
Abstract
When the chondrocytes are isolated from the native cartilage and proliferate in vitro, they soon lose their original ability to express glycosaminoglycan (GAG) and type II collagen, which is termed dedifferentiation, or decrease cell viability. We first examined in vitro cartilage regeneration of tissue-engineered pellets that consisted of human auricular chondrocytes and atelocollagen and that were incubated in vitro under stimulation with bone morphogenetic protein-2 (BMP-2), insulin, and T3. We then examined the administration of those growth factors into the scaffold or in the medium and explored the possibility that the atelocollagen, the hydrogel scaffold of the chondrocytes, may function for drug delivery of the factors. BMP-2 in the atelocollagen with the supplement of insulin and T3 in the medium could not only produce a greater GAG matrix in a shorter period but also sustain cell viability with lower mortality. The insulin in the medium could be better administered only for 2 weeks, rather than 3 weeks, which would save time and cost, hence shortening the in vitro culture of chondrocytes. Our protocol of mixing BMP-2 into the atelocollagen with the supplement of insulin and T3 hormone might provide a new insight into the development of tissue engineering in chondrogenesis.
Introduction
Nowadays, there are two directions for tissue engineering, in situ tissue engineering (ACI) and highly-maturated organoid (the new trend). In situ tissue engineering simply administers cultured cells into the lesion and maximizes the use of in vivo regeneration. Highly maturated organoid enhances the maturation of tissues in vitro and reduces the risk for in vivo changes in morphology and functions. Highly maturated organoid is a pivotal direction that improves that quality of engineered tissues. For that, the development for in vitro maturation of engineered tissues is the major task.
Therefore, we should induce the redifferentiation of the chondrocytes suffering from dedifferentiation that inevitably occurs during the proliferation of culture. Some previous papers described trials in enhancing the redifferentiation of dedifferentiated chondrocytes by the inhibition of the extracellular signal-regulated kinase (ERK) signal 8 or an increase in hydrostatic pressure,9–11 neither of which may be predictable due to cytotoxicity or economic reasons. We previously attempted to optimize the combination of growth factors that have already been in clinical use for stimulation of redifferentiation and found that the combination of bone morphogenetic protein-2 (BMP-2), insulin, and triiodothyronine (T3) strongly enhanced the redifferentiation or the production of cartilage matrices when the dedifferentiated chondrocytes embedded within atelocollagen hydrogel were three-dimensional (3D)-culutred in the medium containing these factors. 12 However, the 3D culture of the tissue-engineered constructs in the medium, if the construct becomes larger, principally includes problems in which the growth factors contained in the medium hardly reach the middle part of such a large construct and sufficient exchanges of oxygen and nutrition cannot be achieved. Due to these difficulties, the 3D-cutlure of the tissue-engineered constructs immersed into the redifferentiation medium may not be applied in practice. Therefore, the purposes of the present study were to increase cell viability and cartilage maturation of the tissue-engineered constructs by optimization of the place or the timing in the administration of growth factors.
In the first part of the present study, by the administration of the growth factors into the scaffold, instead of the medium, we attempted to improve the access of growth factors to the cells located in the middle part of the constructs, so as to enhance the maturation of the tissue-engineered constructs. For that, we examined the size effect on the in vitro cartilage regeneration of tissue-engineered pellets that were incubated in vitro under stimulation with BIT (B: BMP-2, I: Insulin, T: Triiodinothyronine). We then explored the possibility that the atelocollagen, the hydrogel scaffold of chondrocytes, 13 may function for drug delivery of the growth factors. We mixed high concentrations of the growth factors with the atelocollagen hydrogel and embedded the chondrocytes within it for effective stimulation of the cells located in the middle part of the tissue-engineered constructs. Cartilage regeneration of the constructs was evaluated by using this hydrogel in the in vitro experiments.
In addition, reducing prohibitively expensive growth factors used in the 3D culture might be another concern in the tissue engineering for the cartilage. It is well known that cells, growth factors, and a scaffold are the three essential parts in tissue engineering. Here, we would also like to address the “place for the growth factor” and “the time and duration of the growth factor” and attempted to reduce the consumption of time and expense and, hence, to achieve the practical clinical application of the tissue-engineered cartilage. If we could determine the best timing or the proper duration for the administration of the growth factors, we would make the procedure for tissue engineering more efficient and reduce the consumption of the expensive growth factor, hence we could make tissue engineering of the cartilage possible and practical in clinical application.
Materials and Methods
Human chondrocyte preparation
All the procedures of this study were approved by the ethics committee of the University of Tokyo Hospital (ethics permission number 622). Human cartilage was obtained from remnant auricular cartilage of microotia patients who underwent surgery at the University of Tokyo Hospital. After cartilage had been digested in 0.15% collagenase solution for 24 h, isolated chondrocytes were seeded on a 10-cm plastic tissue culture dish in Dulbecco's modified Eagle's medium (DMEM)/F12 containing 5% human serum supplemented with fibroblast growth factor-2 (FGF-2, 100 ng/mL) and insulin (5 μg/mL), as we previously reported. 14
Three-dimensional culture of tissue-engineered cartilage pellets
The chondrocytes proliferated at a 1000-fold increase in numbers in monolayer and then were suspended in 0.8% atelocollagen solution (Kawaken) in MEM at a density of 107 cells/mL. Different volumes of the cell-atelocollagen suspension were placed into the bottom of a 15-mL polypropylene conical tube (BD Falcon) to form a gel in 2-h incubation at 37°C. The DMEM/F12 medium (Sigma-Aldrich) was used at a volume of 2 mL for each gel and cultured in a 37°C/5% CO2 incubator. To induce redifferentiation of the chondrocytes, insulin (MP Biomedicals), recombinant human bone morphogenetic protein-2 (BMP-2; kindly provided by Astellas Pharma, Tokyo, Japan), or 1, 5, 3′-triiodothyronine (T3; EMD Bioscience) were added to the medium, according to the previous report. 15 For the experiments in which the growth factors were mixed within the atelocollagen hydrogel, those factors were added to 1% atelocollagen solution and thoroughly mixed within it, followed by gelation at 37°C for 2 h in the 15-mL conical tubes, and a 2 mL addition of the medium. Throughout the experiments, the medium was changed once every week.
