
Abstract
Tissue engineering approaches using the combination of porous ceramics and bone marrow mesenchymal stem cells (BMSCs) represent a promising bone substitute for repairing large bone defects. Nevertheless, optimal conditions for constructing tissue-engineered bone have yet to be determined. It remains unclear if transplantation of predifferentiated BMSCs is superior to undifferentiated BMSCs or freshly isolated bone marrow mononucleated cells (BMNCs) in terms of new bone formation in vivo. The aim of this study was to investigate the effect of in vitro osteogenic differentiation (β-glycerophosphate, dexamethasone, and
Introduction
Since the first report of the existence of bone marrow mesenchymal stem cells (BMSCs) by Friedenstein et al. in 1968, 3 the multilineage potential and self-renewal capability of BMSCs have been demonstrated extensively both in vitro and in vivo. Under appropriate inductive conditions, BMSCs can be directed into osteoblasts that can regenerate bone tissue, and many studies have succeeded in repairing bone defects using BMSCs as seed cells in animal models as well as in clinical trials.4–12
Because of its excellent osteoconductivity and biocompatibility, calcium phosphate-based bioceramics seem to be a promising candidate as the bone tissue engineering scaffold. Goshima et al. were among the first to demonstrate new bone deposition in porous calcium phosphate ceramics seeded with BMSCs, once the cell/scaffold composites were implanted subcutaneously into syngeneic rats or athymic mice.13–15 After that study, other authors have obtained equivalent results in similar models using BMSCs from different species.16–22 We and other groups demonstrated the effective 3D culture of BMSCs on calcium phosphate ceramics, and have healed osseous defects using a tissue engineering approach in large animals like canines and goats.8,10,20,22
However, before BMSC-based bone tissue-engineering therapies can be widely used in the clinical arena, some optimal conditions for BMSCs' in vitro expansion, osteogenic differentiation, and 3D co-cultivation with the bioceramic scaffolds remain to be addressed. For example, it is not known whether scaffolds seeded with bone marrow mononucleated cells (BMNCs) without being introduced into cell culture are equivalent or superior to those containing in vitro expanded BMSCs in terms of in vivo new bone formation.23,24 Additionally, it has yet to be determined whether prior in vitro osteogenic induction of BMSC/scaffold composites can facilitate the subsequent bone formation in vivo. Some groups demonstrated that ectopic tissue-engineered bone formation was dependent on the osteogenic co-culture of BMSCs combined with calcium phosphate scaffolds before transplantation.17,19,20 There are, in contrast, also several studies utilizing undifferentiated BMSC/scaffold composites to regenerate bone tissue at subcutaneous sites in rats or nude mice13,14 and to repair bone defects in large animals.4–6 Niemeyer et al. found that in vitro osteo-differentiation of BMSCs in scaffolds containing calcium phosphate and collagen type I did not promote the in vivo BMSC-associated bone formation, 25 and Zong et al. showed that bone reconstruction potential of undifferentiated BMSC/PLGA was superior to that of osteo-induced BMSC/PLGA composites in a rat calvarial defect model. 26
These above-mentioned reports mainly focused on whether the differentiation of the cell/scaffold composite is needed before implantation, but little work has been done to investigate the influence of in vitro osteogenic induction of BMSCs before cell seeding and transplantation on the potential of ectopic bone regeneration. In vitro co-culture of the cell/scaffold composites is a cost-intensive and time-consuming procedure for clinical use. If the implanted scaffolds seeded with predifferentiated cells without 3D co-cultivation prove to be equally effective, this will ease the introduction of bone tissue-engineering into clinical practice.
We hypothesized that BMSCs already committed to osteoblasts are capable of more rapid in vivo bone formation than BMNCs and undifferentiated BMSCs. To test this hypothesis, porous beta-tricalcium phosphate (β-TCP) ceramics seeded with freshly isolated human BMNCs, in vitro expanded undifferentiated and osteo-differentiated BMSCs, respectively, were implanted subcutaneously into athymic mice, and their ectopic bone regeneration was compared. A nude mouse model of heterotopic implantation was chosen for this study because it allows the study of the influence of in vitro manipulation before implantation in the absence of local osteoinductive influences, such as growth factors and mechanical loading.16,25
Materials and Methods
Experimental design
BMSCs were isolated from human bone marrow harvested from six healthy donors, expanded, and osteogenically induced in vitro for three passages. Cells from each donor were seeded onto five β-TCP scaffolds and then surgically implanted into the subcutaneous sites of nude mice. For the control groups, freshly isolated BMNCs and noninduced BMSCs from the same donor were also loaded onto five β-TCP ceramics, individually. In vivo bone formation was evaluated by micro-CT and histology at 4, 8, 12, 16, and 20 weeks postimplantation. So at each time point, six implants were harvested from each group (BMNCs+β-TCP group, noninduced BMSCs+β-TCP group, and osteo-induced BMSCs+β-TCP group), and each implant represented cells from one donor.
