
Abstract
We report on the in vitro use of Ca2+/Pi supplementation as a bio-instructive medium to drive human periosteum-derived cells (hPDCs) toward osteogenic differentiation on three-dimensional (3D) porous Ti6Al4V scaffolds. Through a multilevel factorial analysis, we have systematically investigated the biological effect and interactions of Ca2+ or Pi supplementation in three selected media preparations (i.e., basic growth medium, osteogenic medium [OM], and osteogenic medium without β-glycerophosphate [OM−]) and have identified specific conditions which induce proliferation and significant osteogenic differentiation of two-dimensional (2D) hPDC cultures. These findings were translated from 2D to 3D cultures conditions to instruct hPDCs to populate porous Ti6Al4V scaffolds and to differentiate into the osteoblast lineage with collagenous matrix production and subsequent matrix mineralization on the 3D structures. These osteogenic hybrids may potentially serve as a clinically relevant customizable bone reparative unit, providing a biomimetic template to more effectively mediate in vivo bone regeneration.
Introduction
From a clinical perspective, the customization of implant shape is required as skeletal defects are often irregular. This is now achievable due to the recent advances in the field of image-to-computer aided design and additive manufacturing technology. 5 In this context, the titanium-based biomaterial appears to be an attractive candidate, due to its clinical track record as an orthopedic biomaterial that offers excellent biocompatibility and mechanical strength. 6 From a biological point of view, the use of a well-defined cell type that is capable of generating a critical cell mass followed by the assembly of an osteogenic extracellular matrix, is another key factor to produce functional bone reparative unit. 7 This may be realized through the development of a bio-instructive culture medium that could optimally direct a specific cell type through the essential developmental stages of osteogenesis.8,9 Herein, the human periosteum-derived cell (hPDC) is a potential candidate, as it has been shown to be a clinically relevant multipotent mesenchymal stem cell (MSC) that is useful for the repair of skeletal defects in vivo with10–12 or without13,14 the use of scaffold technology.
Until now, the osteogenic medium (OM) reported by Jaiswal et al. 15 remains the gold standard for in vitro osteogenic differentiation of MSCs. 16 Different chemical compounds17–19 and growth factors20,21 have been studied and compared with this standard OM for improved osteogenic differentiation. 22 This includes calcium phosphate (CaP), a potential osteoinductive candidate, 23 which has been shown to have synergistic effects with dexamethasone (Dex, a standard osteogenic differentiation agent) on MSCs for ectopic bone formation.24,25 Additionally, CaP has been shown to promote hydroxyapatite formation that in turn stimulates osteogenic progenitor differentiation and subsequent in vivo bone formation. 26 Previously, we have reported that Ca2+/Pi supplementation into a basic growth medium (GM) enhanced proliferation and osteogenic gene expression of hPDCs in a dose- and time-dependent manner. 27 Our findings are corroborated by recent studies.28–30 However, protein markers that are functionally important for osteogenesis such as alkaline phosphatase (ALP) and collagen type-1 (Col-1) were not upregulated.
Hence, we herein study the possibility of using present knowledge to improve cell culture conditions for higher osteogenicity. For this, a multilevel factorial analysis was adopted as an experimental strategy to analyze six important variables, namely Ca2+, Pi, culture medium composition (GM, osteogenic medium with [OM] and without β-glycerophosphate [OM−]), culture time and their main effects and interaction on proliferation, ALP activity, mineralization, and osteogenic gene expression of hPDCs in vitro. The optimal Ca2+/Pi supplemented culture medium was then used as a bio-instructive medium to drive hPDC cultures on Ti6Al4V scaffolds through an enhanced proliferation stage followed by an osteogenic differentiation stage. Subsequently three-dimensional (3D) collagenous matrix deposition and mineralization was investigated to move closer to a clinically relevant biomimetic osteogenic hybrid that may be used as such or as a biomimetic osteogenic carrier for bone regeneration.
Materials and Methods
Preparation of Ca2+ or Pi supplemented culture media
As previously described, 27 4 and 8 mM of Ca2+ or Pi supplemented media were freshly prepared by diluting the 100×Ca2+ or Pi stock solution (in HEPES buffer) to 1% (v/v) in three types of culture media respectively: (i) GM (DMEM-GlutaMax™-1 [Gibco] + 10% fetal bovine serum (FBS; Gibco) + 1% antibiotics/antimycotics + 1% sodium pyruvate), (ii) OM (GM + 100 nM Dex + 50 μg/mL ascorbic acid + 10 mM β-glycerophosphate), and (iii) OM− (Table 1). These ion concentrations represent the high and low values within the effective concentration window obtained in the previous study. 27 Based on the data sheets provided by the manufacturers of the culture medium, FBS and chemical compounds, it was calculated that all culture media used in this study contained a basal level of 1.838 mM Ca2+ and 0.9419, 1.149, and 11.149 mM of Pi in GM, OM−, and OM, respectively. These values were validated using inductively coupled plasma-atomic emission spectroscopy (data not shown).
Ca2+ is denoted as “C” and Pi as “P,” follows by a number indicating only the concentration of the exogenously added ions in millimole (mM).
GM, growth medium; OM, osteogenic medium; OM−, osteogenic medium without β-glycerophosphate.
Optimization of Ca2+/Pi supplementation for improved cell proliferation and osteogenic differentiation using two-dimensional hPDCs cultures
Culturing of hPDCs
hPDCs were isolated from periosteal biop, 2.2.sies of different donors (n=6, age 14.9 [S.D.±2.1]) as previously described. 31 This procedure was approved by the ethical committee for Human Medical Research of the Katholieke Universiteit Leuven and was done with signed patient informed consent. Pooled passage 3 hPDCs from these donors were expanded in GM and harvested upon confluence before plated onto cell culture dishes at a density of 4500 cells/cm2. After 2 days of culture in GM, the cells were synchronized in serum free medium containing 0.01% bovine serum albumin for 24 h, treatments were started by adding freshly prepared Ca2+ or Pi supplemented media into the cell cultures. The samples were harvested at defined time points for analysis as described below.
Proliferation and ALP activity analysis
The proliferation and ALP activity of hPDCs (in 24-well plate) were assessed at 7, 14, 21, and 28 days of treatment (n=4). Briefly, at each time point, cells were harvested and lysed in 0.05% Triton-X100 (in phosphate-buffered saline [PBS]), frozen and thawed three times, and centrifuged at 13,000 rpm for 1 min to obtain cleared cell lysates. The DNA content was quantified using a Quant-iT™ dsDNA HS assay kit (Invitrogen) and the ALP levels were measured using a BluePhos® microwell phosphatase substrate system (Kirkegaard & Perry Laboratories). The ALP activity was calculated by normalizing the absorbance to the DNA content of the same sample.
Quantitative reverse-transcription polymerase chain reaction
At 21 days of treatment, total RNA of each sample (in 12-well plate, n=3) was extracted using a RNA extraction kit (Qiagen) and quantified using a Nanodrop ND-1000 spectrophotometer (Thermo Scientific). Complementary DNA (cDNA) was synthesized by reverse transcription of 250 ng of total RNA using a RevertAid™ H Minus First Strand cDNA synthesis kit with Oligo(dT)18 as primer (Fermentas). Sybr green qPCR primers (Col-1F 5′-GACGAAGACATCCCACCAAT-3′ Col-1R 3′-AGATCACGTCATCGCACAAC-5′; OCNF 5′-GTGCAGCCTTTGTGTCCAA-3′ OCNR 3′-GCTCACACACCTCCCTCCT-5′; Runx2F 5′-CGCATTCCTCATCCCAGTAT-3′ Runx2R 3′-AGTCTAATGTCTGGGGTCCG-5′; BMP-2F 5′-GTATCGCAGGCACTCAGGTC-3′ BMP-2R 3′-TGGTCTTTGCTCACCCTTTT-5′) were designed to span an intron so only RNA specific amplification was possible. The reverse-transcription polymerase chain reaction (PCR) reaction was cycled in a Rotor-Gene sequence detector as follows: 95°C for 3 min, 40 cycles of 95°C for 3 s, and 60°C for 60 s. Each sample was tested in duplicate and compared with β-actin expression as an external control to allow normalization of results. Relative differences in expression were calculated using the 2−ΔCT method 32 and statistical significance calculated by comparing with the 2−ΔCT of the samples treated with nonion supplemented GM, OM−, or OM.
Histological examination
The cell aggregates that only formed in cultures treated with OM−C4 and OMC4 (i.e., OM− or OM supplemented with 4 mM of Ca2+) after 14 and 21 days were collected and fixed with 4% paraformaldehyde for 1 h before embedding in paraffin for histological sectioning and analysis. Masson's trichrome staining was performed to demonstrate the production of collagen matrix. The formation of mineralized nodules within the cell aggregate was visualized by Von Kossa staining.