Designating the pellets with the combination of growth factors in a scaffold or in the medium by using a sequence coordinate system
Here, we describe the placement of added growth factors with a sequence coordinate system (Fig. 1, in the order of BMP-2, insulin, T3). We used A to designate atelocollagen, while M stood for medium and O for null (neither in the medium nor in the atelocollagen). We used AAA to denote all the growth factors of BMP-2, insulin, and T3 placed into the atelocollagen, and, hence, MMM for all the growth factors in the medium. The control group is designated by OOO. For example,
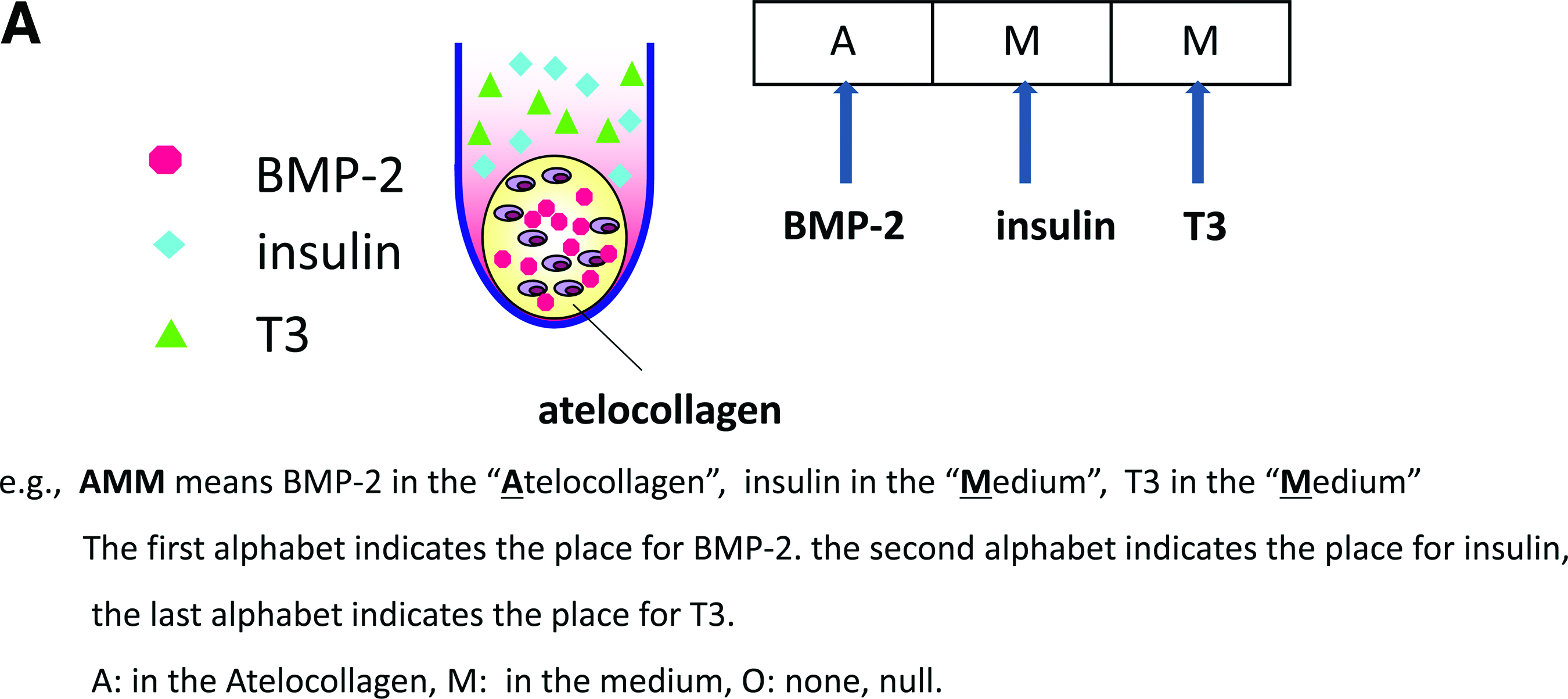
Nominating system.
Cell numbers and viability
A ChemoMetec NucleoCounter was used to measure the total cell numbers, and the living cell numbers were measured by calculating the Propidium iodine-stained nuclei of the cells. In tissue-engineered cartilage pellets, the cells were recovered by the digestion in 0.3% collagenase for 1 h at 37°C. Cell viability was defined as the living cell number divided by the total cell number. 16 Cell mortality was defined as the dead cell number divided by the total cell number.
Histology
The tissue-engineered cartilage was fixed with 4% paraformaldehyde, embedded in OCT compound (Sakura Finetek Japan), and cryosectioned into 6-μm slices. The sections were stained with toluidine blue.
Real-time reverse transcriptase–polymerase chain reaction analysis
Total RNA was isolated from chondrocytes with ISOGEN (Wako Pure Chemical Industries) following the supplier's protocol. Complementary DNA (cDNA) was synthesized from 1 μg of total RNA with the PrimeScript reverse transcriptase (TAKARA BIO Inc.). The full-length or partial-length cDNA of the target genes, including polymerase chain reaction (PCR) amplicon sequences, was amplified by PCR, cloned into pCRTOPO Zero II or pCR-TOPO II vectors (Invitrogen), and used as standard templates after linearization. With SYBR Green PCR Master Mix (Invitrogen), real-time fluorescence detection was performed with an Applied Biosystems 7500 Real-Time PCR System (Applied Biosystems) with the following protocol: initial denaturation for 10 min at 94°C followed by 40 cycles consisting of 15 s at 94°C and 1 min at 60°C. All reactions were run in quadruplicate. Sequences of the primers were 5′-CTCCTCGCTTTCCTTCC TCT-3′ and 5′-GTGCTAAAGGTGC CAATG GT-3′ for type I collagen αI chain (COL1A1); 5′-GAGTCAAGGGTGATCGT GGT-3′ and 5′-CACCTTGGTCT CCAGA AGGA-3′ for type II collagen αI chain (COL2A1); 5′-GAAGGTGAAGGTCGGA GTCA-3′ and 5′-GAAGATGGTGATGGGATTTC-3′ for GAPDH. 17
Photometry analysis of ssDNA apoptosis enzyme-linked immunosorbent assay
Cells were obtained from the atelocollagen pellets by digestion in the 0.5% collagenase for 30 min at 37°C. Cell numbers of the cell suspension solution were calculated by trypan blue staining under microscopy. Five thousand cells were then transferred into each well of a 96-well microplate, and the microplate was then centrifuged at 200 g for 5 min. Medium was then removed and to each microwell, 200 μL fixative was added (80% methanol in phosphate-buffered saline). Then, the microplate was incubated at room temperature for 30 min. The fixative was removed, and the microplate was kept at room temperature for 1–2 h to allow attachment of the cells to the plate. The cell apoptosis was evaluated according to the protocol of the ssDNA Apoptosis enzyme-linked immunosorbent assay (ELISA) Kit (CHEMICON® International, Inc.). The absorbance of ELISA was measured in a standard microplate reader at 405 nm.18,19
Dose response of growth factors for cell viability
A cell suspension (20,000 cells) of 100 μL DMEM/F12 was prepared in the density of 200,000 cells/mL. The cell suspension (100 μL) was then added to each well of a 96-well plate. The microplate was preincubated at 37°C. To the well, 50 μL of various concentrations of BMP-2, insulin, and T3 in different dilution concentrations was added. The plate was placed in the incubator at 37°C for 24 h. CCK-8 solution (Dojindo) of 10 μL was added into each well, and the plate was placed in the incubator at 37°C for 1 h. 20 The optical density of the plate was read at 450 nm to indicate the cell viability in each well. The optical density (OD) values were adjusted by those of standard cell samples, and the relative ratio of the OD values was used for comparison.