Isolation of human BMNCs
All protocols for human tissue handling were approved by the Research Ethics Committee of TongJi University. After informed consent, bone marrow aspirates (30 mL) were obtained from the iliac crest of six hematologically healthy donors (aged 13–69 years) during routine plastic surgical procedures involving exposure of the iliac crest, and added into heparinized tubes (containing 500 units of sodium heparin per milliliter marrow). Human BMNCs were isolated by using the density gradient centrifugation method as previously described, 27 with minor modifications within 12 h from harvest. Briefly, each 10 mL of bone marrow aspirates was diluted with Dulbecco's phosphate-buffered saline (DPBS; Sigma, St. Louis, MO) at a ratio of 1:2, and the buffy coat was obtained from bone marrow by centrifugation (1500 rpm for 20 min). The buffy coat cells were then were layered onto the Percoll solution (1.073 g/mL; Pharmacia, Piscataway, NJ) and centrifuged (1100 g for 30 min at 20°C). After centrifugation, the low-density mononucleated cells (BMNCs, <1.073 g/mL) in the interface were collected and the cell number was counted with a hemacytometer.
In vitro expansion, osteogenic induction, and characterization of BMSCs
BMNCs from each donor were divided into three parts (i.e., each part from 10 mL bone marrow aspirates); one part was used to seed onto the porous β-TCP scaffolds immediately, as will be described below. Two parts were seeded in 100-mm tissue culture dishes (Falcon, B&D, San Jose, CA) at a density of 1×105 cells/cm2 and cultured either in basic culture media (low-glucose Dulbecco's modified Eagle's medium [Gibco, Grand Island, NY] supplemented with 10% fetal bovine serum [FBS, Gibco], 2 mmol/L L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin sulfate [all from Sigma]) or in osteogenic medium (consisting of basic culture medium supplemented with 10 mM β-glycerophosphate [β-GP], 10−8 M dexamethasone, and 50 μM
The phenotypic characterization of human BMSCs at passage 2 were performed by flow cytometry on a FACScan (Coulter Epics Altra; Becton Dickson, San Jose, CA) with specific fluorescein isothiocyanate-conjugated or phycoerythrin-conjugated monoclonal antibodies, including CD14, CD29, CD31, CD34, CD45, CD73, CD79a, CD90, CD105, CD166, HLA-ABC, and HLA-DR. The multilineage potential of BMSCs to differentiate into osteoblasts, adipocytes, and chondrocytes was also confirmed in monolayer culture or in pellet culture as previously described.27,28
Analysis of cell proliferation and osteogenic differentiation
After trypsinization of the primary culture (P0), obtained BMSCs were replated on 25-mm tissue culture dishes at a density of 1×105 cells per dish, and passaged similarly upon reaching next confluence. The cells were expanded in the basic culture medium or induced in the osteogenic medium for three passages (P0, P1, and P2; on average 20–25 days), and both the osteo-induced and noninduced BMSCs of P2 were homogenized with sonication and analyzed for DNA content, alkaline phosphatase (ALP) activity, and calcium content to characterize cell proliferation and osteogenic differentiation. As previously described, DNA content in the lysate was quantified spectrofluorometrically using Hoechst 33258 dye (Sigma) and ALP activity was measured using an ALP Assay (Sigma). 28 After the DNA and ALP assays, each homogenized sample was mixed with an equal volume of 1 N acetic acid and placed on a shaker table overnight to dissolve mineral deposits. Calcium content of the cells was then determined using a calcium quantification assay (Diagnostic Chemicals; Charlottetown, PEI, Canada) by the atomic absorption method. 29
A Varioskan multi-mode detection reader (Thermo Electron, Waltham, MA) was utilized for all the absorbance/fluorescence measurements. Three dishes of cells from each donor were analyzed for both groups, and all measurements were conducted in triplicate. The amounts of ALP and Ca2+ produced by each sample were divided by the total cell number of that sample, which was derived from the DNA measurement, thereby allowing statistical comparisons to be made between the two groups.
Scaffold preparation
The β-TCP powder was synthesized by a chemical co-precipitation method. Briefly, reagent-grade calcium nitrate dihydrate [Ca(NO3)2·2H2O] and diammonium hydrogen phosphate [(NH4)2HPO4] were dissolved in distilled water, respectively. To reach the Ca/P molar ratio of 1.5, the (NH4)2HPO4 solution (0.4 M) was added dropwise into the Ca(NO3)2 solution (0.6 M). The reaction solution was continuously stirred at room temperature overnight, precipitated for 12 h, and then filtered. The resulting precipitate was washed first with distilled water and subsequently with absolute ethanol repeatedly to remove NO3− and NH4+, dried at 100°C for 24 h, and calcined at 900°C for 2 h.