Mineralization assay
For each treatment condition, mineralization of the cell cultures was assessed by Alizarin Red staining (pH 4.2) after 28 days of culture. Briefly, cell cultures were rinsed with PBS followed by fixation in 4% formalin (in PBS) for 10 min and washed with distilled water before staining with Alizarin Red solution for 1 min. Nonspecific staining was removed by rinsing in distilled water. The deposited minerals were quantified by dissolving the dye in 10% cetylpyridinium chloride solution with constant agitation (n=3). Subsequently, 100 μL of the solution from each sample was transferred to a 96-well plate and the absorbance read at 570 nm with a microplate reader (TECAN). 33
In vitro biomimetic engineering of osteogenic hybrids on 3D porous Ti6Al4V scaffolds
Production of additive manufactured 3D porous Ti6Al4V scaffolds
3D cylindrical porous Ti6Al4V scaffolds, (ø 6×3 mm height) with a centralized longitudinal channel (ø 2 mm, to facilitate nutrient-waste exchange), were designed using Magics (Materialise N.V.) software and fabricated by selective laser melting. 5 The physical properties including strut thickness, surface area, pore size, and porosity of the produced Ti6Al4V scaffolds were characterized by micro-computed tomography (μ-CT) (SkyScan N.V.) (n=3). The produced scaffolds were cleaned as previously described 34 and prewetted with GM for 2 h before cell seeding.
Dynamic rotation seeding and culture of hPDCs on 3D porous Ti6Al4V scaffolds
Approximately 200,000 hPDCs (in 35 μL of GM) were drop seeded onto each of the prewetted Ti6Al4V scaffolds in 1.5 mL eppendorf tubes and subsequently statically incubated for 1 h in a humidified incubator to allow cell attachment. Then, an additional 1 mL of GM was added into each sample before they were transferred onto a Mini LabRoller Rotator (Labnet Int. Inc.) to perform dynamic rotation seeding overnight. The cell-seeded scaffolds were then transferred into 24-well plates and placed on a 3D platform rotator (30 rpm; Grant-bio) and treated with freshly prepared Ca2+ or Pi supplemented culture media at each medium refreshment (twice a week) and subsequently incubated for the indicated time-points before analysis. Cell seeding efficiency (CSE) was calculated by measuring the DNA content of the cell seeded-scaffolds and normalized to the DNA content of the cell suspension used for drop seeding (n=3). 34
Monitoring of hPDC growth and viability on 3D porous Ti6Al4V scaffolds
hPDC growth on 3D porous Ti6Al4V scaffolds was monitored by nondestructive analysis of the metabolic activity of each sample at defined time points using AlamarBlue® (Invitrogen) (10% in GM), according to the manufacturer's instructions (n=4). After 28 days of treatment, the total DNA content of all samples was quantified and the corresponding cell numbers were calculated based on the measured DNA content per cell (n=3). Cell viability was characterized by staining the living and dead cells using the Live/Dead® cell viability kit (Invitrogen).
Osteogenic differentiation of the engineered 3D osteogenic hybrid
The representative early and late markers for osteogenic differentiation, including Runx-2, Osterix and ALP, were quantified at 21 days of treatment (n=3). Briefly, the samples were harvested in TRIzol® reagent (Invitrogen), and the total RNA was phase-separated by chloroform before purification using a RNA extraction kit (Qiagen). 1 μg of purified RNA was used to synthesize cDNA and used to measure the osteogenic differentiation related gene expression by Sybr green qPCR as described above: Runx2, Osterix (OsxF 5′-AGTGACCTTTCAGCCTCCAA-3′ OsxR 3′-GGGAAAAGGGAGGGTAATCA-5′) and ALP (ALPF 5′-GGACATGCAGTACGAGCTGA-3′ ALPR 3′-GTCAATTCTGCCTCCTTCCA-5′). Additionally, the ALP activity of each sample was quantified as described above to complement the osteogenic differentiation event (n=3).
Characterization of 3D osteogenic extracellular matrix production
The 3D collagen matrix production and matrix mineralization on porous Ti6Al4V scaffolds was characterized at 21 days of treatment by Picro-Sirius Red staining (1 mg/mL Sirius Red in saturated Picric acid, n=3) 35 and Alizarin Red staining (n=3, as described above). The stained samples were thoroughly washed with distilled water to remove unbound dye and dried at 37°C before qualitative analysis by stereomicroscopy. For quantitative analysis, the picro-sirius red dye was dissolved in 0.2 M NaOH/Methanol (1:1 ratio) with mild shaking overnight, and the optical density was measured at 492 nm using a microplate reader (TECAN). The morphology and CaP crystal structure of the mineralized matrix was characterized by scanning electron microscopy (SEM) coupled with energy dispersive X-ray (EDAX) analysis (FEI XL30 FEG) at 10 kV.
Multilevel factorial and statistical analysis
The in vitro experiments were defined so that experimental data obtained from proliferation (n=4) and gene expression assays (n=3) fitted a design of experiment analysis to assess the main effects and interactions of the following four parameters at different levels: (i) Ca2+ (three levels: 0, 4, and 8 mM), (ii) Pi (three levels: 0, 4, and 8 mM), (iii) medium type (three levels: GM, OM−, and OM), and (iv) culture duration (four levels: 7, 14, 21, and 28 days), on both cell proliferation and gene expression using MINITAB 14® (Minitab Inc.) software. All data were expressed as mean±standard deviation or mean±standard error of the mean. One-way analysis of variance (ANOVA) was performed to evaluate the statistical difference of the means of main effects and interactions of multiple parameters on cell proliferation and osteogenic gene expression. An unpaired Student's t-test (two-tailed) was performed to compare means of two parameters, both by establishing the statistical significance at p<0.05.
Results
Morphological change and osteogenic matrix production of hPDC two-dimensional cultures
Figure 1A shows microscopic images that represent the morphological changes observed in two-dimensional (2D) hPDC cultures after 28 days of treatment with various Ca2+/Pi supplemented media. The morphology of cells cultured in normal culture condition (i.e., GM) appeared fibroblastic (↑), but changed to cuboidal (Δ) when cells were treated with GMC4 and GMC8. No phenotypic change was induced by GMP4 and GMP8 treatments, instead mineral deposits (*) were observed. In contrast, mineral deposition was not observed in OM− and Pi supplemented-OM− conditions, but had occurred in Ca2+ supplemented-OM− treated cultures. Interestingly, OM−C4 treatment resulted in cell aggregation (g) after 14 days of treatment. Similarly, the OM and OMC8 caused mineral deposition, and cell aggregates were formed in OMC4 treated cultures at 14 days. These observations were supported by both the Alizarin Red staining (Fig. 1B) and the quantification of calcium mineral content (Fig. 1D) that showed significantly higher levels of matrix mineralization in the corresponding treatment conditions. Besides the specific red staining of the mineralized matrix (m) by Alizarin Red staining, dark mineral deposits (revealed as dark deposits that is labeled as “*” in Fig. 1B) were observed on the cell monolayer treated with the specific Ca2+/Pi supplemented media which was not found in OM treated conditions.