The biochemical measurement of GAG
The samples were dissolved in 10 mg/mL pepsin/0.05M acetic acid at 4°C for 48 h and then in 1 mg/mL pancreatic elastase/0.1 M Tris-0.2 M NaCl-5 mM CaCl2 at 4°C overnight. The samples were centrifuged at 9100 g for 5 min to remove the residue. The sulfated GAG content was measured by using Alcian blue binding assay 21 (Wieslab AB). GAG in the supernatant was precipitated with Alcian blue solution, and the sediments obtained by centrifugation at 6000 g for 15 min were dissolved again in 4M GuHCl-33% propanol solution. The spectrophotometrical absorbance of the mixture was measured at a wavelength of 600 nm.
Statistical analysis
We prepared nine constructs and measured each sample. Data were expressed as means±standard deviation. Statistical significance was evaluated using analysis of variance or the student t-test for comparison with SPSS software. p<0.05 was interpreted to denote statistical significance.
Results
Size effect on cartilage regeneration in vitro
First of all, we examined the size effects on matrix synthesis and cell viability in tissue-engineered cartilage pellets incubated in vitro for 3 weeks, with the redifferentiation medium (BMP-2 200 ng/mL, insulin 5 μg/mL and T3 100 nM, MMM, labeled as BMP-2, insulin, and T3 in the medium) or without any factors (OOO as control group). Three different sizes of pellets were made, 5, 20, and 100 μL for the MMM and OOO groups. In the histological findings with toluidine blue staining, the pellets of 5 μL in the MMM group showed homogenous metachromasia, suggesting the accumulation of cartilage matrices, although in those of more than 20 μL, only the surface areas exhibited metachromasia. The pellets of control groups OOO did not contain any metachromasia (Fig. 2A). The decrease in matrix accumulation due to the increase in pellet sizes suggested the risk for central necrosis in the large tissue-engineered constructs under the in vitro 3D culture and deterioration of the cartilage regeneration. Comparison was mainly focused on the same size pellets between the MMM and OOO groups. The total cell numbers and dead cell numbers of the pellets were measured by using a NucleoCounter, which revealed that there was no significant difference between the MMM and OOO groups, within the same-size pellets (Fig. 2B). The mortality of the cells between MMM and OOO in the 5 μL and 20 μL pellets was significantly different, while there was no significant difference between the mortality of MMM and OOO in the 100 μL pellets (Fig. 2C). The combination of the growth factors BMP-2, insulin, and T3 could not cause any difference in mortality in the large-sized pellet. We then evaluated the gene expressions of COL1A1, a marker of chondrocyte dedifferentiation, and COL2A1, that of differentiation, in the samples by real-time reverse transcriptase–PCR. The relative ratio of COL1A1 expression showed the highest in the 5 μL pellets of the OOO group (Fig. 2D), although the pellets of the same size in the MMM group significantly expressed COL2A1 (Fig. 2E).
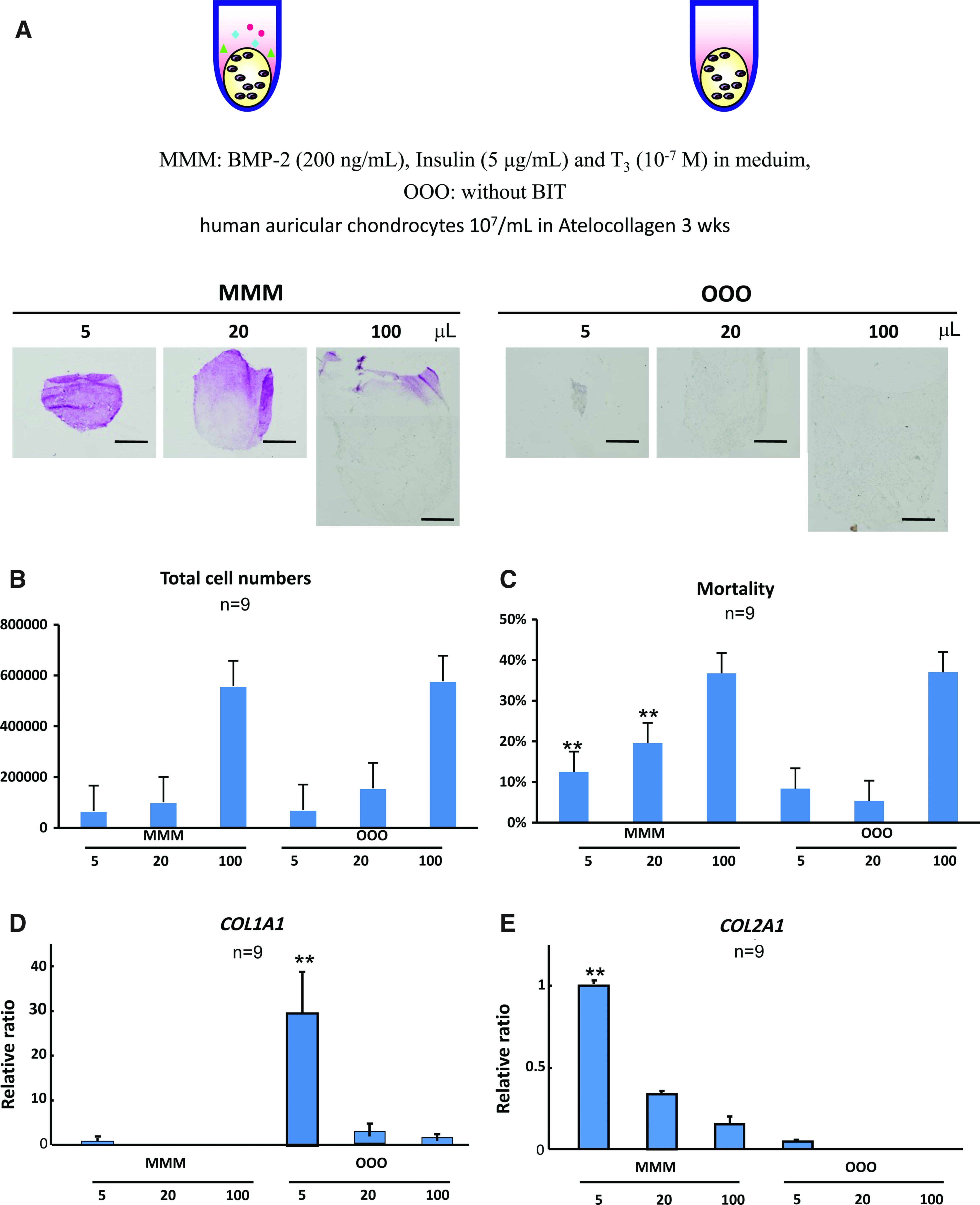
Size effect on cartilage regeneration in vitro.