Porous β-TCP ceramics were fabricated using the polymeric sponge method as previously reported. 20 First, a flowable β-TCP slurry was prepared by mixing β-TCP powders (containing 5% methyl cellulose as the binder) with deionized water, and the weight ratio of liquid to solid component was 2. The slurry was then used to coat the polyurethane foams, which were cut into the cylinder shape (10 mm in length and 5 mm in diameter). After drying in air to improve their rigidity and mechanical strength, the β-TCP-coated foams were subjected to sintering. The sintering schedule was carried out at a heating rate of 1°C/min from room temperature to 600°C to burn out the polyurethane support, then at a heating rate of 3°C/min from 600°C to 1100°C. The products were maintained at 1100°C for 3 h, cooled back to room temperature, and sterilized by 60Co irradiation before use.
Cell seeding and surgical implantation
The β-TCP cylinders were prewetted in DPBS for 12 h and dried with sterilized filter paper before cell seeding. Freshly isolated BMNCs and osteogenically induced and noninduced BMSCs were resuspended in the serum-free basic culture medium at a density of 5×106 cells/mL, and then were slowly injected onto the β-TCP cylinders using a syringe under slight negative pressure (0.3 mL per cylinder). After being incubated for 4 h at 37°C to allow cell attachment, the cell/scaffold composites were surgically implanted into subcutaneous sites of BALB/c athymic mice (6 weeks old, male). Each animal received two cell/scaffold composites under general anesthesia. All the animals enrolled in this research were cared and processed in accordance with protocols approved by the Institutional Animal Care and Use Committee of TongJi University.
After implantation, the left cells in the media were collected and counted. The ratio between the cells that were adherent to the scaffold and the total amount of seeded cells was then calculated to assess the seeding efficacy of different cell types respectively.
In a parallel experiment, to visualize the cell attachment and spatial distribution on the scaffold, BMNCs and osteogenically induced and noninduced BMSCs were prelabeled with 5 μL/mL of Fluorescent 3,3′-dioctadecyloxacarbocyanine perchlorate (DiO; Molecular Probes, Eugene, OR) dye for 15 min before seeding onto the scaffolds. After incubated for 4 h at 37°C, these composites were sectioned in half. Both the outer and central surfaces were subjected to the confocal laser scanning microscopy (CLSM, Leica Microsystem, Germany) examination.
Micro-CT evaluation
Animals were sacrificed at 4, 8, 12, 16, and 20 weeks after implantation, and the implants were harvested and processed for micro-CT (uCT-80; Scanco Medical, Bassersdorf, Switzerland) and histological analysis. The micro-CT images were recorded on a 1024×1024 charge-coupled device detector, with the pixel size set to 20 μm. After scanning, the images were segmented using a nominal threshold value of 220, and a 3D histomorphometric analysis, including the measurements of total volume and mineral density of the implants, was performed automatically.
Histological analysis
After micro-CT scanning, the implants were fixed in 10% buffered formalin, decalcified in 15% formic acid solution from 3 to 7 days, dehydrated through an ethanol series, and embedded in paraffin. For each implant, ten 5-μm-thick sections cut at different levels (200 μm gap level) were stained with hematoxylin and eosin (HE), and the amount of newly formed bone was assessed as a percentage of the bone area versus the total area within the implant sections. 30 Briefly, the area of new bone formation and the total area within the implant sections were quantified from one photomicrograph by semi-automatically drawing a line around the perimeter of these regions with a software tool on the digitized image of the section. The total number of pixels was then determined within the selected area using the Image-Pro Plus 6.0 software system (Media Cybernetics, Silver Spring, MD). The extent of new bone formation was displayed as a percentage of new bone area per total area as defined above within the implant section. For each implant, 10 sections were used for analysis and 4 images from every section were chosen.
Azan-Mallory staining, which can be used to illustrate bone mature degree, was also performed as previously reported. 31
Statistical analysis
All data were collected with n=6 and presented as means±standard deviation (SD). One-way analysis of variance and Student's paired t-test using SPSS 11.0 software were used to determine possible significant differences (p<0.05) between groups.
Results
Characterization of BMSCs
To characterize the BMSC population in vitro, specific cluster differentiation (CD) antigens of undifferentiated cells derived from six donors were examined individually using flow cytometry. The results showed that human BMSCs comprised a single phenotypic population (over 90% homogeneous) at passage 2. As previously reported, they were uniformly positive for CD29, CD73, CD90, CD105, CD166, and HLA-ABC and negative for CD14, CD31, CD34, CD45, CD79a, and HLA-DR. 27
To confirm the multilineage capability of BMSCs, cells at passage 2 were differentiated toward the chondrogenic, osteogenic, and adipogenic lineages under lineage-specific induction conditions. Chondrogenic differentiation in pellet culture for 3 weeks was confirmed by immunohistochemical staining for the presence of collagen type II deposition (Fig. 1A). Osteogenic induction of confluent BMSCs for 2 weeks (without passaging) resulted in the extracellular matrix mineralization and calcified nodule formation, which was detected by von Kossa staining (Fig. 1B). Adipogenic differentiation in monolayer culture for 2 weeks characterized by intracellular lipid droplets accumulation was revealed by Oil Red O staining (Fig. 1C).

