
Abstract
Objective:
Articular cartilage has a poor capacity for spontaneous repair. Tissue engineering approaches using biomaterials and chondrocytes offer hope for treatments. Our goal was to test whether collagen sponges could be used as scaffolds for reconstruction of cartilage with human articular chondrocytes. We investigated the effects on the nature and abundance of cartilage matrix produced of sequential addition of chosen soluble factors during cell amplification on plastic and cultivation in collagen scaffolds.
Design:
Isolated human articular chondrocytes were amplified for two passages with or without a cocktail of fibroblast growth factor (FGF)-2 and insulin (FI). The cells were then cultured in collagen sponges with or without a cocktail of bone morphogenetic protein (BMP)-2, insulin, and triiodothyronine (BIT). The constructs were cultivated for 36 days in vitro or for another 6-week period in a nude mouse–based contained-defect organ culture model. Gene expression was analyzed using polymerase chain reaction, and protein production was analyzed using Western-blotting and immunohistochemistry.
Results:
Dedifferentiation of chondrocytes occured during cell expansion on plastic, and FI stimulated this dedifferentiation. We found that addition of BIT could trigger chondrocyte redifferentiation and cartilage-characteristic matrix production in the collagen sponges. The presence of FI during cell expansion increased the chondrocyte responsiveness to BIT.
Introduction
For successful MACI, a sufficient number of autologous chondrocytes should produce abundant cartilaginous matrix, but the number of chondrocytes that are isolated from the collection site should be limited even if this site is not weight bearing. It is therefore crucial to amplify the cell reservoir for clinical purposes, but dedifferentiation of chondrocytes occurs during the long-term culture necessary for mass cell production 2 . The cells start to express proteins typical of fibrous tissues such as type I collagen rather than hyaline cartilage proteins. This loss of cell function leads to the formation of a fibrocartilage biomechanically inferior to the original hyaline cartilage. To overcome this limitation, it is therefore necessary to find optimal culture conditions that allow amplification of human chondrocytes and their subsequent chondrogenic reprogramming. In this context, growth factors play an indispencible role in the steps of cell amplification and chondrogenesis induction. More precisely, during chondrocyte amplification growth factors can influence not only cell proliferation, but also cell potential to subsequently redifferentiate into a suitable environment. Several studies have indicated that fibroblast growth factor (FGF)-2 presents such a dual effect on human chondrocytes3–5 . Regarding more specifically the step of chondrogenic induction, several bone morphogenetic proteins (BMP), including BMP-2, -4, -6, -7, -9, and -13, stimulate cartilage matrix macromolecule synthesis by articular chondrocytes 6 , revealing their potential to promote cartilage repair. We previously showed that BMP-2 can stimulate chondrogenic expression in human articular chondrocytes during their amplification or after their amplification on plastic without any sign of hypertrophic maturation or osteogenic expression6,7. This indicates that the chondrogenic properties of BMP-2 can specifically be used for the therapeutic support of the adult chondrocyte phenotype. Addition of FGF-2 with insulin during amplification on plastic followed by addition of BMP-2 with insulin and triiodothyronine (T3) (designated BIT) during culture in collagen gel was found to be an optimal combination for chondrogenic expression of human auricular chondrocytes in collagen gels 8 . These exciting results prompted us to investigate in detail the effects of the same combinations of soluble factors on knee articular chondrocytes during their amplification on plastic and culture in collagen sponges, because ACI and MACI protocols have relied on articular chondrocytes. We used sponges rather than gels because we found that collagen gels but not collagen sponges trigger induction of two catabolytic enzymes of cartilage, the metalloproteinases (MMP)-1 and MMP-13, in bovine articular chondrocytes 9 . This event, rarely evaluated in studies on scaffold biomaterials for chondrocyte cultures, suggests that collagen sponges could be better scaffolds than collagen gels in term of cartilage stability. In view of clinical application, we used collagen sponges that are already in use for human skin regeneration but have not been tested for cartilage repair. Special attention was given to the newly synthesized matrix in the sponges through careful analysis of its protein composition. To further investigate the feasibility of using cartilage reconstructed in collagen sponges for clinical application, we implanted the collagen sponge–chondrocyte constructs cultured under different conditions in human osteochondral blocks. The samples were then transplanted into the subcutaneous pouches of nude mice and cultivated for 6 weeks before histologic and immunohistochemical examination.
Materials and Methods
Cell amplification
Primary cultures of human chondrocytes were prepared from macroscopically healthy zones of osteoarthritic knee joints obtained from 10 donors (age range 51-79) undergoing total knee replacement. The study was performed in full accordance with local ethics guidelines, and cartilage samples were collected after written informed consent of the donors was obtained. Chondrocytes were extracted as previously described 7 . Briefly, small slices of cartilage were sequentially digested in culture medium consisting of Dulbecco's modified Eagle medium/Ham's F12 (Gibco-Invitrogen) with 0.2% trypsin (Sigma) for 30 minutes at 37°C, followed by 0.06% bacterial collagenase A (Roche Applied Science) overnight. The cells were then seeded at a density of 1.5×104 cells/cm2 in culture medium supplemented with 10% newborn calf serum (NCS) (Hyclone), 50 μg/ml streptomycin (Panpharma, and 2 μg/mL amphotericin B (Bristol Myers Squibb). This medium for monolayer culture was refered as control medium (CTRm). At this stage, cells were designated P0. Thirty-six hours after seeding (P0-36h), medium was changed and was supplemented or not with 5 ng/mL FGF-2 (R&D Systems) and 5 μg/mL insulin (Umuline Rapide, Lilly). The cocktail of FGF-2 and insulin was designated FI. The culture medium containing FI or not was replaced 3 times per week until cells reached confluence. The cells were then trypsinized and replated at the same density for another passage (P1). After each trypsinization, cells were stained with Trypan blue; those that did not take up the dye were counted with a hemocytometer, and growth rate was calculated.