This depicts the effect of the growth factor BMP-2, insulin, and T3 on increasing the GAG production and also reflected the difficulty in making large pellets of tissue-engineered cartilage even though we manipulated the growth factors. Since the critical size of pellets for central necrosis seemed between 5 and 20 μL due to those data, the 10 uL was used in the experiments thereafter.
The growth factors mixed within the atelocollagen hydrogel
In order to prevent the deterioration of matrix production and chondrocyte viability in the middle part of the larger pellets, as seen in Figure 2, we explored the possibility of mixing the growth factor within the atelocollagen hydrogel. Within the atelocollagen hydrogel, we mixed large amounts of the growth factors that were equivalent to those contained in the medium of 2 mL, which was the volume our lab usually used in the protocol for 3D culture of the tissue-engineered cartilage pellets. The amounts of growth factors were 400 ng for BMP-2, 10 μg for insulin and 200 pmole for T3.
In the histological findings of the AAA pellets stained by toluidine blue, metachromasia was present, while that of MMM also showed the accumulation of cartilaginous matrices (Fig. 3A). No metachromasia in the OOO pellets was observed even under high magnification (Fig. 3A). The apoptosis analysis shows that the AAA seemed lower in apoptosis, followed by MMM and by the OOO (Fig. 3B). The cell counts showed that AAA and MMM pellets tended to manifest a large total cell number, although the control group showed less. The MMM group was noted to have the highest total cell number among the three groups (Fig. 3C). The OOO group shows the lowest numbers of total cells. The cell mortality (Fig. 3D) detected from the NucleoCounter was consistent with the apoptosis analysis (Fig. 3B). Either AAA or MMM was different from the control groups in mortality, while there was no significant difference between AAA and MMM.
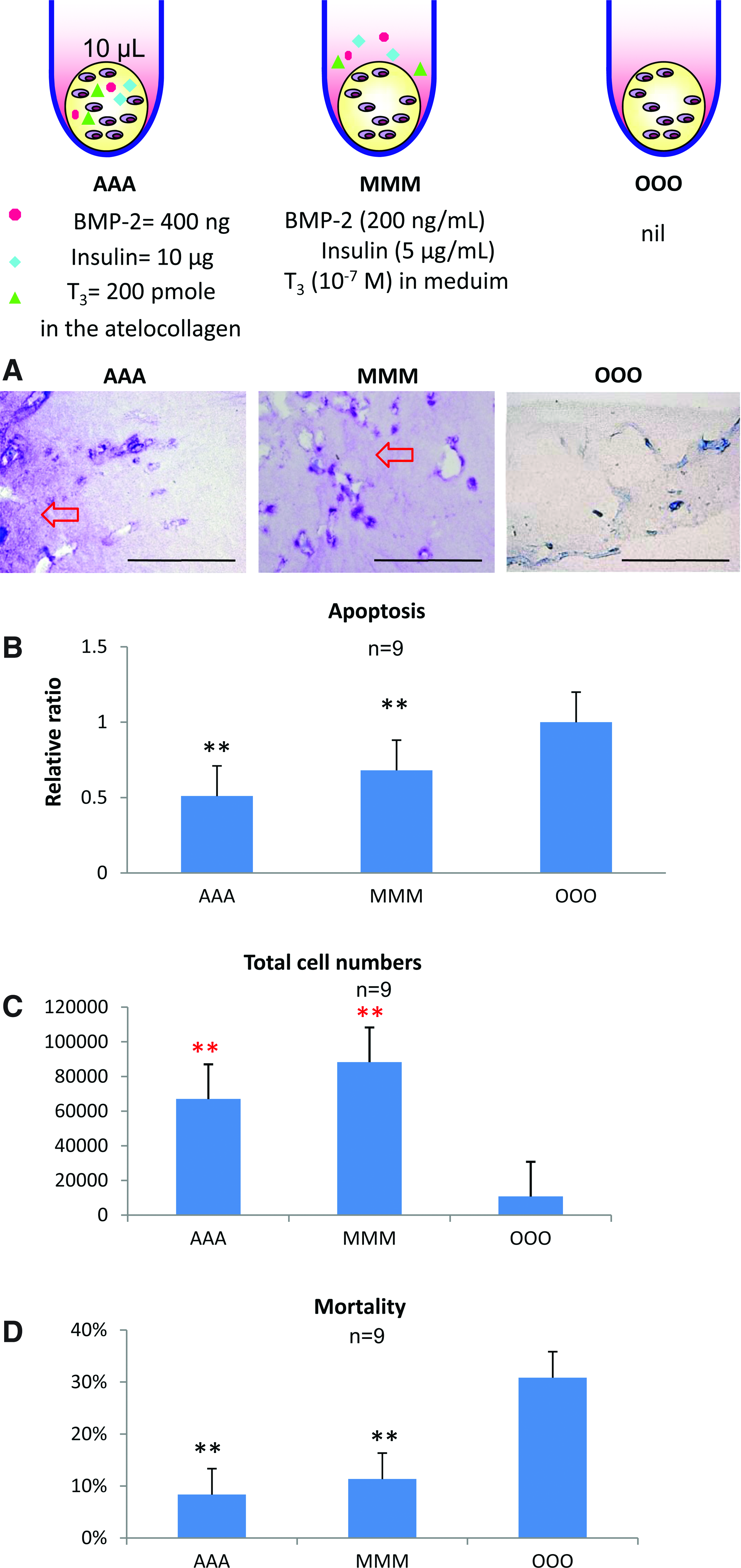
The growth factors mixed within the atelocollagen hydrogel. The tissue-engineered cartilage pellets of 10 μL were cultured in vitro for 2 weeks.