Multilineage differentiation of human BMSCs at passage 2.
Effect of osteogenic differentiation on cell proliferation
About (9.88±1.68)×106 BMNCs could be isolated from each 10 mL bone marrow aspirate. For BMSCs cultured in the osteogenic medium, the CFU-f number was 130±45.2 at day 7, and the cells expanded to (29.33±9.71)×106 before seeding. The noninduced cells had 129.5±45.8 CFU-f, and proliferated to (28.98±7.94)×106 after three-passage culture. Compared with the control group, both the clonogenicity of the marrow aspirates (CFU-f numbers) and the monolayer cell proliferation rates were not affected by osteogenic differentiation (p>0.05, Table 1, Fig. 2A, B). However, after induction in the osteogenic medium for three passages (about 20–25 days), a morphological change from elongated fibroblast-shaped cells to cuboidal osteoblast-like cells could be observed (Fig. 2C, D). BMSCs cultured in basic media displayed a distinctly lower ALP activity and Ca2+ content than the osteogenically induced cells except for the ALP activity of Donor 1 (p<0.05, Fig. 2E, F).
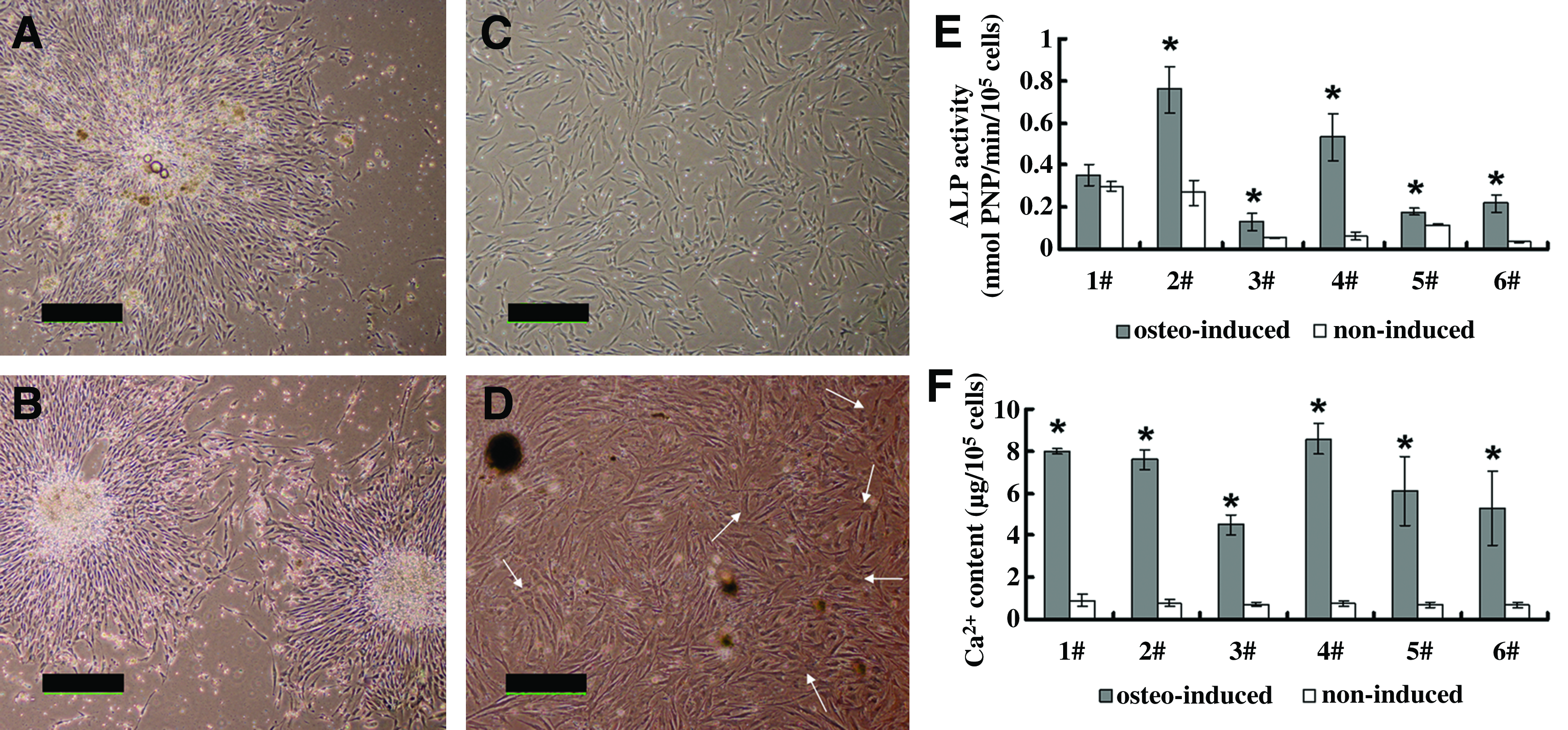
Cell proliferation and osteogenic differentiation in vitro. BMSCs' colonies formed under both undifferentiated
BMSCs, bone marrow mesenchymal stem cells; BMNC, bone marrow mononucleated cells; CFUs, colony-forming units.