Cell culture in collagen sponges
Collagen sponges composed of native type I (90-95%) and type III (5-10%) collagen were prepared from calf skin (Symatese Biomatériaux, Chaponost, France). They were cross-linked to increase their stability using glutaraldehyde and sterilized using 25 kGy gamma radiation. They were then punched using a skin biopsy punch (Laboratoires Stiefel) into discs 5 mm in diameter and 2 mm thick (corresponding to a volume of 0.04 cm3). Their pore size was approximtely 100 nm. Passaged P1 chondrocytes contained in 10 μL of culture medium were seeded onto the sponges with 13×106 cells/cm3, and the sponges were placed in 48-well culture plates and incubated at 37°C for 2 hours. One milliliter of culture medium containing 10% NCS and supplemented with 50 μg/mL 2-phospho-L-ascorbic acid (trisodium salt, Fluka) was then added to the wells in the absence or presence of 200 ng/mL of recombinant human BMP-2α (Dibotermine-alpha, drug form of BMP-2 contained in the kit InductOs, Wyeth), 5 μg/mL insulin (Umulin, Lilly), and 100nM thyroxin T3 (Sigma). The cocktail of BMP-2, insulin, and T3 was designated BIT. The culture medium containing 10% NCS and ascorbate was referred to as control medium for the culture in scaffold (CTRs). Medium was replaced every 2 days over a culture period of 21 or 36 days.
Implantation of collagen sponges in osteochondral blocks
Osteochondral blocks were punched using 10-mm-diameter hollow punches (reference 694-C-320-N, SAM Outillage, France,
Gene expression analysis using reverse transcription polymerase chain reaction
Total RNA was isolated from the chondrocytes cultured in monolayer or in collagen sponges after a 21-day culture period using the NucleoSpin RNA II kit (Macherey-Nagel) according to the manufacturer's instructions. Total RNA was then reverse transcribed as previously described 10 .
Conventional polymerase chain reaction (PCR) amplifications of the different type II isoforms transcripts were performed in a PCR Thermal Cycler MyCycler (Bio-Rad) using primers as previously described 6 (Table 1).
Primers are presented in a 5′-3′ orientation, with that for the forward strand (+) being above that for the reverse strand (-). The annealing temperature and the number of PCR cycles used for each primer set are indicated. The source of the databank used for designing the primers is presented as an accession number. When primers have been used in other studies, the references are indicated.
PCR, polymerase chain reaction.
Real-time PCR amplifications were performed as previously described 6 (Table 2). Glyceraldehyde 3-phosphate dehydrogenase Ct value was substracted from the target sequence Ct value to obtain ΔCt. The level of expression was then calculated as 2-ΔCt and expressed as the mean of triplicate samples.
Primers are presented in a 5′-3′ orientation, with that for the forward strand (+) being above that for the reverse strand (-). The source of the databank used for designing the primers is presented as an accession number. When primers have been used in other studies, the references are indicated.
Antibodies
D. J. Hartmann kindly provided polyclonal rabbit antibodies to human type II collagen (Ref 20211) 11 . Polyclonal rabbit antibodies to aggrecan were from Novotec (Ref 24411), to Sox9 were from Millipore (Ref AB5535), and to actin were from Sigma (Ref A5060). Monoclonal antibody to type IX collagen (23-5D1) was a gift of B. R. Olsen. 12 Secondary antibodies were alkaline phosphatase–conjugated anti-rabbit or anti-mouse immunoglobulin (Ig)G (Bio-Rad) and horseradish peroxidase (HRP)-conjugated anti-rabbit IgG (Cell signaling for Western blotting assays and Vector ImmPRESS kit for immunohistochemistry experiments).
Western-blot analysis
After 36 days of in vitro culture, collagen sponges were frozen in liquid nitrogen. After being thawed, they were rinsed with phosphate-buffered saline containing 2mM Ethylenediaminetetraacetic acid and 0.2mM phenylmethylsulfonyl fluoride, refrozen, ground with a mostar and pestle into a fine powder, and resuspended and boiled in Laemmli buffer. For Western-blot analysis, equivalent amounts of proteins were separated using sodium dodecyl sulfate–polyacrylamide gel electrophoresis on 4% to 12 % gradient gels for sponge samples and on 6 % gels for culture medium samples. After transfer, membranes were probed with primary antibodies; washed; and incubated with alkaline phosphatase–conjugated IgG for type I, II, and IX collagens and actin detection or with HRP-conjugated IgG for Sox9 detection. After multiple washes, bound antibodies were detected on X-ray films using an Immun-star AP or HRP chemiluminescent substrate (Bio-Rad). Membranes were reprobed with antibodies after stripping (Re-Blot Plus Strong, Chemicon).
Immunohistochemical analysis
Histologic examinations of the collagen sponges seeded with chondrocytes were performed after 36 days of in vitro cultivation and a subsequent 6 weeks of cultivation in osteochondral blocks implanted in nude mice. The collagen sponges cultured in vitro only were rinsed in phosphate-buffered saline and fixed for 24 hours with 4% neutral-buffered formalin. After dehydration in a graded series of ethanol baths, samples were embedded in paraffin. After retrieval of the osteochondral blocks from mice, samples were fixed for 24 hours with 4% paraformaldehyde and then decalcified for 5 days in acidic solution (Eurobio) before being embedded in paraffin. Hematoxylin and eosin counterstaining and immunohistochemical analysis was performed on 4- to 5-μm sections. Incubation with type II collagen antibodies was followed by incubation with HRP-conjugated secondary antibodies. Sections were revealed with diaminobenzidine and observed suing a DM 4000B microscope (Leica) directly coupled to a color camera (Digital Camera DXM1200, Nikon). Image acquisition was achieved using Metaview software (Universal Imaging).