These findings suggest the possible superiority of AAA with MMM over the OOO group and encourage us to manipulate the growth factors into the medium for 3D culture in vitro of chondrocytes.
Twenty-seven groups in 3D culture and cytotoxicity of BMP-2, insulin, and T3 hormone
For detailed analyses of the effects of each growth factor within the scaffold atelocollagen hydrogel or within the medium, we measured the GAG contents in the pellets, in which BMP-2, insulin, or T3 was placed into the medium or into the atelocollagen hydrogel. We prepared 27 groups in which three factors (BMP-2, insulin, and T3) were treated using 3 options (in the medium, in the atelocollagen gel, and no administration) for each growth factor and incubated those pellets for 3 weeks. In the 27 groups, we noticed that BMP-2 and insulin take the major role in the production of GAG. AAO, AMO, OMO, MMO, OMM, and AMM showed high GAG production (Fig. 4A). Regarding those six groups, three groups had BMP-2 embedded in the atelocollagen (AAO, AMO, and AMM), while insulin was found in the medium of five groups (AMO, OMO, MMO, OMM, and AMM). T3 seemed to negatively function in matrix production when it was placed in the atelocollagen (Fig. 4A, XXA, X=O, M, or A). Insulin either in the medium or in the scaffold tended to increase the GAG content (Fig. 4A, XAO or XAM, XMO or XMM, X=O, A, or M).
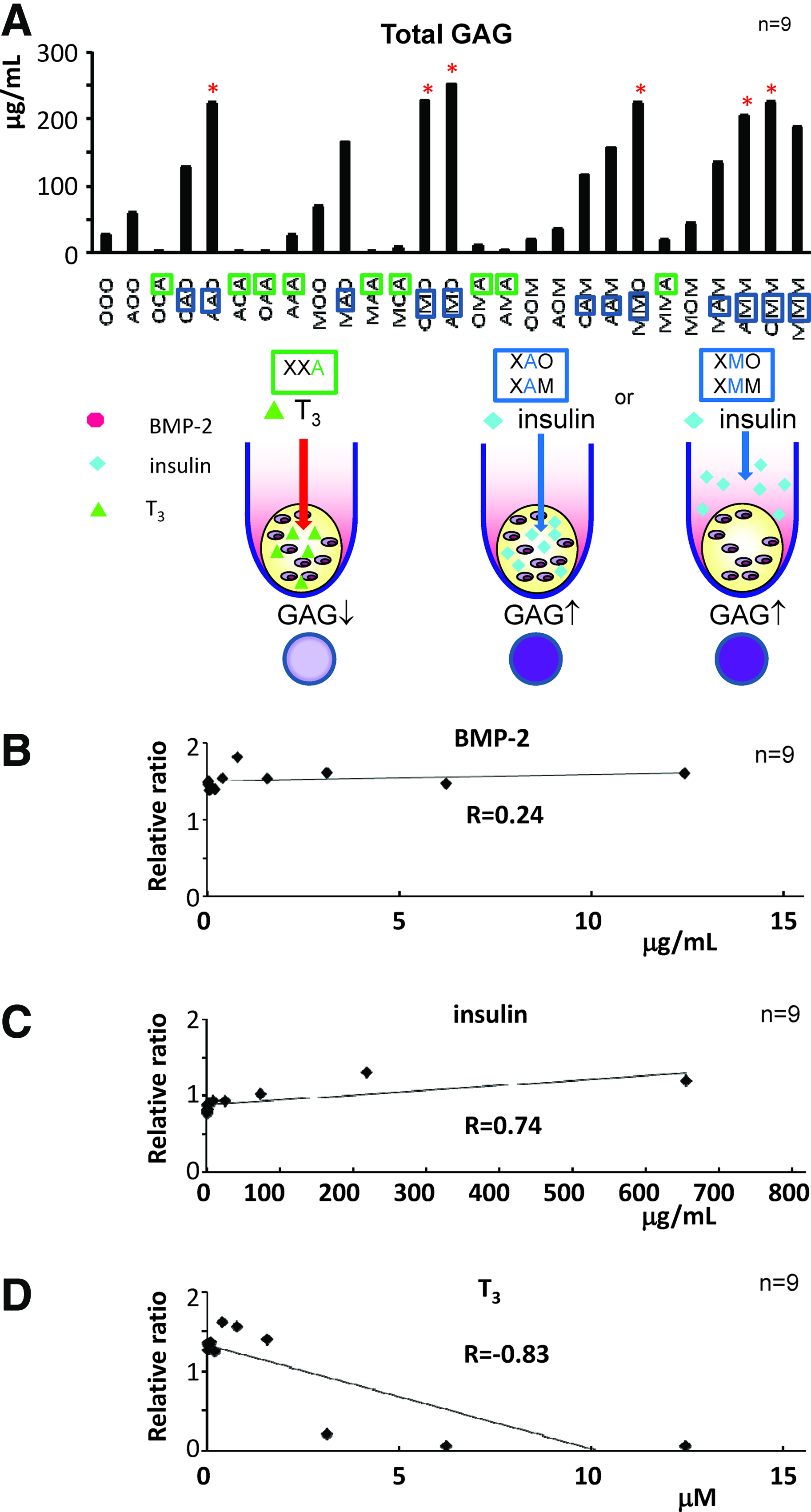
Twenty-seven combinations and cytotoxicity and dose response of the growth factors.
In order to evaluate the cytotoxicity of each growth factor at a high concentration, dose-response experiments of BMP-2, insulin, and T3 for the cell viability were performed. As a result, BMP-2 showed no obvious influence on the cell viability in the range of approximately 10 μg/mL (R=0.24. Fig. 4B), while cell viability was slightly promoted by an increase in the insulin concentration (0–600 μg/mL, R=0.74, Fig. 4C). However, higher doses of T3 than 2 μM significantly enhanced cell mortality (R=−0.83 Fig. 4D), supporting the results that the atelocollagen containing a high concentration of T3 showed lower GAG content (XXA, X=M or A) in Figure 4A.
These data indicate that the T3 in the atelocollagen is harmful to the chondrocyte with minimal GAG production, though T3 in the medium could still promote GAG production. Either in the medium or in the atelocollagen, insulin could induce production of the cell numbers and enhance the GAG production.
