for an ideal protocol for myocardial decellularization, we performed a direct comparison of these three protocols on a histological and biochemical level as well as in terms of their biocompatibility for 3D in vitro studies. In addition, we introduced a novel, fourth protocol, which was specifically developed for myocardial decellularization via coronary perfusion and aimed at the employment of additional acting agents other than those in the three previously established protocols.
As a major finding of this study, none of the analyzed protocols proved as an entirely ideal treatment to achieve an optimal decellularized biological matrix, that is, a scaffold that is free of donor cell material along with preserved extracellular components. Moreover, the performance of the analyzed protocols varied considerably depending on the type of applied tests. Likewise, each protocol presented as more powerful or less suitable, depending on the specific matrix quality that was tested, such as macroscopic appearance, degree of donor cell removal, extent of biochemical conservation of collagenous ECM, and retention of GAG. However, these differences remained widely obscured upon biocompatibility studies using differentiated murine C2C12 myoblasts.
For the experiments presented here, we adopted the principle of coronary artery perfusion via cannulation of the ascending aorta, a procedure that was described by Ott et al. for an effective myocardial exposure to the decellularization treatment. In order to exclude perfusion-related differences in tissue treatment and decellularization results, all hearts were decellularized under the same pressure conditions using a software-controlled perfusion system. This way, we sought to create the basis for a direct comparison of the biochemical combinatory effects of the employed solutions rather than physical effects that might be exerted by the different perfusion pressures and varying perfusate volumes as described in the original first reports.8,9,11 Interestingly, protocols I and II, which were considerably shorter than protocols III and IV, resulted in an improved preservation of GAG on the cost of higher remaining DNA values, but only protocol I was able to protect both basement membrane proteins, collagen type IV and laminin. In direct comparison of protocols I and II, elastin, another pivotal component of the cardiac ECM, was better preserved by protocol I, accompanied, however, with increased cellular remnants as determined by histology and colorimetric quantification. We suggest that these differences may be attributed to the enzymatic action of trypsin in protocol II. Trypsin is known to have a nonspecific degradation effect on the ECM. However, trypsin is occasionally used for particularly challenging tissues due to its robust action on cell–matrix junctions. Cardiac ECM that was generated by either one of the short protocol I or II displayed signs of considerable amounts of retained cellular material, as evident in histology by two different stainings as well as in immunolabeling for α-actin. The apparently good results of protocol II when a colorimetric determination of PSR and FG was performed (Fig. 4) have to be evaluated in the context of the considerable loss of total collagen type I content, which was significantly more pronounced than observed with any other protocol (Fig. 5D).
In contrast, the two long-lasting protocols, III and IV, resulted in an improved outcome in terms of cell removal evident, both upon histology and upon biochemical demonstration of a highly significant reduction of DNA throughout all different cardiac regions, even in the apical regions and the septal wall. The qualitatively clear histological appearance of the ECM products of protocol IV, for example, in H&E and Movat staining, was paralleled by best cell alignment when myoblasts were seeded on the decellularized ECM scaffolds. Notably, cell alignment is one of the general crucial endpoints for in vitro tissue culture systems. Hence, numerous previous efforts have been undertaken to direct cell morphology and thereby cell function, particularly in the context of premature or mature cardiac cells. 3 Upon direct cell seeding of myoblasts on myocardial ECM decellularized by protocol IV, we observed a high degree of spontaneous alignment by laser scanning microscopy. These findings were particularly surprising as the procedures described by protocols III and IV represent extended treatments with rather powerful chemical compounds.
On the other hand, specific proteins, such as laminin and collagen type IV, have been reported to impact on the fate of cardiomyocytes and their progenitors. 18 Therefore, the presence of these two proteins, as prominent surrogates of an intact basement membrane, was analyzed and resulted in diverse findings for each protein. Both long-term protocols (III and IV), but also the short-term protocol II, failed to conserve laminin, and only the short treatment by SDS and Triton-X-100 (protocol I) delivered convincing results. A different picture was observed for collagen type IV, where only protocol II failed. A third quality ranking was constituted when elastin content was measured and demonstrated a superior performance of the short protocols I and II. The potency of various ECM proteins, particularly that of elastin and basement membrane constitutes, e.g., laminin and collagen type IV, has been convincingly demonstrated in previous studies. 19 However, these studies have mostly focused on single components of the ECM. Here, we demonstrate a multifaceted character for four different decellularized myocardial scaffolds. Furthermore, passive biomechanical properties, as dominated by ECM components like elastin and the ground substance GAG, may have an impact on cell differentiation pathways and influence the outcome of an in vitro engineered cardiac construct. 20 Moreover, the fate of a decellularized ECM scaffold with enclosed repopulated cells is dependent on immunological interactions that may be provoked by residual donor cell components, such as DNA, cell membrane fragments, or cytoskeletal proteins, when the construct is implanted into an allogenic recipient. Based on our findings it may be suspected that in the context of 3D cell cultures, each of the analyzed specific scaffolds may induce a unique complex of cell–ECM interactions, thereby triggering a distinct cell behavior in vitro, as well as in vivo. In this scenario, favorable and undesired characteristics of a single ECM scaffold may act in an antagonistic manner, for example, remnant donor cytoplasmatic and nuclear components vs. high degree of basement membrane preservation. Future studies involving decellularized ECM-based progenitor or stem cell culture may further elucidate the appropriateness of individual decellularization protocols for the in vitro creation of a desired cell differentiation niche.
The presented study suffers from a number of limitations, including the range of analyzed matrix characteristics. Further analysis of ECM fragmentation and breakdown products are necessary to further delineate the impact of ECM loss in contrast to the impact of present ECM debris on cell behavior. Also, a thorough analysis of ECM-bound growth factors after decellularization is needed, as similar studies on decellularized hepatic ECM has proven a wide range of bioactive components still present in the decellularized matrix. Finally, a single chemical component analysis may improve our understanding on the action of individual detergents and enzymes that are typically used in a cocktail of acting agents.
In conclusion, after successful decellularization, the resulting ECM scaffolds present profound differences regarding actual freedom of cellular components and preservation of crucial ECM proteins, with a changing performance level regarding different scaffold characteristics. By the addition of a further detergent class, for example, saponin, to an already-established protocol, a further gain in decellularization may be achieved, although the general effects on the ECM were partially also accentuated. Therefore, a detailed analysis for each individual experimental setup is mandatory and should be preferred over the simple adoption of a previously as successful reported protocol, particularly in experiments involving cell entities with specific basement membrane requirements.
Footnotes
Acknowledgments
The authors are thankful to R. Eurich and A. Wiss (University of Heidelberg) for their excellent technical support. The authors are also grateful for the generous support of the Dietmar Hopp Foundation. The collaborative support of engineo GmbH (Kelkheim/Germany) for engineering and software programming during the development of the automated perfusion system is highly appreciated. Financial support was provided through a research grant from the Dietmar Hopp Foundation (St. Leon Rot, Germany).
Disclosure Statement
Dr. K.-H. Preuss is co-founder and co-owner of the engineering company engineo, which provides commercial software and engineering products.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.