Statistical analysis
Differences in cell multiplication and relative gene expression between experimental groups were analyzed using the Mann-Whitney U-test for nonparametric analysis. P<.05 was considered to be significant. The number of experiments performed is noted in the figure legends.
Results
Chondrocyte proliferation
Human articular chondrocytes were expanded under Good Manufacturing Practice (French Health Agency Afssaps no. TCG/07/M/001) conditions in the presence of 10% NCS supplemented or not with FI. The growth rate was significantly higher in the presence of FI, when calculated with chondrocytes isolated from seven donors (Fig. 1).
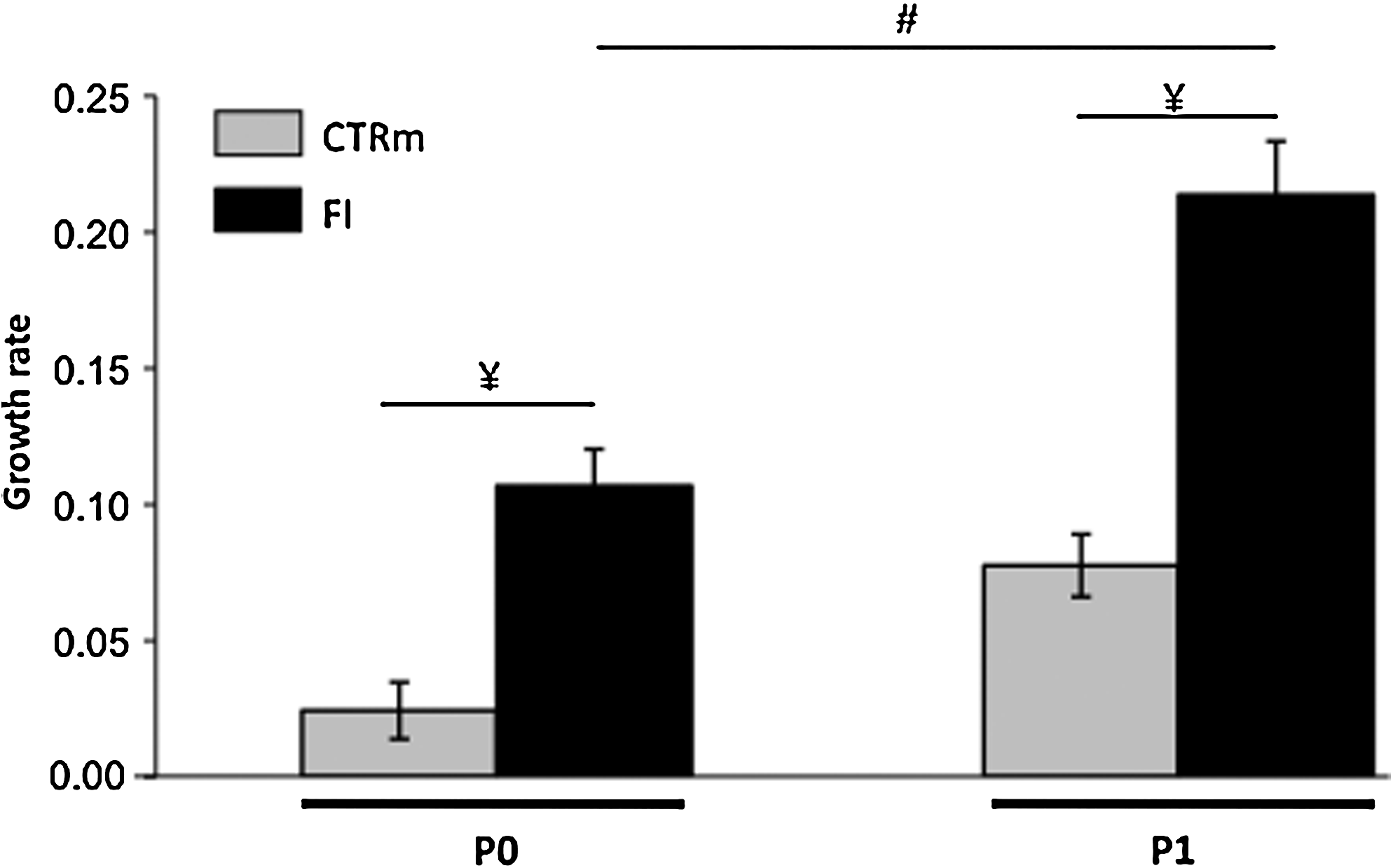
The combination of FGF2 and insulin (FI) stimulates the growth rate of human articular chondrocytes amplified on plastic. Chondrocytes at passages P0 and P1 were cultured in the presence of 10% newborn calf serum (NCS) (indicated as CTRm) or in the presence of 10% NCS supplemented with FI (indicated as FI) (n=7, ¥ p<0.01, significant effects of FI versus CTRm; # p<0.01, significant effect of FI in P1 versus FI in P0). FGF2, fibroblast growth factor 2.
Gene expression analysis in monolayer culture and in the collagen scaffolds
Gene expression analysis was performed to quantify the relative abundance of the transcripts coding for type II collagen and aggrecan, the two major proteins of cartilage and for SOX9, a transcription factor essential for COL2A1 and AGGRECAN expression regulation 13 . Human articular chondrocytes isolated from four donors were cultured separately on plastic in the presence or not of FI and then in collagen sponges in the presence or not of BIT. From P0 to P1, expansion in monolayer resulted in progressive loss of COL2A1 expression, and this decrease was particularly marked in the presence of FI for P1 chondrocytes (∼80% less with FI than under control culture conditions) (Fig. 2). During expansion in monolayer, expression of AGGRECAN and SOX9 remained relatively stable in control cultures but was noticeably lower in the presence of FI (∼60% less for P1 chondrocytes with FI than under control culture conditions). After a subsequent 21-day culture period in collagen sponges, COL2A1 was not detected when BIT was omitted from the culture medium (Fig. 2A). In contrast, COL2A1 expression was regained in the presence of BIT whatever the first culture conditions used in monolayer and to a level higher than in P0-36 h chondrocytes (4.5 times greater), although the levels of AGGRECAN and SOX9 expression measured after cell amplification on plastic did not change significantly in collagen sponges (Fig. 2B and C). Extremely low levels of expression of genes coding for type X collagen, an indicator of chondrocyte hypertrophy, and for osteocalcin, a bone-specific marker, were detected in chondrocytes 36 hours after their extraction, and these levels remained low or undetectable afterward for any cell culture condition at any time point (data not shown).