The best 6 combinations among 27 groups
Based on the consistent results of 27 combinations of growth factors and their cytotoxicity investigation, we excluded the possibility of placing T3 into the scaffold in the 3D culture of chondrocytes. To reduce the number of candidate combinations and to explore the possible mechanism, we focused on the best 6 groups of pellets among 27 combinations at the 3-week 3D culture of tissue-engineered cartilage (Fig 4A). The best six groups were as follows (Fig. 5A): AAO (BMP-2 and insulin in the atelocollagen), AMO (BMP-2 in the atelocollagen and insulin in the medium), OMO (insulin in the medium), MMO (BMP-2 and insulin in the medium), OMM (insulin and T3 hormone in the medium), and AMM (BMP-2 in the atelocollagen, insulin and T3 hormone in the medium). The macroscopic pictures of six groups showed no major difference (Fig. 5A). The total cell number of AAO (Fig. 5B) was observed to be the least and showed a significant difference from that of the other five groups (p<0.01), suggesting a limited proliferating effect of insulin in the atelocollagen. There was no significant statistical difference among the mortality of the six groups, but we might still notice the lowest mortality of OMO (insulin alone in the medium) and the higher mortality found in the pellets with BMP-2 (AAO, AMO, and AMM) (Fig. 5C). From the perspective of GAG production (Fig. 5D), BMP-2 was better placed in the atelocollagen in the presence of insulin in the medium (AMO>MMO, p<0.05), while the GAG production of AMM was significantly higher than that of MMO (AMM>MMO, p<0.05), revealing the importance of the synergic action of the three factors and their important role in the “suitable place.” The value of the AMM tended to be higher than that of the AMO, though they showed no statistical difference. The histological findings of the six groups stained by toluidine blue (Fig. 5E) showed a high degree of metachromasia in the AMO and AMM groups and reflected the measurement of GAG production (Fig. 5D).
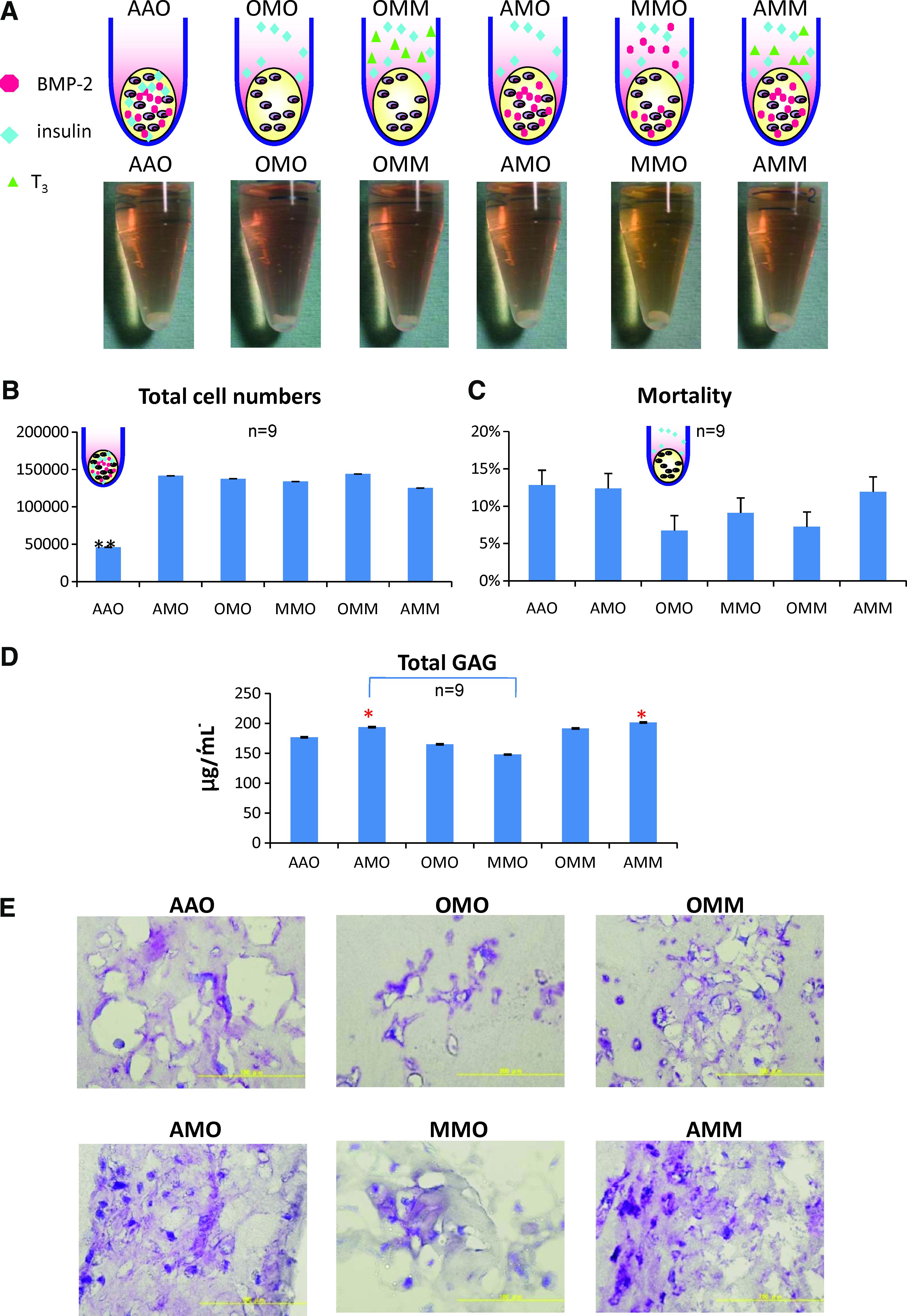
The best 6 combinations among 27 groups. As to find out the best combinations and to explore the possible mechanism, we focused on the best 6 groups of pellets at the 3-week, 3D culture of tissue-engineered cartilage (Size: 10 μL). The best 6 groups were as follows: AAO (BMP-2 and insulin in the atelocollagen), AMO (BMP-2 in the atelocollagen and insulin in the medium), OMO (insulin in the medium), MMO (BMP-2 and insulin in the medium), OMM (insulin and T3 hormone in the medium), and AMM (BMP-2 in the atelocollagen, insulin and T3 hormone in the medium).