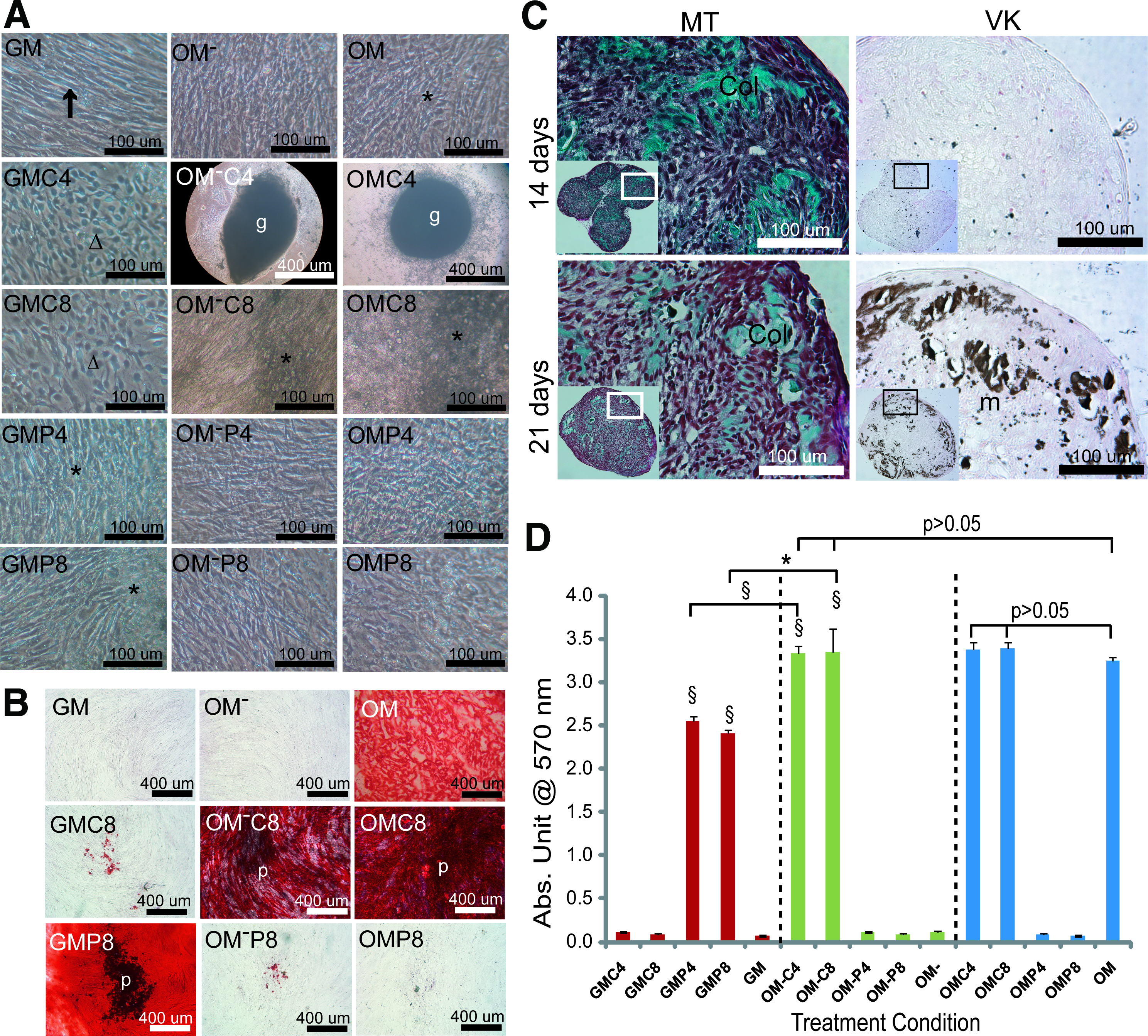
Morphological change and mineralization of two-dimensional (2D) hPDC cultures after 28 days of treatment.
By histological analysis of the cell aggregates formed in OM−C4 and OMC4 conditions, abundant collagen matrix (Col, stained green by Masson's trichrome staining) was observed at 14 days of treatment with no evidence of mineralization (Fig. 1C). This collagen matrix gradually diminished over time and was replaced by an accumulation of mineralized nodules (m, stained brownish by von-Kossa staining) at the periphery of the aggregates at 21 days. Similar dynamics of bone-related matrix production was observed in cell aggregates obtained from the OMC4 condition (data not shown).
Proliferation of 2D hPDC cultures and the multilevel factorial analysis
Based on the DNA content measurements, Ca2+/Pi supplemented-GM induced significantly higher proliferation of hPDCs (1.9–2.6-fold higher) than the nonsupplemented GM at each time point (Fig. 2Ai). This mitogenic effect was more prominent for GMC4 (at 21 [p<0.05] at 28 days [p<0.01]) and GMP4 (at 7 [p<0.05] at 14 days [p<0.001]) when compared with GMC8 and GMP8, respectively. There were no significant differences when similar [Ca2+] and [Pi] were used to treat the cells for the same duration (p>0.05 for all time points). Interestingly, OM− treatment induced a superior pro-mitogenic effect when compared with GM, indeed OM− treatment for 7 days resulted in a 1.5-fold higher DNA content than GM treatment for 28 days (p<0.01) (Fig. 2Aii). This pro-mitogenic effect of OM− was further enhanced by Ca2+/Pi supplementation that resulted in ∼1.6–1.8-fold higher DNA content than the supplemented-GM counterparts at 7 days. Encouragingly, OM−P4 consistently resulted in the highest DNA content at 7 (p<0.01), 14 (p<0.05), and 21 days (p<0.01) before declining slightly at 28 days (probably due to cell–cell contact inhibition effect [Ref. 36 ]). At this time point, the OM−C8 treatment produced the highest DNA content (about 1.5-fold higher than OM−P4 treatment [p<0.001]). Although cell aggregation could be observed in OM−C4 treated cultures at 14 days, the DNA content increased till 21 days before declining at 28 days, but not statistically lower than OM−.
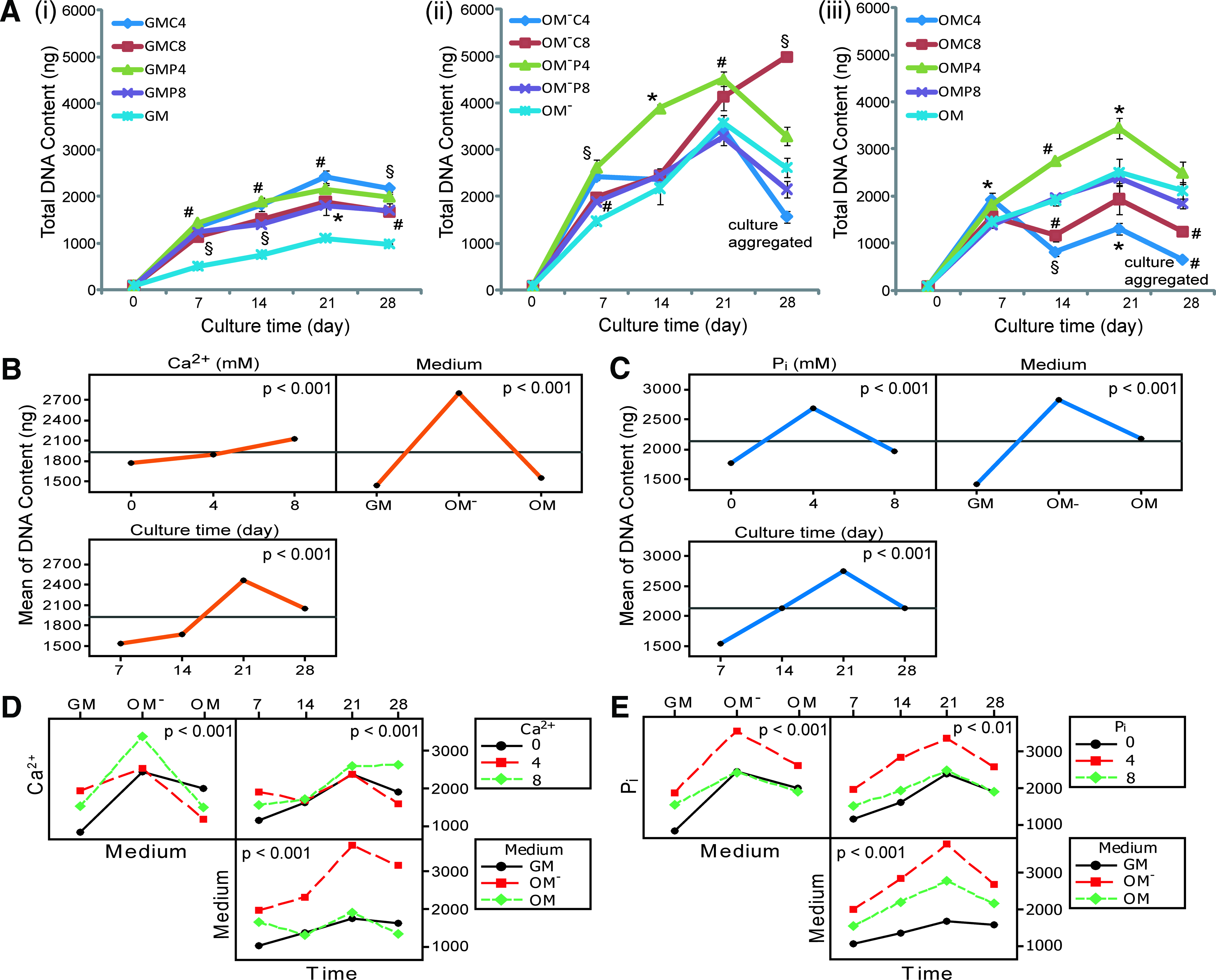
Proliferation of hPDC 2D cultures over 28 days of treatments and the multilevel factorial analysis.