Characterization of β-TCP scaffolds
The Ca/P atomic ratio of the synthesized β-TCP powders was determined by inductively coupled plasma analysis, and the result is 1.501. Powder X-ray diffraction (XRD) pattern revealed that except for one small peak of hydroxyapatite, all the other diffraction peak positions match well with the standard XRD pattern of β-TCP (ASTM F1088-04a, 2010), indicating that the crystal phase purity of β-TCP is about 97.3%. The porous β-TCP ceramic cylinders have a volume porosity of 81.7%±3.3%, with pore sizes ranging from 300 to 500 μm and interconnecting pores of 100–200 μm (Fig. 3).
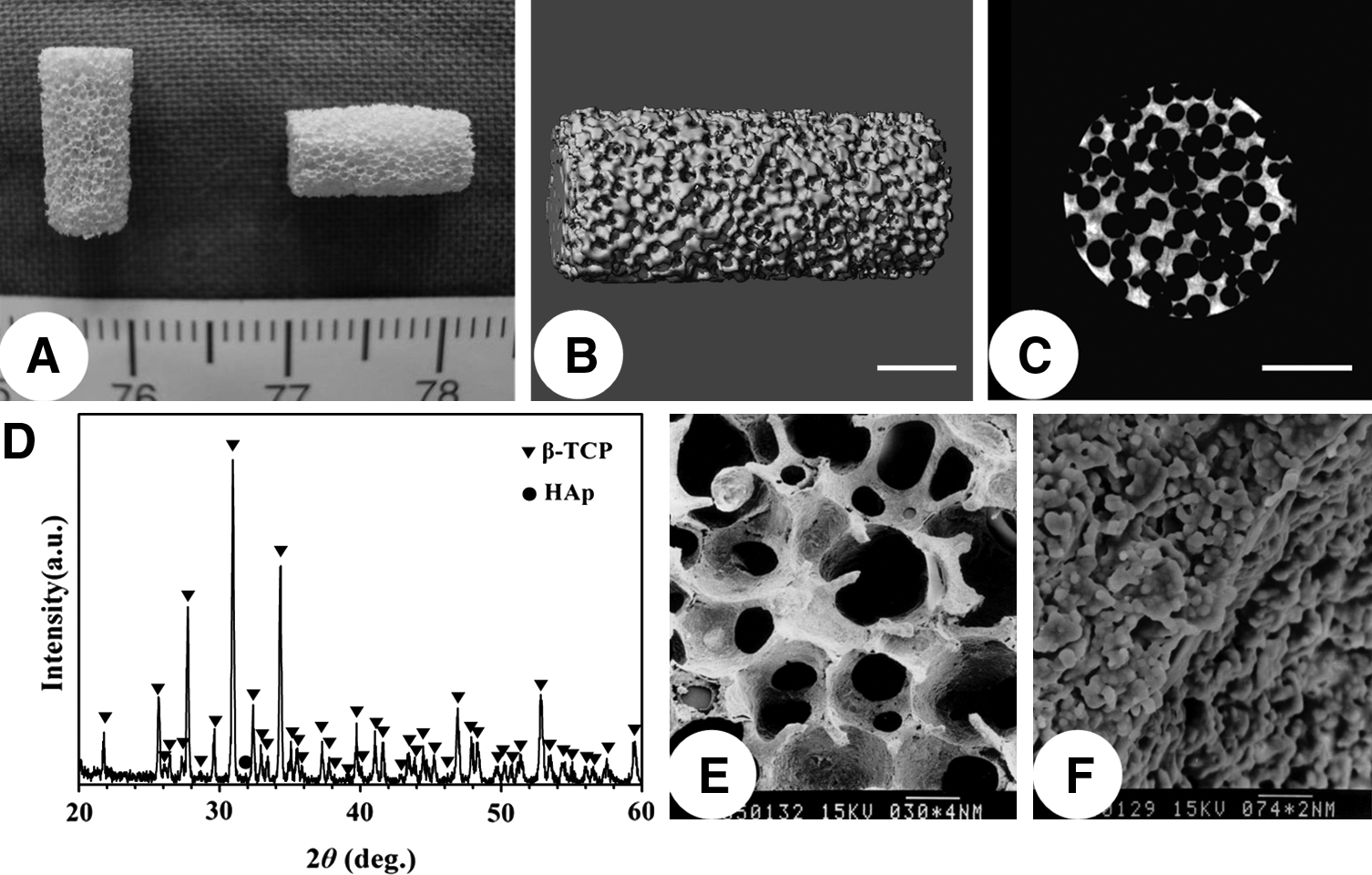
Characterization of β-TCP scaffolds.