Time-specific administration of insulin in the medium for BMP-2 in the atelocollagen
Since we had the results of a good combination of BMP-2 in the atelocollagen and insulin in the medium (AMO) in Figure 5D and Figure 5E, we would like to determine the time influence of insulin on the expression of the tissue-engineered chondrocytes of AOO within the period of 3 week in vitro culture. Insulin (5 μg/mL) was added in the medium for the 10 μL AOO pellets at the different time points to investigate the influence of the time factor with the administration of insulin on the chondrocytes in the BMP-2-containing atelocollagen. The group of the 1st week showed the weakest GAG production (Fig 6A) among the 1-week administration groups (1st week < 2nd week, 1st week <3rd week, p<0.01). The group (1st week + 2nd week) showed great value (318.8 μg/mL) in GAG production among all the groups, without any significant difference from that of the 3-week administration group (1st week +2nd week +3rd week) (315.4 μg/mL) (Fig. 6A). This indicates that placing BMP-2 into the atecollagen could achieve a high GAG production with insulin supplemented in the medium before the end of the 2nd week in vitro culture.

Time-specific administration of insulin in the medium for BMP-2-embedded 3D culture (AOO pellet + insulin in the medium). Since we had the results of the good combination of BMP-2 in the atelocollagen and insulin in the medium (AMO) in Fig 5-D & Fig 5-E, the time influence of insulin on the expression of the tissue-engineered chondrocytes of AOO (Size: 10 μL) within duration of 3 weeks was also determined. We added the insulin (5 μg/mL) in the medium for the AOO pellets at the different time point to investigate the influence of time factor on the chondrocytes.
The control group labeled as nil showed the smallest total cell numbers, compared with the other seven groups (Fig. 6B), indicating the necessity of insulin in the medium for the chondrocyte pellets with BMP-2 embedded in the scaffold atelocollagen. Additional results also supported this hypothesis; there was no significant difference between any two groups of the other seven groups with insulin, indicating that no matter when or for how long the insulin is administered into the medium for the pellets embedded with BMP-2 within the atelocollagen (AOO), insulin could proliferate the number of chondrocytes (Fig. 6B). The control group labeled as nil was found significantly higher in mortality than the other seven groups (Fig. 6C), while there was no significant difference between any two groups of the other seven groups with insulin. This result repeats the same tendency in total cell number (Fig. 6B), indicating that once the insulin is administered into the medium for the culturing the pellet AOO (pellet embedded with BMP-2 in the atelocollagen), we could reduce the cell mortality (Fig. 6C). Here, the results re-emphasized the important role of insulin in reducing the mortality in the same tendency by increasing the total cell numbers (Fig. 6B).
The data suggest that the AMO for 2 weeks shows no differences from that of AMO for 3 weeks in terms of total GAG, total cell numbers, and mortality, implying that the shorter culture period is sufficient for in vitro culture. Insulin could increase the total cell numbers and dead cell numbers but still could reduce the mortality of the AOO pellets. That is to say the AMO model is ideal for cell proliferation with reduced mortality.
Discussion
Atelocollagen is the protein in which telopeptides, the main antigenic site of collagen, is removed by the enzyme. The atelocollagen is applied as a tissue filler, a hemostatic agent, or wound dressing in clinical practice. 22 It also functions as a scaffold in cartilage tissue engineering, because the atelocollagen hydrogel allows a gradual proliferation, the matrix production of chondrocytes, and the maintenance of their phenotype.23,24
The atelocollagen hydrogel stands for an useful carrier for the growth factors25–27 or the exogenous genes.28,29 Under the conditions of neutral pH and 37°C, the atelocollagen spontaneously assembles to become fiber like. This indicates that the administered atelocollagen is not immediately absorbed in the living body but remains for some period, which has advantages as a persistent release carrier. The usefulness of the collagen hydrogel as the carrier of drugs was indicated in the applications for IFN-alpha, 30 IL-2,31,32 NGF,33–35 bFGF,2,36–39and hGH. 40 The present article has first shown that the atelocollagen hydrogel also functions as an effective carrier of BMP-2.
BMP-2 plays a key role in the development of bone and cartilage, involving the hedgehog pathway, the TGF-β pathway, and cytokine-cytokine receptor interaction. When BMP-2 is implanted into a collagen sponge, it could induce the formation of new bone and can be adopted for the treatment of bony defects, delayed union, and nonunion.41,42 BMP-2 is also essential in chondrogenesis especially in matrix production.43,44
Since there seemed to be no single mechanism for the controlled release of growth factors, multiple factors including the molecular sizes of factors or the microstructures of atelocollagen may be involved. The molecular weight of BMP-2 is approximately 26 kDa, which is rather large and can be trapped into the microstructure of the atelocollagen hydrogel. We reported that the insulin embedded within the atelocollagen hydrogel continued to be retained for some days, which may be rather short to promote the redifferentiation of dedifferentiated chondrocytes. However, this growth factor rapidly stimulates the intercellular signaling including via P38 and Akt within some minutes, suggesting that it may be sufficient for the commitment of chondrocyte differentiation or redifferentiation. 45
Interaction of BMP-2 and atelocollagen
BMP-2 also has a high affinity for collagen, and Takeuchi et al. has found the recombinant human bone morphogenetic protein-2/atelocollagen compound to be a novel material for ossicular reconstruction.46,47
Suzawa 48 et al. demonstrated that mouse osteoblastic MC3T3-E 1 cells constantly expressed messenger RNAs (mRNAs) for BMP-2 and BMP-4, and the BMPs were accumulated within collagen-rich extracellular matrices by the cells. BMPs enhanced osteoblastic differentiation, mediated by the binding with collagen. BMPs together with the collagen of extracellular matrices participated in the induction of osteoblastic differentiation of nonosteogenic mesenchymal cells and cells in the osteoblast lineage as well.
A similar phenomenon is also noted in chondrogenesis. The dense collagen scaffold applied with BMP and/or TGF-β was found to stimulate the production of extracellular matrix chondroitin by Glowacki et al. 49 Hou et al. 25 also observed the excellent biocompatibility of an rhBMP-2-atelocollagen scaffold with a capacity for osteogenesis and chondrogenesis in rats within 2 weeks. Since the domain I of perlecan (Pln) produces heparan sulfate chains that link and unlink heparin binding growth factors, Yang et al. 50 noted that the Pln domain I (PlnDI) could be compounded with collagen II (P-C) fibrils to enhance linkage of BMP-2, and, hence, the compound supports the formation of cartilage and cartilage-like tissue in vitro. It was proved that P-C fibrils provide an improved biomimetic material for the binding and holding of BMP-2 to support chondrogenic differentiation. It is also noted that polylactic acid (PLA)-based scaffolds coated with P-C fibrils bound more BMP-2 than either PLA scaffolds or those coated with collagen II fibrils alone.
BMP-2 takes on a major role in the initial stage in chondrogenesis within the first 24 h following the fracture. 51 Since BMP-2 is needed for the early stage, mixing BMP-2 into atelocollagen makes sense for it to be highly concentrated in the scaffold.