The mitogenic effect of OM was not as prominent as that of OM−, however the DNA content in OM cultures was still 2.2–2.9-fold greater than GM over 28 days (Fig. 2Aiii). Interestingly, OM supplemented with 4 mM of Pi (i.e., OMP4) resulted in the highest DNA content at 14, 21, and 28 days (ANOVA, p<0.001). Both OMP8 and OM showed a similar effect on cell proliferation over 28 days, whereas OMC8 treatment produced a significantly lower DNA content than OM at 14 and 28 days (p<0.01 for both cases). Surprisingly, cell aggregation in OMC4 treated cultures had an adverse effect on cell proliferation that resulted in the lowest DNA content at 14 (p<0.001), 21 (p<0.05), and 28 days (p<0.01).
Based on the multilevel factorial analysis, the Ca2+, Pi, medium type and culture duration had a significant effect on hPDC proliferation (p<0.001) and their interactions were statistically significant (p<0.01) except for the interaction between Pi, medium and culture time (Pi*Medium*Time, p>0.05) (Supplementary Table S1 showing the general linear model and ANOVA analysis; Supplementary Data are available online at
Induction of ALP activity in 2D hPDC cultures
As previously reported, no induction of ALP activity was seen in hPDC cultures treated with GM or Ca2+/Pi supplemented-GM over the 28 days. 25 However, both OM and OM− treatment induced significant ALP activity from 3.56 abs. unit/mg DNA (at 7 days) to value of 64-fold higher at 28 days (229.16 abs. unit/mg DNA) (Fig. 3A). Interestingly, supplementation of Ca2+ into OM−, specifically OM−C8, induced a higher ALP activity than OM− at 14 days that became significantly higher at 21 days (p<0.05) before declining at 28 days (p<0.01) (Fig. 3Ai). OM−C4 treatment resulted in a significantly lower ALP activity than OM− over 21 days before rising to a level comparable to that of OM−C8 treatment at 28 days (a normalizing effect caused by the relatively low DNA content at 28 days due to cell aggregation in OM−C4 condition). Supplementation of Pi into OM− was not beneficial, as OM−P8 treatment caused a significantly lower ALP activity than OM− at 14 and 21 days (p<0.05 for both cases) (Fig. 3Aii). Conversely, supplementation of Ca2+ or Pi into OM had a negative effect on ALP activity that was significantly lower than OM treatment at 21 days by OMP8 (p<0.01) (Fig. 3Aiv) and at 28 days by OMC4 (also due to low DNA content caused by cell aggregation, p<0.01) (Fig. 3Aiii).
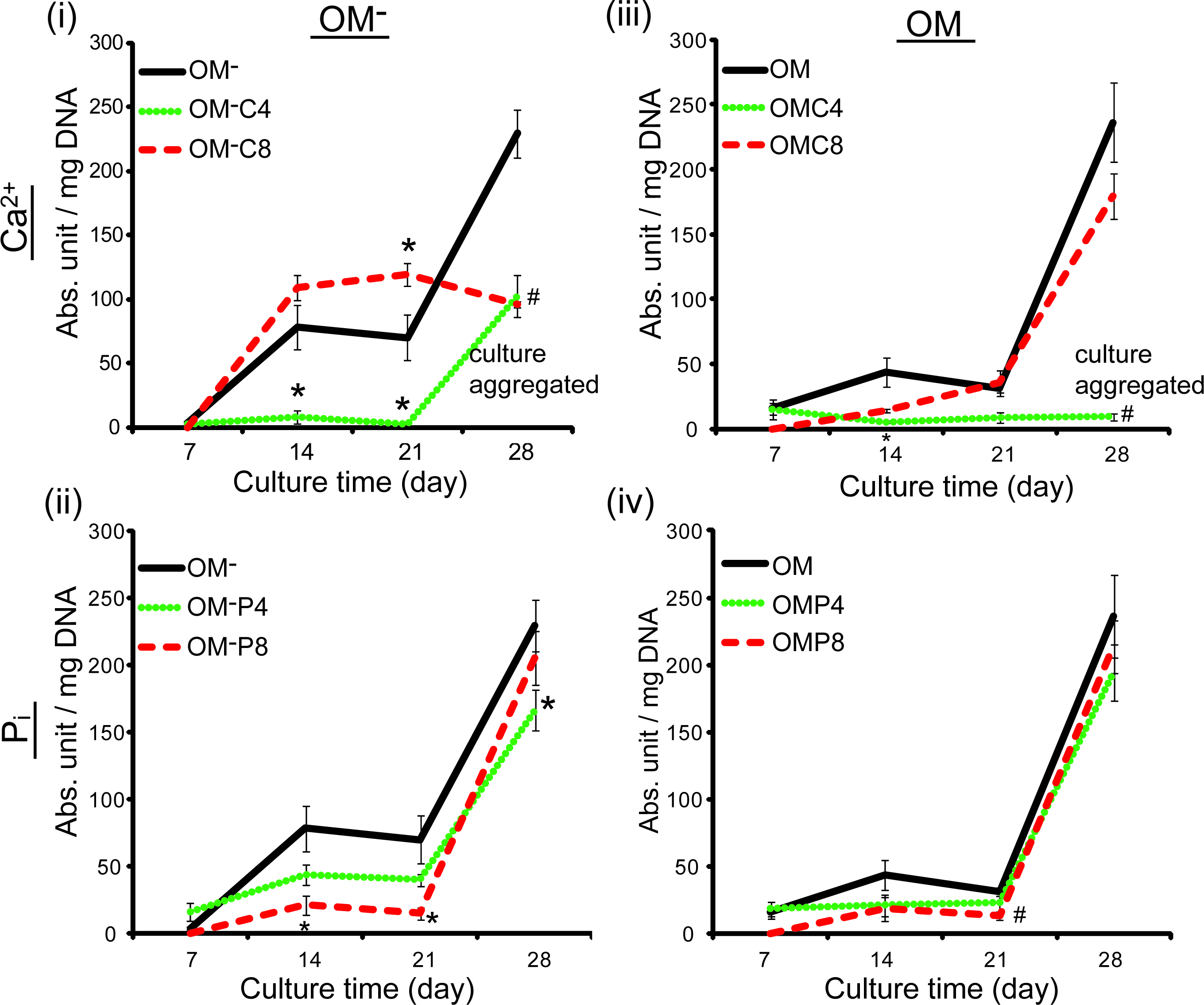
Dynamics of alkaline phosphatase (ALP) activity of 2D hPDC cultures over 28 days of treatment with Ca2+/Pi supplemented-OM−
Multilevel factorial analysis of osteogenic gene expression in 2D hPDC cultures
The main effects and interactions of Ca2+, Pi and medium type on Col-1, OCN, Runx2, and BMP-2 gene expression of 2D hPDC cultures after 21 days of treatment were analyzed by a multilevel factorial analysis. The results of which are presented in Figure 4 and Supplementary Table S2, the latter showing the general linear model and the ANOVA analysis.