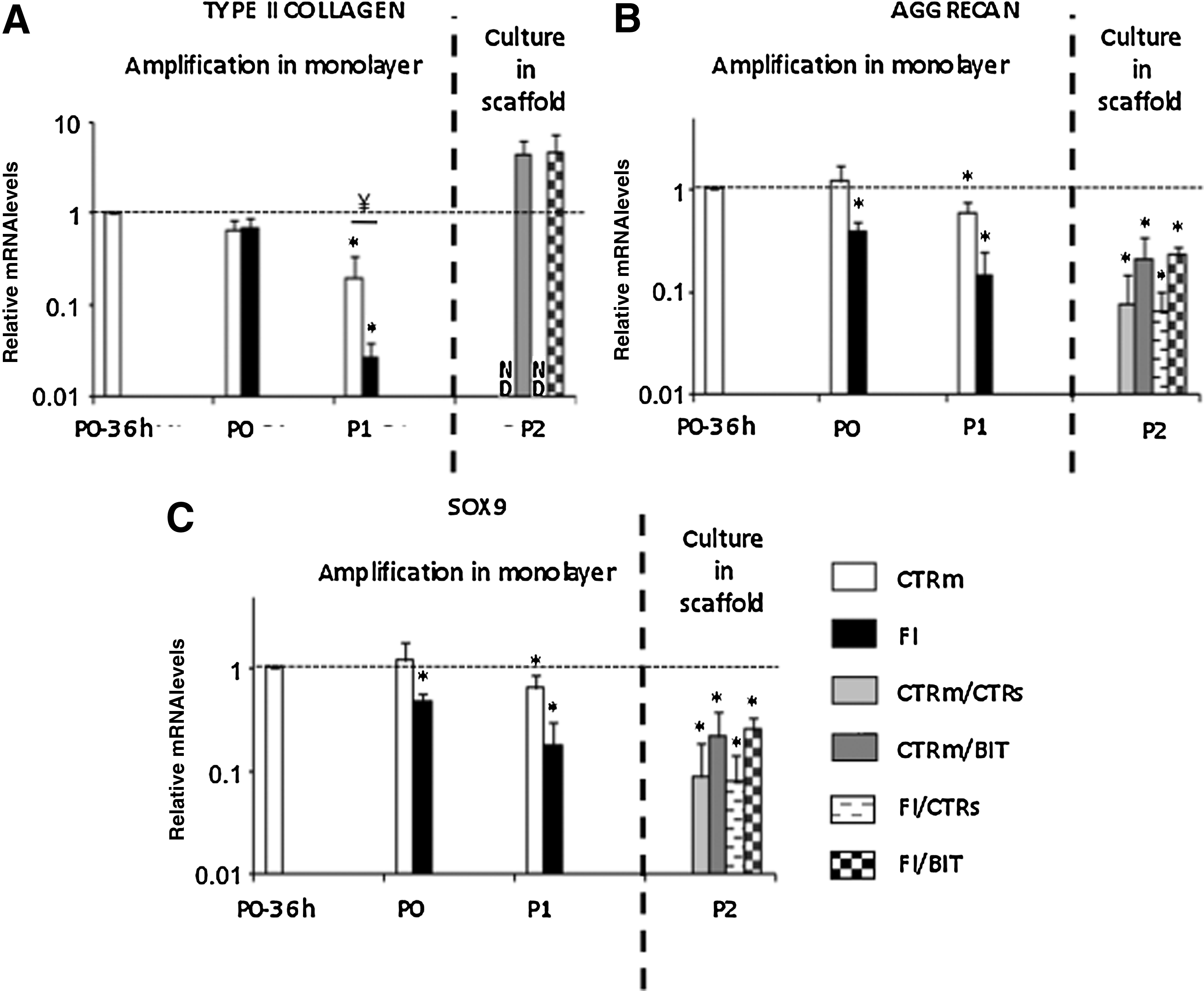
The combination of bone morphogenetic protein-2, insulin, and triiodothyronine (T3)[BIT] reinduces chondrogenic gene expression in human articular chondrocytes cultured in collagen sponges. The chondrocytes were first amplified in monolayer and then cultured in the sponges for 21 days, as indicated. The diverse culture conditions including or not the cocktails of factors are mentioned in the legends of the bars. The italic slash separates culture conditions in monolayer and culture conditions in scaffold. Real-time polymerase chain reaction analyses were performed to measure levels of messenger RNA, and the corresponding gene products are indicated
Regarding COL1A1, coding for type I procollagen α1 chain, subculturing from P0 to P1 on plastic led to greater expression, a well-known phenomenon associated with chondrocyte dedifferentiation in vitro (data not shown). Neither addition of FI nor any culture condition used afterward for the sponges influenced this difference (data not shown). We then evaluated the ratio of type II to type I procollagen messenger RNA (mRNA) levels, a differentiation index for chondrocytes 14 now widely used to monitor the differentiation status of chondrocytes in the diverse ACI or MACI protocols. The ratios measured for the collagen sponge–chondrocyte constructs cultured in the presence of BIT reached values close to the value measured in P0-36 h chondrocytes (Fig. 3A), suggesting that a certain degree of chondrocyte redifferentiation occurred in the presence of the BIT cocktail. Tthe value reached with the FI–BIT combination was not significantly different from the value measured in P0-36 h chondrocytes (Fig. 3A). To further evaluate the status of the chondrocyte phenotype, we analyzed expression of the alternatively spliced isoforms of COL2A1 mRNA, considering that the IIB isoform is the only spliced variant characteristic of well-differentiated chondrocytes, whereas precursor cells undergoing chondrogenesis express the other IIA, IIC, and IID isoforms 15 . We used primers spanning exons 1 to 7 and conventional reverse transcriptase –PCR, as described previously 6 . Our data showed that COL2A1 was transcribed almost exclusively as type IIB mRNA once chondrocytes were seeded in collagen sponges and cultured with BIT, regardless of the preliminary culture conditions used for cell amplification (Fig. 3B). Taken together, these data indicated that the BIT cocktail could trigger redifferentiation of human articular chondrocytes in the collagen sponges.