In chondrocytes, cell viability tended to be decreased when matrix production was enhanced, which may correspond to the physiological finding in which the decrease in chondrocyte viability and the incidence of apoptosis increased in the hypertrophic chondrocytes of the growth plates.12,52–54 Since BMP-2 would decrease cell viability during the progression of hypertrophy, the combination of insulin with BMP-2 could overcome the deterioration in cell viability owing to BMP-2. The mortality of (OOO+BMP-2) was quite high; above 20%, reaching near 40% (data not shown), while the mortality of (AOO+insulin) was decreased to around 10% in Figure 6C. The control group (labeled as nil, without the addition of insulin, i.e., AOO alone) manifested high mortality above 20% (Fig. 6C). This result also depicts the important role in chondrocyte viability or in cell proliferation.
Time course
The time factor experiments involved the groups for different periods of growth factor administration (1-week duration, 2-week duration, 3-week duration, and nil).
BMP-2 plays a key role in the early stage of chondrogenesis and is regarded to function better in the atelocollagen than in the medium, and the continuous administration of insulin and T3 is essential in the GAG production and maintenance of chondrocytes.
Insulin is produced in the pancreas, and the T3 hormone is secreted by the follicular cells of the thyroid gland, and hence, both hormones are sent to the target via the blood supply. BMP-2 is synthesized locally, but the other two growth factors are distributed systemically, which also reflects some implication from our model. Our AMM model, placing BMP-2 in the scaffold for the early reaction and a continuous supply of insulin and T3 hormone (thyronine) in the medium, just repeats the physiology.
The half life of those growth factors is also considered. BMP-2 has a half life of 5–6 days in the joint, while an insulin molecule yielded endogenously by the pancreatic beta cells is degraded in approximately one hour after its initial liberation into circulation (the half-life of insulin is 4–6 min). In comparison with the half time of insulin, that of T3 is only 2.5 days. From the data just provided, BMP-2 has the longest half life, meaning that it is an ideal stable growth factor functioning longer in situ in the scaffold. The shorter half lives of insulin and the T3 hormone indicate better functioning in the medium and a requirement of frequent change in the medium containing the growth factors.
Putative Mechanism of AMM Protocol
As the top 2 groups in GAG production among 27 groups, AMO and AMM have no significant differences though AMM has a higher GAG value. In addition, T3 might be indispensible in chondrogenesis for the tissue-engineered cartilage. At least for AMO, the insulin could better be administered only for 2 weeks instead of 3 weeks, which would save time and cost and, hence, shorten the period of in vitro culture. Thus, our recommendation is AMO or AMM for 2 weeks in vitro culture, though the influence of T3 still requires further investigation. The merit of our protocol using the BMP-2 embedded pellets with a supplement of insulin in the medium is not only for achieving high GAG production within a shorter period but also for greater survival of the chondrocytes even in the presence of insulin.
Synergic reaction of BMP-2 and insulin
Gelse 55 proposed a functional relation between different kinds of chondrogenic growth- or differentiation factors where the mechanism HIF-1α (Hypoxia inducible factor-1α) was found to transactivate Sox9, which is a key transcription factor for many cartilage-specific genes. BMP-2 signaling involves not only the Smad1/5/8 cascade, but also, similarly to IGF-I, stimulates the PI3K/mTOR and the MAPK/extracellular signal-regulated kinase kinase (MEK)/ERK pathway. Especially UO126, a MEK inhibitor, almost completely disabled HIF-1α induction by BMP-2 or IGF-I. Inhibition of PI3K by Wortmannin and inhibition of mTOR by Rapamycin influenced the HIF-1α levels only mildly. IGF-1 and BMP-2 had no effect on the HIF-1α or PHD2 gene expression; that stands in contrast to the effects of TGF-β, which was proved to inhibit PHD2 gene expression via Smad2/3. If we could know more about the mechanism of the associated pathways and could see how BMP-2 shares the common pathways with IGF-I or insulin through the scaffold and the medium, respectively, we would be able to make great progress in the tissue engineering of cartilage. Koay et al. 56 suggest that oxygen availability has remarkable effects on the differentiation and synthetic potentials of hESCs and may have significances for the development of technologies with regard to engineering cartilage.
Collagen offers hypoxic microenvironment and, hence, in which HIF-1α transactivates the sox 9 transcription factor and the following pathway to increase chondrogenesis. Nillesen et al.56–58 noticed that the collagen scaffold containing no FGF2 and vascular endothelial growth factor was found with high density of HIF-1α positive cells (36.6%±9.9%). This also indicates the advantages of the atelocollagen scaffold in the cartilage tissue engineering.
The importance of a suitable place for specific growth factor cannot be overemphasized. AMM (BMP-2 in the atelocollagen, insulin and T3 in the medium) might be the ideal model for 3D culture of in vitro tissue engineering of cartilage. T3 could not be placed into the scaffold atelocollagen; otherwise, the cell would die with minimal or none GAG production. It might be possible to place T3 into the scaffold by diluting the concentration. If it is really difficult and impossible, finding out the timing for administration of hormone in the diluted concentration into the medium might be the only solution to keep the quality of tissue-engineered cartilage without hypertrophy.
The success of tissue engineering depends on interactions of the cells with the scaffolds, which not only support cell retention and functions but can also promote the cell activity through the biological stimulation by signaling molecules. The atelocollagen is regarded as one of the ideal scaffolds, because it not only facilitates the manipulation of the chondrocyte seeding, but also retains the growth factor as to enhance the cell viability in the tissue-engineered constructs and to promote the cartilage regeneration, as well. In conclusion, as our protocol of mixing BMP-2 or insulin into the atelocollagen, we proposed the possibility of enhancement of cartilage regeneration in vivo without the application of 3D in vitro culture.
In the future study, nanotechnology should be applied in the cartilage tissue engineering to decide where to place the growth factors in the scaffold atelocollagen. Controlled release and drug delivery system of the growth factor in the scaffold might be the next field we are going to focus. Great challenges are expected though multiple disciplines are required in our next project. Since tissue engineering of the cartilage is still in its infancy, our protocol of BMP-2 embedded within pellets with insulin supplement just shed a light on the strategy of making tissue-engineered cartilage with a larger size.
Conclusion
BMP-2 in the atelocollagen with the supplement of insulin in the medium could not only produce the higher GAG matrix in a shorter period but also sustain the cell viability with lower mortality. We are going to investigate the gene expression of the associated pathways so as to elucidate the detailed story in the tissue engineering.
Footnotes
Disclosure Statement
No competing financial interests exist.