Cell seeding efficacy and attachment on the scaffolds
The cell seeding efficacy measurement showed that (89.46±4.42)% of the osteogenically induced BMSCs could adhere to the β-TCP scaffold, and (90.96±3.07)% of the noninduced cells adhered to the material. There was no significant difference in seeding efficacy between these two groups (p=0.717). In contrast, only (27.01±6.47)% of BMNCs remained onto the scaffold 4 h after seeding, exhibiting significantly lower seeding efficacy than BMSCs (p<0.01).
The different cell seeding efficacies were further confirmed by the CLSM observation. As shown in Figure 4, DiO prelabeled BMSCs, no matter whether osteo-induced or not, covered most of the pores on the outer surface, and spread homogeneously inside the inner surface pores. On the contrary, fewer BMNCs adhered to the pore surfaces of scaffold, as evidenced by less green fluorescence.

Fluorescent images of DiO-labeled cells on surfaces of β-TCP ceramics at 4 h postseeding (scale bar: 200 μm). Color images available online at
Micro-CT evaluation
To follow-up the process of new bone regeneration and β-TCP degradation, micro-CT measurements were taken at 4, 8, 12, 16, and 20 weeks postoperatively, and representative images from each group are shown in Figure 5A. In the osteo-induced group, the implant volume remained almost the same from 4 weeks to 12 weeks, and then began to decrease gradually. About 25% of the scaffold volume was degraded at 20 weeks. For the noninduced and BMNC groups, their volumes showed a steadily decreasing trend during the observation period. After 20 weeks, more than 60% and 70% of the original volumes were degraded in these two groups, respectively. Both groups illustrated a distinctly lower volume than that of the predifferentiated group over the observation period except at 4 weeks (Fig. 5B, p<0.05). Quantitative analysis also revealed that the mineral density of the osteo-induced implants peaked at 8 weeks, and then began to decrease gradually, while in the noninduced and BMNCs groups, there was a continuous decrease in implant density from 4 weeks to 20 weeks. The mineral density in the osteo-induced group was significantly higher than that of the other groups at each time point checked except at 4 weeks (Fig. 5C, p<0.05).

Micro-CT measurement of the implants.
Histological analysis
Table 2 summarizes the in vivo histological findings of the cell/scaffold composites at different harvest time points. When β-TCP ceramics combined with osteo-induced BMSCs were implanted subcutaneously into nude mice, tissue-engineered bone formed in the scaffold pores after 4 weeks. The new bone tissue in adjacent pores could be connected through the interconnected pores of the scaffold, thus forming networks of interconnecting trabecular structures. Osteogenesis developed and extended throughout the scaffold at 8 weeks by a process of intramembranous ossification. Almost all of the pores were completely filled with newly formed bone and the regenerated marrow tissue. Evidence of bone remodeling and degeneration was observed at 12 and 16 weeks after implantation, respectively. With continued bone resorption, bone marrow cavities were enlarged, but bone tissue still could be observed in most of the pores at 20 weeks. On the contrary, the pores of the scaffolds seeded with noninduced BMSCs and BMNCs were filled with fibrous-vascular connective tissue, producing no evidence of new bone formation at any harvested times, except for one case. For the noninduced BMSCs of Donor 6, immature bone tissue could be observed in about 10% of all the pores at 8 weeks, in <5% pores at 12 weeks, and then disappeared thereafter. While the BMNCs of the same donor exhibited evidence of new bone formation only at 8 weeks in about 20% pores of the scaffold. With the degradation of the scaffolds in these two groups, the porous structure collapsed after 12 weeks, and the cell-mediated bioresorption event of β-TCP residues surrounded by multinucleated giant cells and connective tissues still could be observed at 20 weeks (Fig. 6).

Histological examination of HE staining.
More detailed cellular events of osteogenesis in the osteo-induced group were illustrated by higher magnification observation of HE staining (Fig. 7A). At 4 weeks after implantation, many spindle-shaped cells became condensed in the scaffold pores and appeared to be producing substantial amounts of bone matrix, which was very immature in structure (pink). The intramembranous ossification progressed vigorously at 8 weeks. Active osteoblasts lined on both the outer and inner surfaces of newly formed woven bone and deposited more extracellular matrices, and the resting osteoblasts had transformed into osteocytes embedded in lacunae of the mineralized bone matrix. The boundary between new matrix (red) and woven bone (light purple) could be clearly distinguished. The vigorous bone formation appeared to become inactive at 12 weeks. Newly regenerated bone containing osteocytes embedded within lacunae was being remodeled, and no osteoblastic layer lining the luminal surface was found. Thick woven bone tissue had been remodeled into thin lamellar bone at 16 weeks, and newly formed marrow composed of hemopoietic cells and adipose tissue occupied the intertrabecular space. At 20 weeks, the amount of bone tissue as well as the number of osteocytes became much fewer and the hematopoietic tissue was more abundant. The extent of osteogenesis was quantitatively illustrated in Figure 7B. The average percentage of new bone area to total area increased from 4 weeks to 8 weeks, peaked at 8 weeks, and then began to decrease gradually from 12 to 20 weeks.