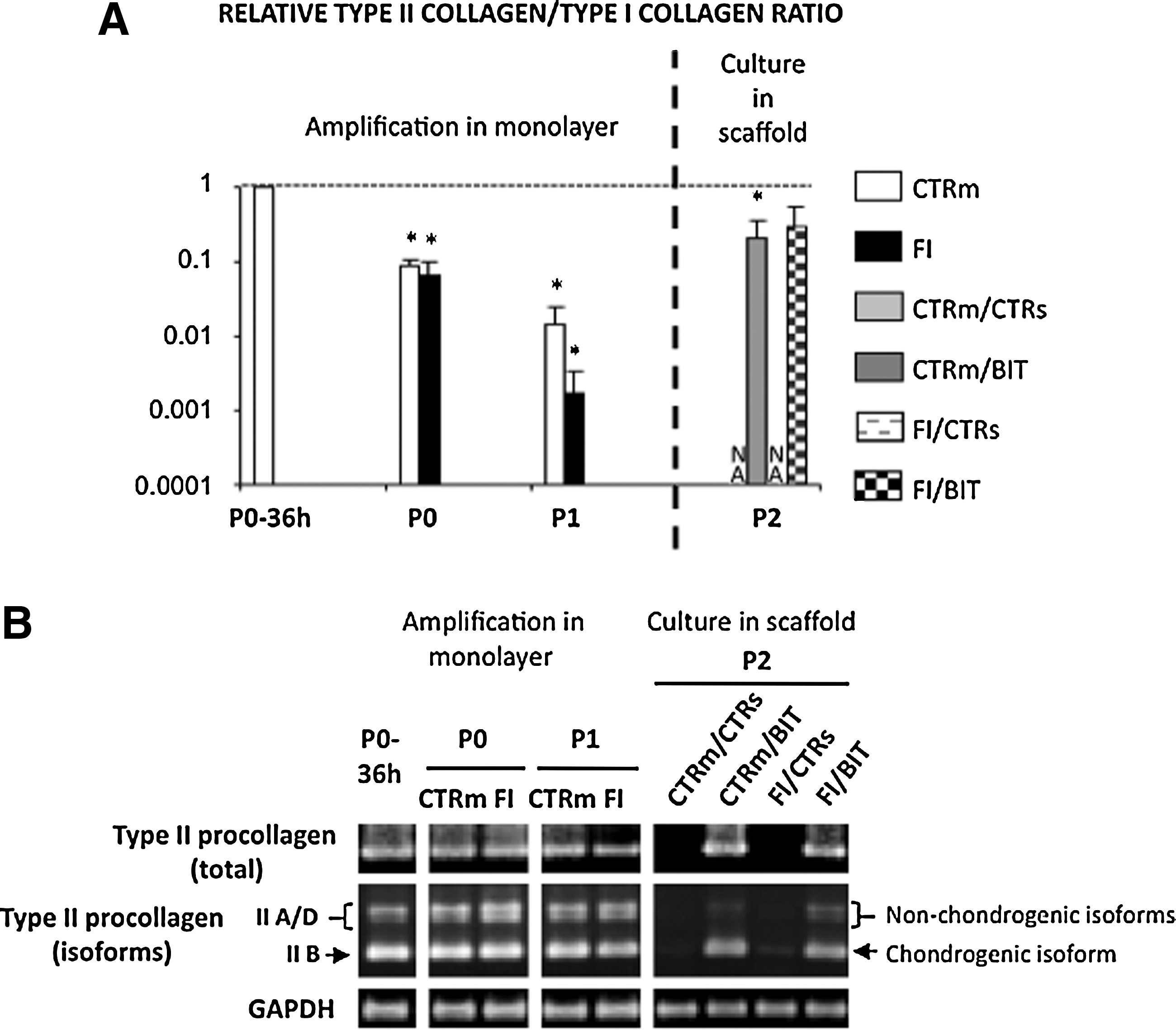
The combination of BMP-2, insulin, and triiodothyronine (BIT) reinduces cartilage-characteristic collagen expression in human articular chondrocytes cultured in the scaffolds. The chondrocytes were first amplified in monolayer and then cultured in the sponges for 21 days in presence or absence of the different cocktails of soluble factors, as indicated. The diverse culture conditions are mentioned in the legends of the bars. The italic slash separates culture conditions in monolayer and culture conditions in scaffold.
Analysis of the proteins newly synthesized by the chondrocytes in the collagen scaffolds
We next looked for cartilage-characteristic proteins synthesized and deposited by the chondrocytes in the collagen sponges. Chondrocytes isolated from six donors were cultivated separately on plastic and then in collagen sponges under the diverse culture conditions described previously. The constructs were divided into two sample sets: one for the analysis using Western-blot and immunohistochemistry after 36 days of in vitro cultivation and one for immunohistochemistry analysis after subsequent cultivation for 6 weeks in osteochondral blocks implanted in nude mice. The protein analyses were performed after 36 days of in vitro cultivation because accumulation of proteins with time facilitated their detection.