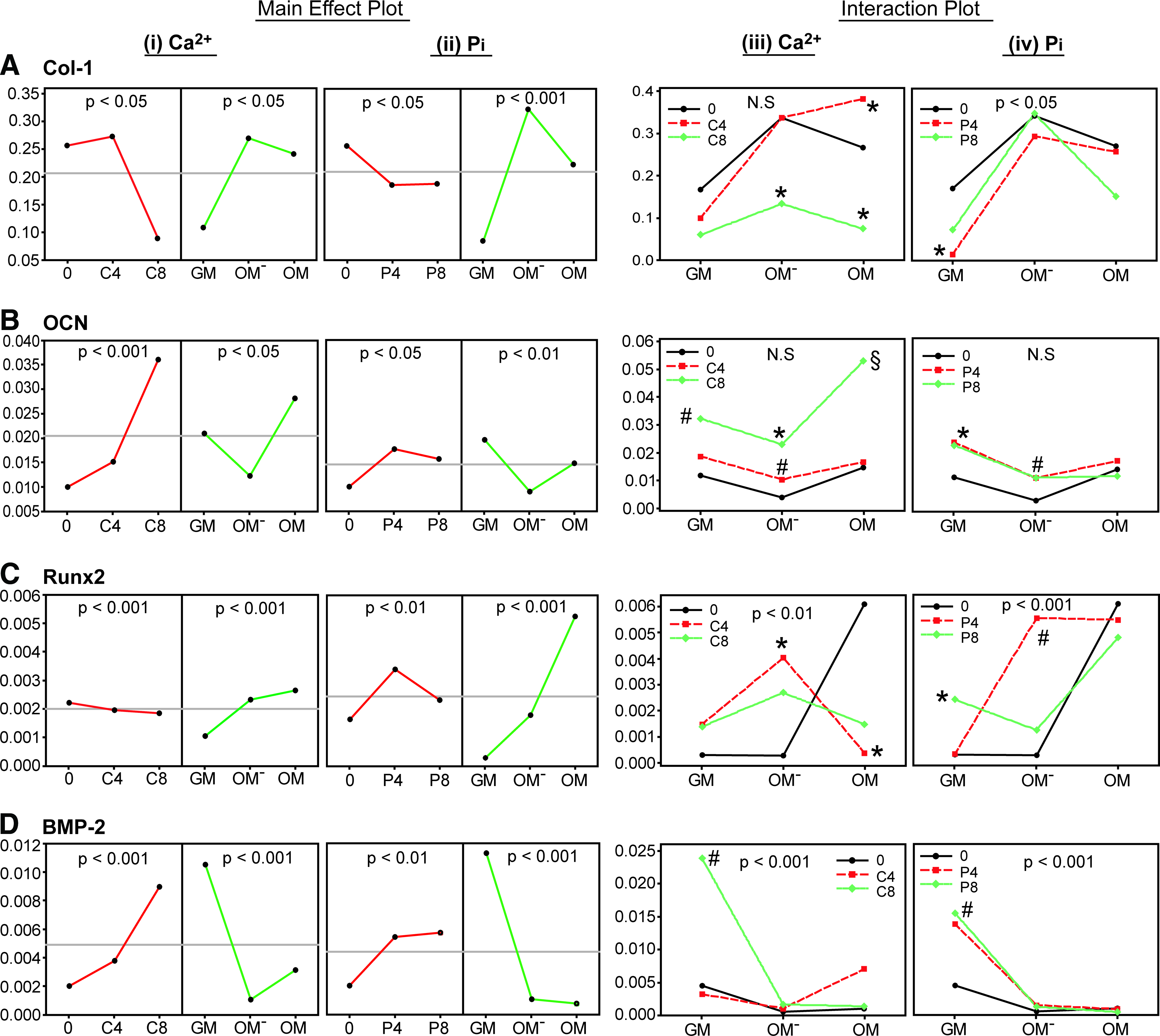
Multilevel factorial analysis of the main effects
Based on the main effect plots, an increase in [Ca2+] and [Pi] had a significant adverse effect on Col-1 expression (p<0.05 for both cases), meanwhile OM− or OM supplemented with Ca2+ (p<0.05) or Pi (p<0.001) significantly promoted Col-1 expression (Fig. 4Ai, ii). These results indicated that the adverse effect of Ca2+ and Pi on Col-1 expression could be compensated by supplementing appropriate concentrations of these ions into OM− or OM. This is shown by the interaction plots, where OMC4 and OM−P8 induced a comparable Col-1 expression level to that of OM and OM−, respectively (Fig. 4Aiii, iv). The expression of OCN was significantly enhanced by the addition of Ca2+ (p<0.001) or Pi (p<0.05), and the OM was the optimal medium for Ca2+ (p<0.05), and GM for Pi (p<0.01) supplementation (Fig. 4Bi, ii). This was supported by the interaction plots, where OMC8 (p<0.001) and GMP4 (p<0.05) significantly upregulated OCN expression (Fig. 4Biii, iv).
Downregulation of Runx2 expression by Ca2+ was observed, however the effect was not significant, and supplementation of Pi significantly upregulated the gene expression (p<0.01), meanwhile OM was more potent than OM− and GM to enhance Runx2 expression (Fig. 4Ci, ii). However, supplementing Ca2+ into OM significantly downregulated the gene expression (p<0.05), and OM− appeared to be the medium of choice for Ca2+ supplementation, where OM−C4 significantly enhanced Runx2 expression (p<0.05) (Fig. 4Ciii). Interestingly, supplementing 4 mM of Pi into OM−, induced significantly higher Runx2 expression than the nonsupplemented-OM−, which was comparable to the effect by OM treatment (Fig. 4Civ). Finally, BMP-2 gene expression was significantly enhanced by Ca2+ (p<0.001) or Pi (p<0.01) in GM in a dose-dependent manner. However, this effect was less pronounced when these ions were supplemented into OM− or OM (p<0.001 for both media) (Fig. 4Di, ii). These results suggest that GM should be used with Ca2+ or Pi supplementation to significantly upregulate BMP-2 expression, as shown by the significant upregulation of BMP-2 expression by GMC8 and GMP8 treatments (p<0.01 for both cases) (Fig. 4Diii, iv).
Scaffold characteristics and hPDC growth and viability on 3D porous Ti6Al4V scaffolds driven by Ca2+/Pi supplemented-OM−
Based on the μ-CT analysis, the produced 3D porous Ti6Al4V scaffolds had a mean strut thickness of 256.11±7.39 μm, providing a total strut surface area of 2.396±0.09 cm2 per scaffold (Fig. 5A). The scaffolds were fully open structures with a porosity of 80.56%±0.53%, a mean pore size of 821.82±6.35 μm and a total pore volume of 64.82±1.96 mm3 per scaffold (excluding the void volume of the 2 mm centralized longitudinal channel).
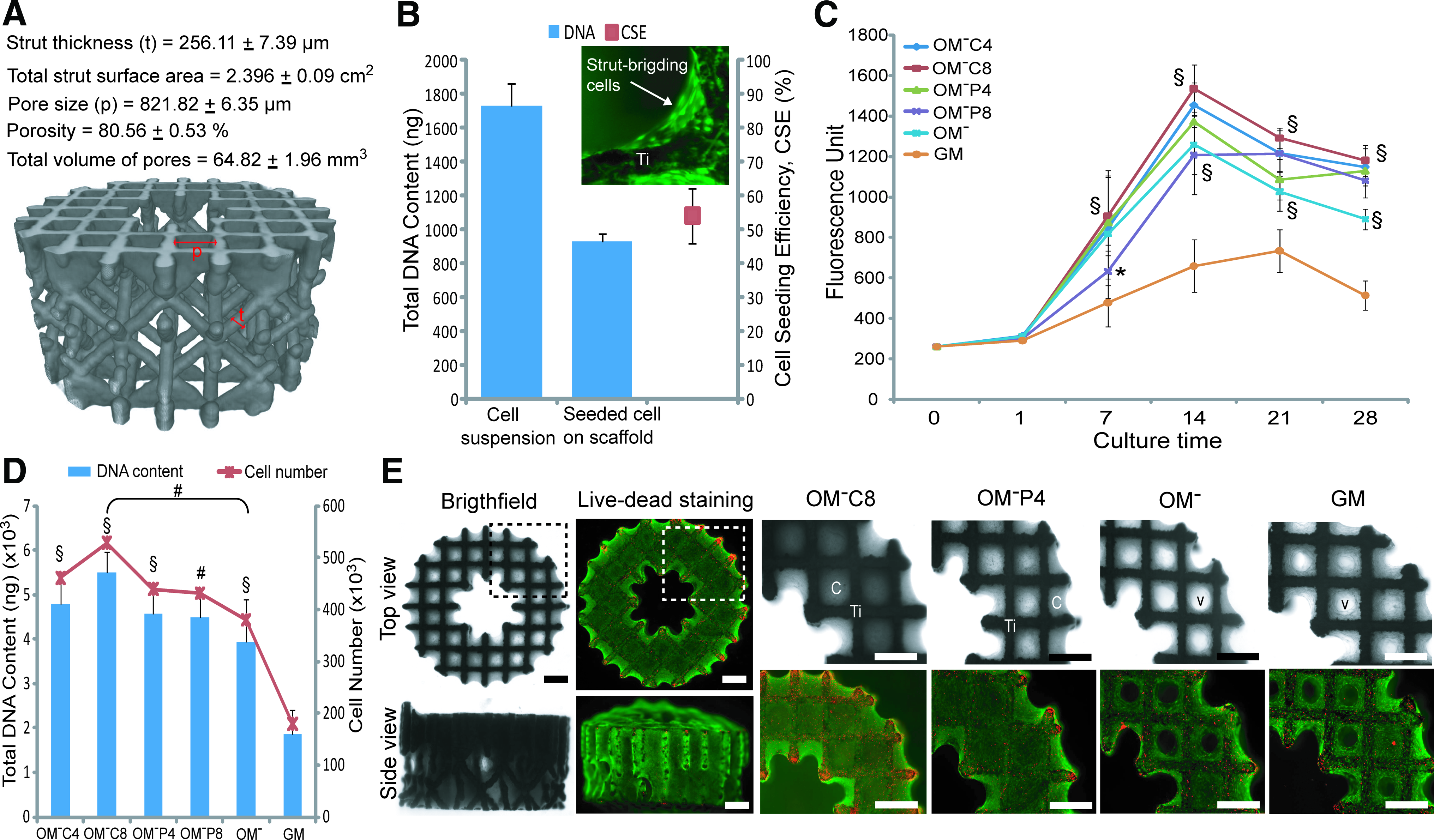
In vitro engineered three-dimensional (3D) osteogenic hybrid in porous Ti6Al4V scaffold driven by OM− supplemented with Ca2+ or Pi.