Azan-Mallory staining also confirmed the process of bone formation exhibited by the osteo-induced implants with time. Newly formed woven bone (light blue) associated with the invading vasculature was detected in the pores at 4 weeks, and mature bone tissue (dark red) appeared at 8 weeks. More extensive and remodeled bone was observed at 12 weeks, and this phase of bone remodeling apparently subsided at 16 weeks. At 20 weeks, the newly formed marrow cavities had occupied the central part of the pores, and the amount and mature degree of bone tissue decreased dramatically (Fig. 8).

Azan-Mallory staining also illustrated the different developing stages of osteogenesis in the osteo-induced group with time. Newly formed bone appeared blue at early stage, and became red and dark after maturation (scale bar: 100 μm). Color images available online at
Discussion
As one possible approach to treat bone defects in orthopedic surgery, the composite grafting of calcium phosphate ceramic and BMSCs with osteogenic potential has been used as a bone substitute recently. However, despite the promising results reported in large-animal studies and clinical cases,6,8,11,12 these BMSC-based osteogenic grafts are yet to be included in the routine toolkit of surgical practice. 32 One of the major possible reasons lies in the lack of standardized procedures for the production of these engineered tissues.
This study investigated a relatively easy method to engineer bone tissue in vivo, which may contribute to the standardization and reproducibility in the production and performance of tissue-engineered bone. When porous β-TCP ceramics combined with preosteogenically differentiated BMSCs were implanted subcutaneously into nude mice, massive de novo bone formed in the pore regions of the ceramics as early as 4 weeks and maintained for 20 weeks postimplantation. On the contrary, except for one case, the noninduced culture-expanded BMSCs showed only fibrovascular tissue in the porous scaffolds. These composites were negative for bone formation no matter how long they were kept in vivo. For that exception, the noninduced BMSCs initiated bone formation much later and produced much less bone than the induced cells did, indicating that the use of predifferentiated osteoblasts enhanced the rate and extent of osteogenesis.
Interestingly, four distinctly different stages took place during the sequence of heterotopic bone formation of osteo-induced BMSCs, that is, bone formation, maturation, remodeling, and resorption. Woven bone formed at 4 weeks after implantation, progressed and matured at 8 weeks, and showed remodeling (lamellar bone) at 12 weeks. From 4 to 12 weeks, the volume of implants without bone formation shrunk over 30%–50%, while the implants containing bone regeneration almost remained their original volumes (Fig. 5B), indicating that the degradation rate of β-TCP scaffolds can well match the speed of new bone formation. From 16 to 20 weeks, bone degeneration occurred, and the implant volume also decreased accordingly. Since only donor cells contribute to heterotopic bone formation,33,34 it seems conceivable that this bone resorption phenomenon is caused by the implantation site. When lacking mechanical stimulation and muscle tension, tissue-engineered bone tends to be unstable and is often reabsorbed at subcutaneous unloaded sites. 19
It is known that the osteogenicity of implanted cell/scaffold composites requires a certain number of BMSCs, as the natural process of bone regeneration does, but the critical cell density required for ectopic osteogenesis is still debated. Despite the positive histological findings of bone formation in vivo in many studies, the cell seeding densities varied between different laboratories and different cell sources.13–22,32 We found that the minimal concentration of cell suspension required for ectopic osteogenicity of human BMSC/β-TCP composites was 2×106 cells/mL (unpublished data). So the initial cell density used for seeding ceramics in this study was determined as 5×106 cells/mL, and the results of predifferentiated BMSCs group showed reproducible, abundant, and uniform bone tissue formation after transplantation in nude mice.
The potential of ectopic bone regeneration by freshly isolated BMNCs was also evaluated in this study. Human BMNCs represent a heterogeneous population of cells, including stromal cells, fat cells, macrophages, endothelial cells, HSCs, and BMSCs. Since most of these populations are nonadherent, only (27.01±6.47)% of BMNCs could be seeded onto the scaffolds, much lower than that of the culture-expanded BMSCs, as evidenced by measurements of the cell seeding efficacy and observation of the CLSM. So after isolating BMNCs with the density gradient centrifugation method, BMSCs are further selected on the basis of adhesion and proliferation on the plastic substrate. Nonadherent cells decrease during progressive passages, and the adherent population becomes homogenous at passage 2, illustrating the typical MSC immunophenotypes and multilineage differentiation potential.