When immunolocalization was used to assess the morphologic distribution of type II collagen in the scaffolds cultured in vitro, strong staining was observed in the constructs treated with BIT whereas no staining could be detected in the other scaffolds (Fig. 4). Type II collagen was found to accumulate preferentially at the periphery of the BIT-treated scaffolds, but the distribution of type II collagen staining was more homogenous and spread deeper when chondrocytes were first amplified on plastic in the presence of FI (Fig. 4). Similar distribution of aggrecan was seen with anti-aggrecan antibodies in the BIT-treated constructs, whereas aggrecan remained undetectable in the other sponges (data not shown).
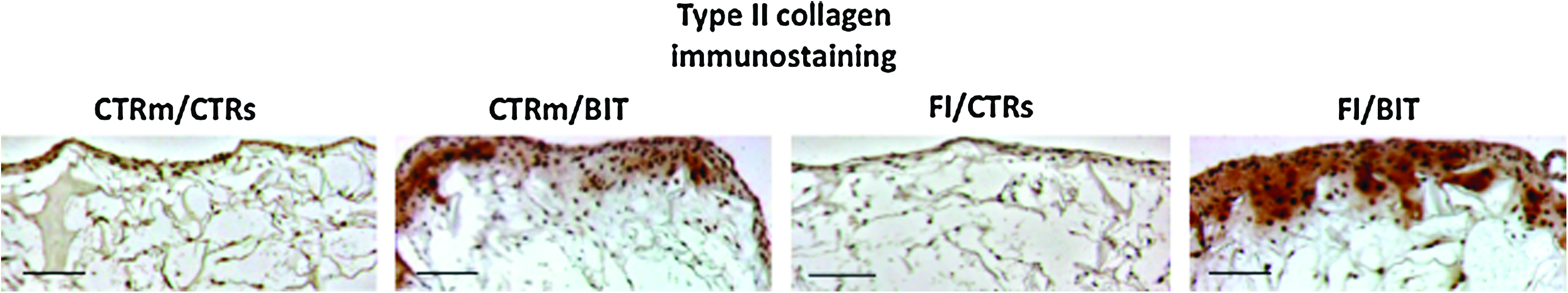
Immunohistochemical analyses of the scaffolds cultured for 36 days in vitro. The human articular chondrocytes were first amplified in monolayer and then cultured in collagen scaffolds in the presence or not of the cocktails of factors, as indicated. The italic slash separates culture conditions in monolayer and culture conditions in scaffold. Immunostaining for type II collagen shows a strong signal in BIT- constructs treated with a cocktail of bone morphogenetic protein-2. insulin, and triiodothyronine, particularly around the chondrocytes, suggesting accumulation of this type of collagen. These results are representative of cultures of chondrocytes isolated from three donors (scale bar: 200 μm). Color images available online at
We next extended our study with a more-detailed identification of important markers of cartilage matrix using Western-blot analysis of the proteins newly synthesized by the chondrocytes in the collagen scaffolds. When the different combinations of culture conditions used for cell expansion and culture in scaffolds in vitro were compared, our results revealed that sequential combination of FI and BIT led to the highest level of type II collagen synthesis (Fig. 5). Covalently cross-linked dimers and trimers of type II collagen molecules could be detected on our Western blots, suggesting the presence of a fibrillar network (Fig. 5). In addition, cell cultivation with a FI–BIT combination resulted in the highest levels of synthesis of Sox9 and type IX collagen, another typical component associated with the collagen fibrils in cartilage (Fig. 5). Faint bands corresponding to type II and IX collagens and to Sox9 were also revealed when the scaffolds were cultured in the presence of BIT, after cell expansion in control conditions, whereas these proteins were not detected when the other combinations of culture conditions were used (Fig. 5).

Sequential exposure of the chondrocytes to the cocktail of fibroblast growth factor-2 and insulin (FI) for amplification on plastic and to the cocktail of bone morphogenetic protein-2, insulin, and triiodothyronine (BIT) for cultivation in collagen scaffold leads to robust expression of cartilage-characteristic proteins in the scaffolds. Western-blotting analysis of type II and IX collagens and of Sox9 after a 36-day culture period in the scaffolds. The cell culture conditions are indicated with an italic slash to separate culture conditions in monolayer and culture conditions in scaffold. On the right, the positions of mature type II collagen chains (mature) and unprocessed (pro) or processing intermediate of type II procollagen–containing amino-propeptide (pN) are indicated. The upper bands represent dimers (β) and trimers (γ) of type II collagen molecules (the trimers are more or less visible depending on the blots). At the bottom of the panel, immunoblotting for actin is shown as a loading control. The positions of molecular mass markers are indicated on the left. These results are representative of cultures of chondrocytes isolated from three donors.
Aiming at regenerating cartilage that is applicable for clinical use, we next investigated the cartilage constructs in human osteochondral blocks transplanted subcutaneously into nude mice. The histologic examination of the retrieved samples after 6 weeks of subcutaneous cultivation revealed that the matrix-based defect treatment led to complete filling of the defects (Fig. 6). The interface zone between repair tissue and existing cartilage showed good integration for all the samples (Fig. 6). The chondrogenic character of the tissue constructs was then examined by immunostaining with antibodies against type II collagen. Marked staining was oberved on tissue sections of the constructs treated with BIT, regardless of the first cell expansion culture conditions (Fig. 7B and D). Density of staining in the BIT-treated constructs was similar to that of normal existing cartilage, and this staining was more homogeneously distributed throughout the constructs when the cells were first amplified in the presence of FI (Fig. 7B and D). In addition, type II collagen was not detected using immunohistochemistry in the other types of constructs (Fig. 7A and C). In parallel, weak immunostaining with anti-aggrecan antibodies could be detected in the constructs treated with BIT (data not shown).