A CSE of 53.9%±8.11% was achieved when seeding 200,000 hPDCs onto these scaffolds (Fig. 5B), thus resulting in a seeded cell density of ∼45,000 cells/cm2 (based on the measured value of 10.39 pg DNA per cell in the cell suspension used for drop seeding) which was 10×higher than the cell density used for 2D hPDC cultures in this study. Live-dead staining of the seeded scaffold showed that the seeded living cells were bridging the strut corners (inset of Fig. 5B), thus suggesting that the currently used seeding protocol accommodates initial cell numbers onto the scaffold at a higher cell to surface ratio than 2D cultures.
Subsequent culturing of these cell seeded-scaffolds in OM− induced a significantly higher cell growth (1.4–1.9-fold higher) than in GM over the 28 days that was further enhanced by Ca2+ or Pi supplemented-OM− treatments, as indicated by the increase of the metabolic activity over 28 days (Fig. 5C). Among these, OM−C8 induced the highest metabolic activity at all time points (p<0.001), which was 1.8–2.3-fold higher than GM. This was supported by quantification of DNA content from the 28 day samples, where cell-seeded scaffolds treated with OM−C8 produced the highest DNA content (∼529,000 cells per scaffold). This was 1.4-fold higher than OM− (p<0.01, ∼381,000 cells per scaffold) and 2.9-fold higher than GM (p<0.001, ∼180,000 cells per scaffold) treatments (Fig. 5D). Brightfield microscopic images showed that the scaffolds were highly populated by cells growing in three dimensions, with almost complete closing of the pores by hPDCs under Ca2+ or Pi-supplemented treatments. This phenomenon was not observed at the same point in time for OM− or GM treated samples (Fig. 5E). These highly viable, cell filled constructs are termed “engineered 3D osteogenic hybrids.”
Osteogenic differentiation of the engineered 3D osteogenic hybrids
Quantitative PCR analysis showed that the engineered 3D osteogenic hybrids under Ca2+/Pi supplemented-OM− treatments for 21 days exhibited significantly higher expression of Runx2 than in GM condition (p<0.01 for OM−C4, and p<0.05 for OM−C8, OM−P4, and OM-P8) which was comparable to the effect of OM− treatment (p>0.05) (Fig. 6A). Additionally, Osterix (Osx)–a transcription factor essential for osteoblast differentiation and bone formation 37 was found to be significantly upregulated by the Pi supplemented-OM- treatments (but not Ca2+) that resulted in 3.97-fold (by OM-P4, p<0.05) and 4.40-fold (by OM-P8, p<0.001) higher expression than OM− or GM treatments (Fig. 6B). Interestingly, Pi supplemented-OM− (but not Ca2+) induced significantly higher ALP gene expression than GM treatment (p<0.05), which was comparable to OM− treatment (p>0.05) (Fig. 6C). Similar ALP protein activities were observed in different treatment conditions (Fig. 6D).
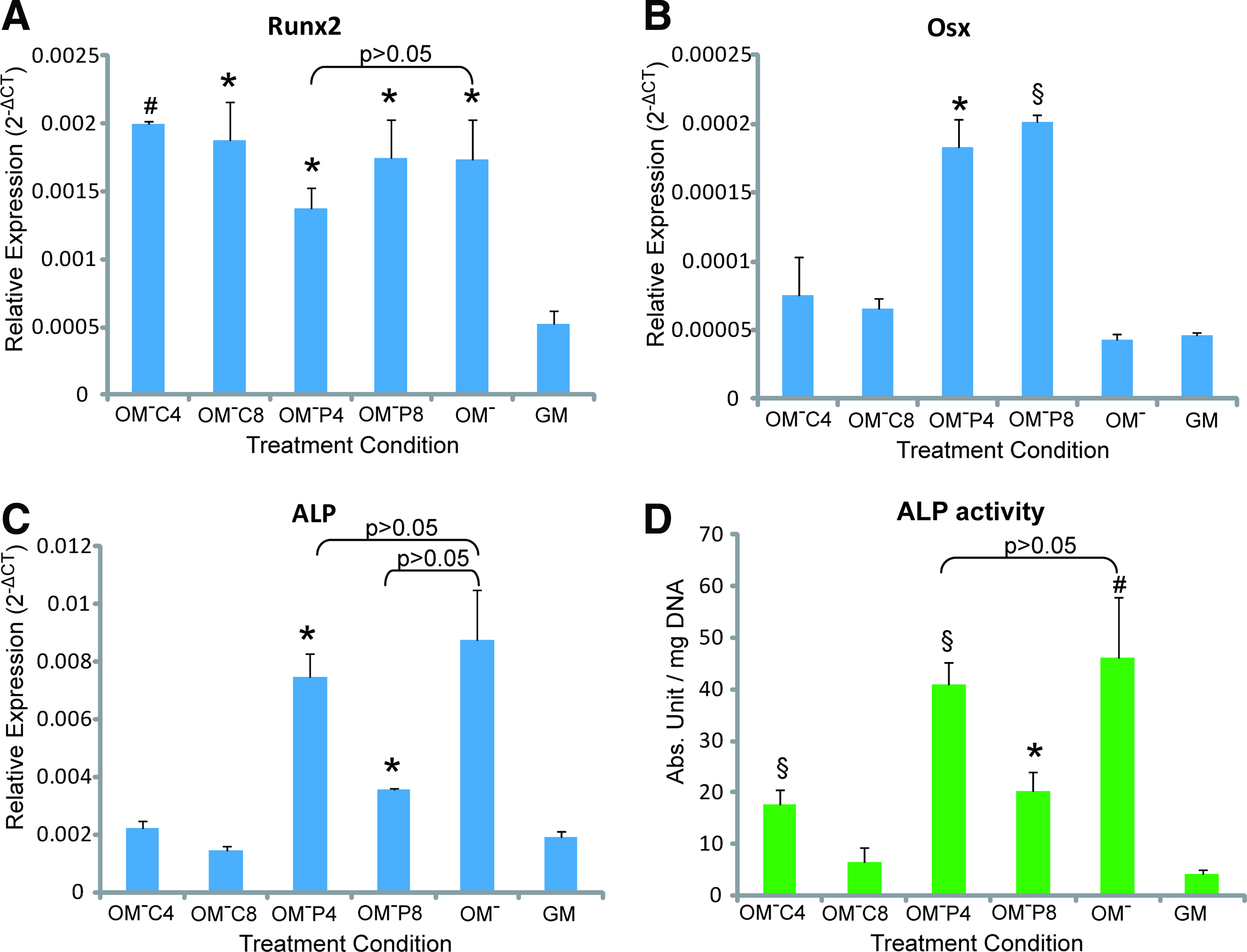
Osteogenic differentiation of the engineered 3D osteogenic hybrid as compared with the GM condition, by quantitative polymerase chain reaction analysis on
3D collagenous matrix production and matrix mineralization on the engineered 3D osteogenic hybrids
Figure 7A shows representative brightfield images of the noncollagenous constructs and collagenous constructs formed under different treatment conditions. This collagenous matrix (Col) was found to be deposited throughout the porous Ti6Al4V scaffolds in a 3D manner (stained in orange) after treatment with Ca2+ or Pi supplemented-OM− and nonsupplemented-OM−, but not for GM. Quantitatively, OM−C4 treatment induced the highest 3D collagenous matrix production that was significantly higher than OM− (p<0.05) and GM (p<0.01) treatments (Fig. 7B). Mineralized matrix deposition (↑) was observed throughout the porous Ti6Al4V scaffold (stained in red) after treatment with Ca2+ or Pi-supplemented-OM−, which was not observed in GM treated samples (Fig. 7C). Quantitatively, Ca2+ and Pi-supplemented-OM− resulted in ∼3-fold higher mineralized matrix formation than OM− and GM (Fig. 7D). Interestingly, this mineralized matrix was found mostly around the corner of the strut nodes but not in the centre of the pores. By SEM, this mineralized matrix had a distinctive microstructure and a specific CaP crystal structure. Ca2+ supplemented-OM− treatment resulted in nanometer rod-shape crystal structures (∼50 nm in width) and Pi-supplemented-OM− treatment produced submicrometer spherical CaP crystal particles (∼100–250 nm in diameter) (Fig. 7E). These structures were confirmed to be CaP as shown by EDAX analysis (insets of Fig. 7E).