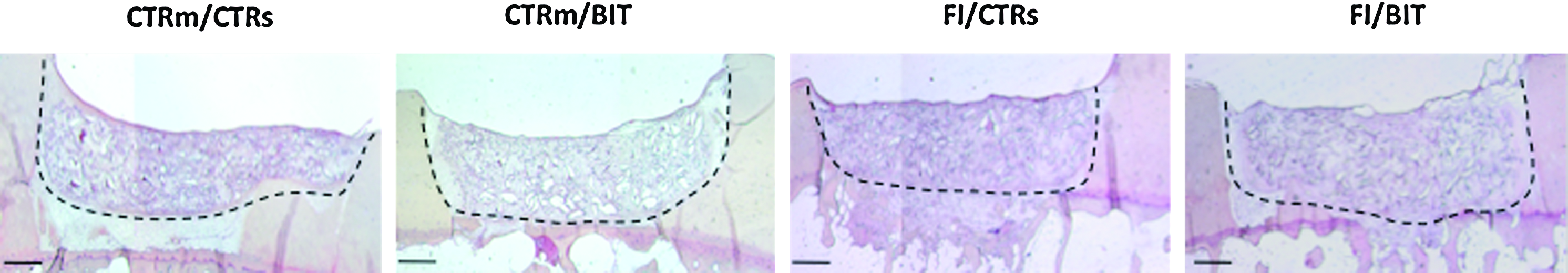
Defects of osteochondral blocks treated using the different types of chondrocyte–scaffold constructs. After in vitro cultivation for 36 days, collagen sponge–cell constructs were implanted into defects created at the surface of osteochondral blocks. The blocks were then implanted into the subcutaneous pouches of nude mice. Hematoxylin and eosin staining after 6 weeks of subcutaneous cultivation showed good integration for all the samples. The dotted lines indicate the interface zone between repair tissue and existing cartilage. The cell culture conditions indicated correspond to the in vitro culture conditions as described in the other figures. Color images available online at
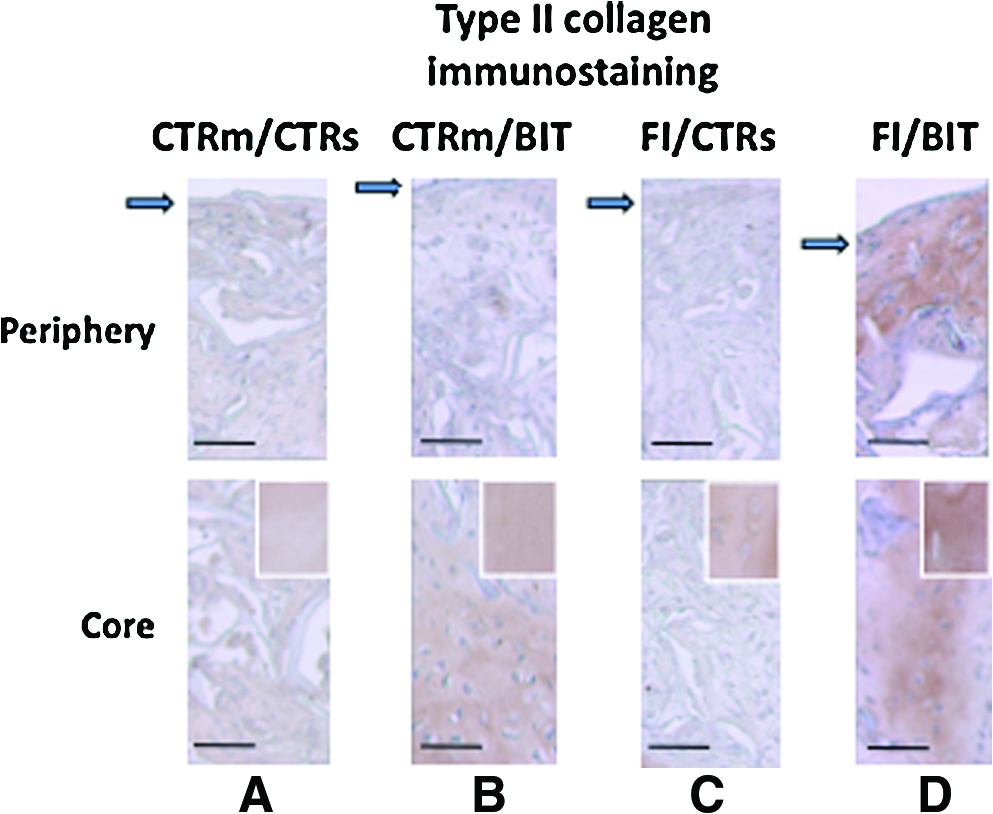
Type II immunostaining of the constructs implanted in osteochondral blocks, as described in Figure 6. The cell culture conditions indicated correspond to the in vitro culture conditions described in the other figures. On the top, peripheral zone of the scaffolds. Arrows indicate the surface. At the bottom, core of the scaffolds. For comparison, inserts show density of type II immunostaining in the existing cartilage supporting the scaffold (scale bar: 100 μm). Color images available online at
Discussion
The present study was undertaken to develop a tissue engineering strategy for regeneration of permanent cartilage using human articular chondrocytes, collagen scaffolds, and bioactive molecules. We chose to use combinations of FGF-2 and insulin for cell amplification on plastic and BMP-2, insulin, and T3 for chondrogenic induction in collagen sponges, because these combinations have been shown to be optimal to generate tissue-engineered cartilage from human auricular chondrocytes cultured in three-dimensional atelocollagen gels 8 . The first goal that had to be achieved was to increase the number of cultured chondrocytes for the cartilage regenerative medicine. In the present study, human articular chondrocytes were amplified under Good Manufacturing Practice. Our control conditions for cell expansion contained 10% NCS to mimic the cell amplification procedure used in our clinical trial of ACI at Lyon Sud Hospital (Centre Hospitalier Lyon Sud, Hospices Civils de Lyon, CHLS, HCL) 16 . The synergistic effects of FGF-2 with insulin on the proliferation of human auricular chondrocytes was previously demonstrated, 17 and we confirmed the positive effect of FI in the human articular chondrocytes and proved its usefulness in combination with NCS. Although FGF-2 and insulin are mitogenic to chondrocytes18,19, their exact mechanisms of action on cell cycle and DNA synthesis remain to be unravelled. The present study also shows that FI stimulates dedifferentiation of the chondrocytes, as judged in particular by the significant decrease in expression of the gene coding for type II collagen, the major protein of cartilage. It has previously been shown that human dedifferentiated chondrocytes could reexpress their differentiated phenotype upon transfer into three-dimensional matrices such as alginate beads,20–22 and this chondrogenic reexpression could vary widely depending on the nature of the serum used to supplement the culture medium 23 . Here, our results indicate that cultivation in collagen sponges in the presence of 10% NCS is not sufficient to achieve efficient redifferentiation of human articular chondrocytes, whatever the expansion conditions. The nature of the serum or of the scaffold used in this study could explain the lack of ability of the chondrocytes to recover their differentiated phenotype in these conditions. It has recently been shown that the type of actin organization associated with a round cellular shape controls chondrocyte-specific gene expression 24 . The collagen sponges are porous matrices that provide large spaces between the collagen walls at the cellular scale. Thus, this type of scaffold, unlike agarose or alginate gels, does not induce cell rounding. Our preliminary studies at the gene level univocally indicated that addition of BIT was responsible for redifferentiation of the chondrocytes in collagen sponges.
Our analyses at the protein level confirmed the efficiency of the BIT cocktail to induce cartilage-characteristic matrix production in the collagen scaffolds. In particular, our Western-blotting analysis confirmed the effect of FI for cell expansion on the abundance of the subsequent cartilage protein production in the collagen sponges. Our immunohistochemical analyses of the collagen sponges cultured in vitro and after implantation in nude mice were globally in agreement with Western-blotting data. The immunostaining for type II collagen was consistently more intense and more widespread throughout the scaffolds when chondrocytes were sequentially exposed to the FI and BIT cocktails. The densities of the staining observed in the sponges after in vitro cultivation or after subsequent implantation in vivo should not be compared because tissue processing of osteochondral blocks involves a decalcification step that may affect antigen exposure. In an attempt to evaluate the degree of cartilage reconstruction in the scaffolds, we internally controlled each section of the blocks by comparing the density of type II collagen staining in the repair tissue with that of normal adjacent cartilage on each slide. It was apparent that FI and BIT was the most advantageous combination for human articular chondrocytes to build a cartilaginous matrix within the collagen sponges.
Taken together, the results of the present study indicate that the same combination of soluble factors (FI and BIT) used in a previous study to generate cartilage from auricular chondrocytes in atelocollagen gels 8 is also efficient in generating cartilage from articular chondrocytes in collagen sponges. This cartilage reconstruction occurs without hypertrophic maturation or osteogenic transdifferentiation of the chondrocytes. We found that BIT stimulated Sox9 expression in the sponges, whereas the previous study revealed an inhibitory effect of BIT on Sox9 mRNA levels in the atelocollagen gels 8 , although gene expression and synthesis of Sox9 were analyzed after 21 and 36 days of culture in our study, whereas Sox9 mRNA levels were evaluated after 7 days of culture in the study of Liu et al. 8 Thus, further time course analyses of Sox9 expression at the gene and protein level are needed to determine whether the opposite effects of BIT observed on Sox9 expression in the two studies reflects different signaling mechanisms of chondrocyte redifferentiation. Although we did not evaluate here the biomechanical properties of the reconstructed cartilage, the presence of cross-links in the newly formed matrix is a sign that chondrocytes were able to synthesize enzymes necessary for correct maturation and stabilization of collagen molecules and their packing into collagen fibrils. Maturation and cross-linking of the collagen network contributes to the functional properties of the articular cartilage. In the present study, we also observe that the cartilage matrix built in the sponges cultured in static conditions in vitro was not homogenously distributed (cell distribution and matrix mainly at the periphery of the scaffold), but this drawback could be overcome using a perfusion bioreactor. Nevertheless, the chondrocytes maintained their capacity to produce extracellular matrix during cultivation of the sponges in vivo. In this case, when cells were exposed to the FI–BIT combination, we noticed that the staining of the matrix appeared more widely distributed throughout the repair tissue. This observation indicates that constructs continue to mature during the implantation period in nude mice. Although the mechanical properties and other biologic characteristics under the skin do not exactly match the conditions in a normal joint, the constructs remained well integrated in the surrounding cartilage after the 6-week implantation period. Thus, the results of this study using a selection of soluble factors and a true human organ culture model suggest that collagen sponges are suitable biomaterials for cartilage engineering. Moreover, collagen sponges are easy to shape and calibrate. This last point is important for obtaining the best possible filling of the cartilage defect, like in developing OA. What is more, collagen sponges can be grafted using minimally invasive arthroscopic surgery because they can be sealed with fibrin glue.
Footnotes
Acknowledgments
This work was financially supported by the Centre National de la Recherche Scientifique, Université Lyon 1, and ANR TecSan (2006 Promocart program). We thank Novotec (Lyon) for the histologic preparations and the technical facilities of IFR 128 (BioSciences Gerland-Lyon Sud) for the quantitative PCR analyses and animal housing at the Plateau de Biologie Expérimentale de la Souris.
Disclosure Statement
No competing financial interests exist.