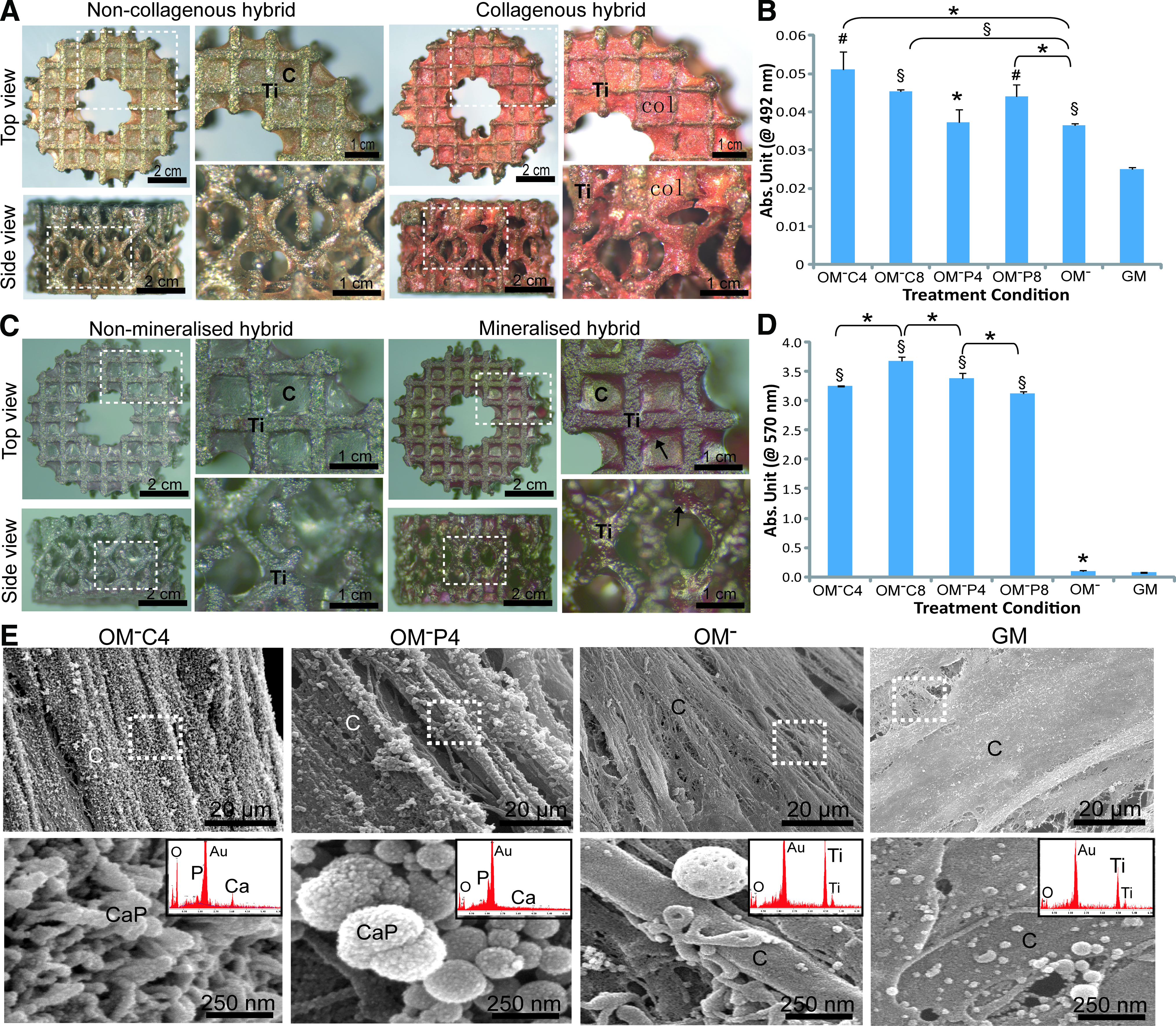
Osteogenic extracellular matrix production of the engineered osteogenic hybrids after 21 days of treatment in Ca2+/Pi supplemented-OM− as compared with GM condition. Representative images of
Discussion
In this study, we systematically investigated three types of culture media, supplemented with Ca2+ or Pi, as bio-instructive medium for hPDCs. The purpose of which was to biomimetically engineer a 3D osteogenic hybrid in vitro. We have described the change cell morphology and of matrix mineralization that were dependent on the different treatment conditions. These phenomena may be associated with the osteogenic differentiation of hPDCs by Ca2+/Pi supplementation. 38 The presence of dark mineral deposits (which were not observed in the OM treated condition) in addition to the specific calcium-mineral staining by Alizarin Red, may possibly be due to the presence of a high Pi level or an interaction between Ca2+ and OM− on promoting CaP precipitation. Through the use of multilevel factorial analysis, 39 we have described the main effects and interactions of [Ca2+], [Pi], medium type and culture time on cell proliferation and osteogenic gene expression. Our results showed that OM− induced the highest proliferation of hPDCs in 2D cultures (as compared with GM and OM) that was synergistically enhanced by specific Ca2+ or Pi supplementation. This enhancement of proliferation by OM− is believed to be induced by Dex and ascorbic acid, as these compounds are known to promote cell proliferation.40,41 Meanwhile, Ca2+ and Pi induced proliferation was associated with the role of these ions in modulating cell cycle progression. 42
Additionally, we have shown that the native non-ALP expressing hPDCs 43 could be differentiated into a highly ALP positive cell population through the use of OM− or OM (an effect mainly attributed to Dex [Ref. 44 ]), which has been reported to be an essential entity for ectopic bone forming capability of human MSCs in vivo. 45 However, the presence of Ca2+ and particularly the presence of Pi (inorganic phosphate) in OM− or OM (organic phosphate) compromised the effect of Dex on ALP induction. This reduction in ALP activity may be due to the activation of the cellular regulatory feedback loop that responds to the supraphysiological levels of these ions added into the cultures. This represents an important regulatory mechanism of hPDCs in controlling biological apatite formation. 46 This data was supported by the Alizarin red staining of 2D cultures where no matrix mineralization was induced by Pi-supplemented-OM− or–OM. Moreover, Alizarin red staining revealed no matrix mineralization in 2D cultures treated with OM− (although high ALP activity was measured), whereas matrix mineralization was observed in OM and in Ca2+ supplemented-OM− or–OM treated cultures that contained β-glycerophosphate. These data suggest that the availability of an appropriate chemical form of Ca2+ and Pi is critical to induce matrix mineralization.47,48 However, the effect of β-glycerophosphate induced mineralization has been previously reported to cause the inhibition of further cell growth.49,50 This provides more insight in the fine-tuning of culture conditions to induce mineralization in hPDC cultures.
In this study, we have demonstrated that the downregulation of Col-1 gene expression by Ca2+ and Pi on 2D hPDC cultures could be compensated by supplementing these ions into OM− or OM, which both contained ascorbic acid that serves as a co-factor for collagen synthesis. 51 This finding was supported by the histological analysis of the cell aggregates that produced abundant collagen matrix. This is of crucial importance as collagen matrix production is essential to support proliferation and differentiation of MSCs 52 and being a predictive biomarker for the bone-forming potency of MSCs. 53 Subsequently, our results suggested that Ca2+/Pi supplementation into OM− or OM had significant beneficial effects on osteogenic differentiation of 2D hPDC cultures, as OCN and Runx2 expression were significantly upregulated. These phenomena can be explained by the effect of Ca2+ on the upregulation of OCN expression (as OCN is a calcium-binding protein and a negative regulator of extracellular matrix mineralization) 54 and by the fact that Pi maybe associated with the presence of a Pi sensing mechanism of osteoblastic cells that is functionally linked to a nuclear export system of Runx2. 55 Indeed, Runx2 is a key transcription factor associated with osteochondroprogenitor differentiation 56 and high expression of Runx2 by MSCs is beneficial to accelerate bone defect healing. 57 However, the upregulation of a potent osteoinductive growth factor (i.e., BMP-2) 58 induced by Ca2+/Pi supplemented-GM was abrogated when an OM− or OM system was used. This is possibly due to the dominant differentiation effect of the osteogenic supplements in OM− or OM (such as Dex) that overrules the Ca2+ or Pi mediated expression of BMP-2.
Based on the experimental data obtained from 2D hPDC cultures, we concluded that Ca2+/Pi supplemented-OM− was the most bio-instructive medium for in vitro biomimetic engineering of a 3D osteogenic hybrid. Indeed, our results showed that the enhanced proliferative effect of this medium was successfully extended to 3D cultures in porous Ti6Al4V scaffolds, where Ca2+/Pi supplemented-OM− induced significantly higher cell growth and facilitated the formation of 3D hybrid. This hybrid was comprised of highly viable cells secreting osteogenic extracellular matrix that filled the pores of 3D Ti6Al4V scaffolds. This 3D cellular morphogenesis has been shown to be essential for ex vivo formation of human bone. 59 In addition, this study showed that there was a different surface area-dependent cell growth in 3D porous scaffolds as compared with 2D culture, and also that a specific medium can direct high cell proliferation (thus a critical cell mass production) together with osteogenic differentiation. 60
Further, we have provided evidence that 3D hPDC osteogenic differentiation was driven by Ca2+/Pi supplemented-OM−, where the treated 3D hPDC cultures showed significant upregulation of Runx2 and Osterix 61 expression as compared with nonsupplemented OM− and GM. This data indicates the potency of this medium to induce functional differentiation of hPDCs into the osteogenic lineage. This osteogenic functionality was further supported by the significantly higher ALP expression and activity mediated by these treatments as compared with the non-ALP inducing GM-treated cultures. These data conflicted with the results obtained in 2D hPDC cultures of this study that may be associated with the three dimensional character of the hPDC cultures with Ca2+ and Pi supplemented-OM− on ALP secretion. 62 In fact, production of ALP is necessary to facilitate physiological mineralization of the extracellular matrix that is highly enriched in Col-1–a physiological calcification process exclusive to bone-matrix formation. 63 Interestingly, 3D mineralized matrix deposition was mainly found at the corner of the scaffold struts, which may be a 3D-induced high elastic strain energy storage, caused by tensile deformation of the stretched strut-bridging collagen fibers that provide possible nucleation sites for biological apatite growth and propagation. 64 Microscopically, the deposited CaP crystal structures of the mineralized matrix were morphologically different that in turn may influence the physicochemical properties of the deposited CaP minerals (including the ions dissolution kinetics) and thus also the bioactivity of these mineralized structures in relation to bone induction. 23
The in vitro production of these lab-scale size osteogenic hybrids driven by bio-instructive Ca2+/Pi supplemented media may become part of a clinical-scale manufacturing strategy for reparative units. Of course, as bone is a dynamic mineralized tissue that is actively remodeled and highly vascularized, the production of merely an osteogenic constructs in vitro (without seeing the full cascade of events of osteogenic differentiation) is only a first step and still requires proof of its in vivo bone forming capacity. Therefore, ectopic nude mice models and load-bearing orthotropic large animal models will be used to assess the clinical relevance of these hybrids as bone forming units (Fig. 8). In the case that this first approach is not directly successful, the in vitro produced osteogenic hybrids can also be decellularized to obtain a biomimetic osteogenic carrier with a pregenerated osteogenic extracellular matrix that can serve as an osteoinductive and cell-customized template to be seeded with a critical mass of fresh osteogenic cells. The above strategies can also be part of a feedback-loop approach to optimize the in vitro hybrid process until a critical cell density is reached and in vivo osteoinductivity is shown. We anticipate, based on the current data, that no single universal ion supplemented medium can promote the complete cascade of osteogenic events and that most likely several specific culture medium supplements in a specific sequence will become part of the above strategy. 12 This will likely result in an effective and complete induction of the distinct stages of osteogenesis, taking into account cell type and species customisation. 43
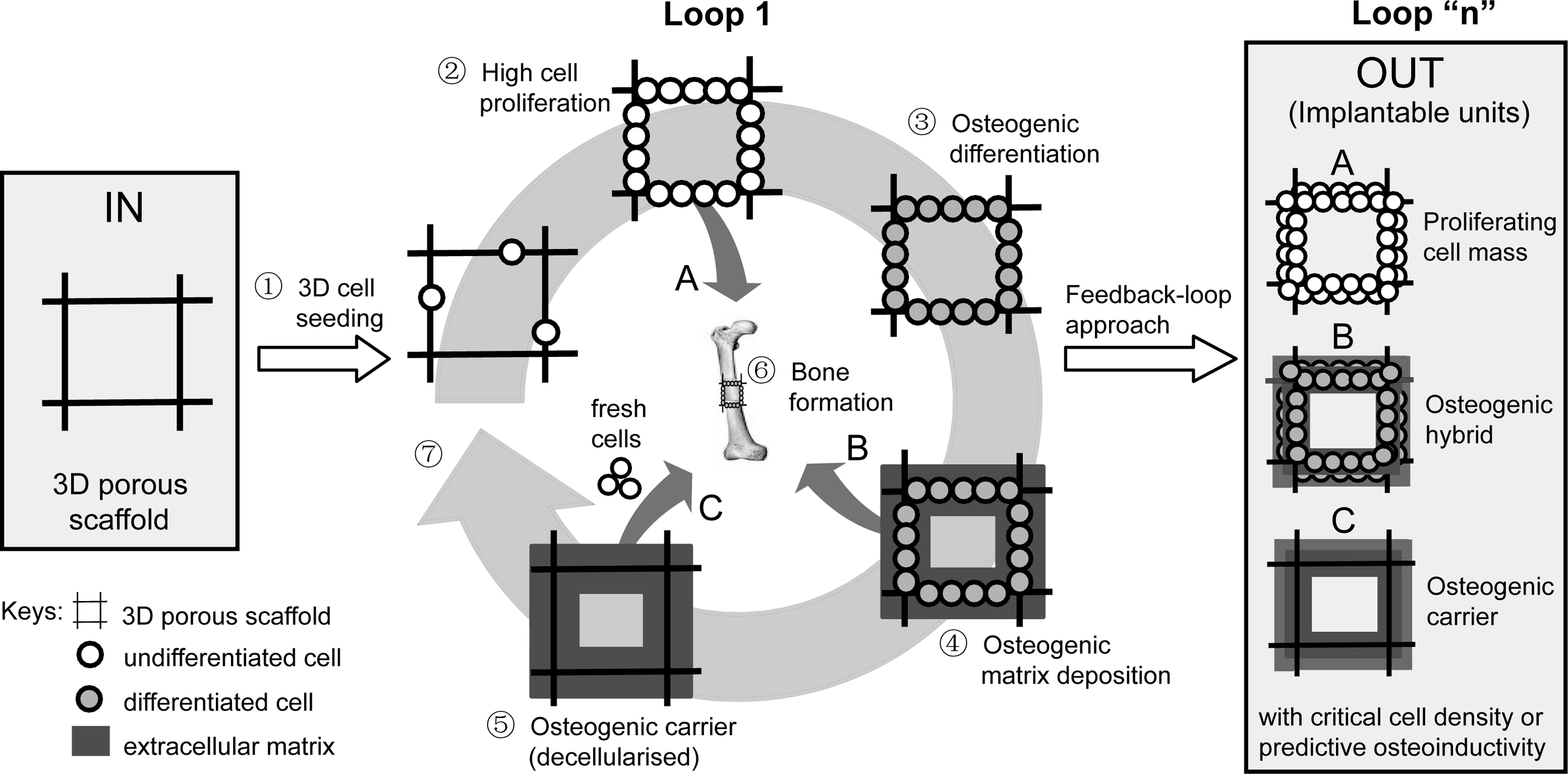
Schematic presentation of the in vitro biomimetic engineering strategy of a 3D osteogenic hybrid/carrier driven by bio-instructive Ca2+/Pi supplemented media for effective bone regeneration by manufacturing three types of implantable units
Conclusions
Through a systematic analysis of in vitro 2D to 3D biological behavior, we have gained valuable insight into the effect and interaction of Ca2+ and Pi supplementation in osteogenic inductive media on the proliferation and osteogenic differentiation of MSC-like osteoprogenitors, that is, hPDCs. Herein, we describe a bio-instructive culture environment to direct in vitro osteogenic events, by influencing cell proliferation and differentiation and instructing the differentiating cells to produce the necessary biological building blocks three-dimensionally (i.e., generation of critical cell mass, extracellular matrix synthesis, and mineralization). This biomimetic approach and 3D assembly into a provisional osteogenic hybrid can become part of a strategy for effective bone regeneration.
Footnotes
Acknowledgments
This work is funded by the K.U.Leuven IDO project 05/009–QuEST and is part of Prometheus, the Leuven Research and Development Division of Skeletal Tissue Engineering of the Katholieke Universiteit Leuven:
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.