Based on data of the nondifferentiated cells, each 10-mL marrow aspirate yielded about (9.88±1.68)×106 BMNCs, and produced (130±45.2) CFU-f after 7 days in culture. So the initial number of BMSCs represents only (0.0013±0.0002)% of the mononucleated cells (calculated as the percentage of CFU-f among the corresponding BMNCs), consistent with the results of some previous reports.32,35–37 Although the cell seeding density was the same in three groups, an insufficient number of osteo-progenitor cells was initially seeded per scaffold in the BMNCs group, and produced limited and delayed bone formation only in one case.
Gan et al. used freshly enriched autologous BMSCs combined with β-TCP ceramics as bone grafts to treat posterior spinal fusion, and 95.1% cases achieved good spinal fusion results. 23 They, however, reported that an average of 252 mL of bone marrow aspirate per patient is needed to enrich sufficient BMSCs. Since the amount of BMSCs and their differentiation potential decrease with aging or disease,19,38,39 it frequently is impractical to obtain such large amounts of bone marrow with the requisite numbers of BMSCs, and this approach may be least applicable in those situations where it is most needed. Therefore, expanding and preinducing a relatively small amount of BMSCs into an appropriate number of osteo-progenitor cells will be of great clinical benefit.
β-TCP is well known as a biodegradable material with good osteoconductive capacity and demonstrates efficiency as scaffolds for bone tissue engineering.40,41 Fabricated using the polymeric sponge method, the β-TCP ceramics have a macroporous structure, which can guarantee the uniform cell distribution and blood vessel ingrowth, resulting in the extensive bone formation throughout the scaffold. Our previous study demonstrated that β-TCP scaffolds alone exhibit no osteoinductive properties and result in no de novo bone deposition at subcutaneous sites in nude mice 20 ; thus, this control group of implanting cell-free scaffolds was not included in the present work. As to the β-TCP biodegradability, we did not analyze the scaffold degradation rate because of demineralization treatment in this study. When samples are stained using the undecalcified hard-tissue sectioning techniques, it is possible to distinguish the newly formed bone from the undegraded scaffolds accurately. 42 For decalcified samples, most β-TCP scaffolds were dissolved during the demineralization procedure. Only corresponding blank areas with some remnant ghost of β-TCP were left within the implant sections (Fig. 6C). Thus, the comparison of the volume of new bone formation versus remains of scaffolds was limited in this study.
Current osteoinduction conditions include culturing cells in the presence of dexamethasone, L-ascorbic acid, and β-GP.27,35 To determine the effect of osteogenic condition on the clonogenicity and proliferation rate of BMSCs, human BMNCs were cultured in the presence or absence of these supplements after isolation. The results showed that osteo-induction changed neither the colony-forming efficiency (total number of colonies formed per 10 mL marrow aspirates) nor the total cell number at the end of the culture period. Compared with the noninduced cells, in vitro osteogenic differentiation of BMSCs was confirmed by higher levels of ALP activity and calcium deposition, indicating that delivering a mature osteoblastic population is capable of more rapid and extensive bone formation in vivo. Future studies are required to investigate the correlation of these osteogenic markers and subsequent ectopic bone formation and to establish which marker in vitro may be used to predict in vivo osteogenesis.
One limitation of this study is the usage of FBS in the culture medium. Because of the possible cross contamination from the animal product, animal serum used in human cell culture is not recommended by regulatory agencies around the world now. Therefore, further studies of investigating an animal product-free culture medium for producing the clinical-grade stem cells are urgently needed.
Another limitation of this work is that the present animal model provides no osteoinductive growth factors and mechanical loading conditions. Since undifferentiated BMSCs can be driven into osteogenic lineage solely by the local microenvironment of bone, 25 this study requires to be further evaluated in an orthotopic site, such as a critical-sized bone defect model.
Conclusion
In the present study, the osteogenicity of human BMNCs and in vitro expanded undifferentiated and osteogenically differentiated BMSCs was compared in an athymic mouse model. The results demonstrated that in vitro osteogenic induction of human BMSCs before seeding β-TCP ceramics significantly enhanced subsequent ectopic bone formation compared to samples implanted with undifferentiated BMSCs and fresh BMNCs. Therefore, it may be possible to obtain a relatively small amount of autologous BMSCs from patients, expand and predifferentiate them into an appropriate number of osteo-progenitor cells, incorporate these cells with porous ceramics, and graft the composites into the donors to facilitate the repair of bone defects.
Footnotes
Acknowledgments
The authors gratefully thank Dr. Li Zhao (National Tissue Engineering Research Center of China) for providing graciously the β-TCP scaffolds. We acknowledge Dr. James D. Kretlow (Division of Plastic Surgery, Baylor College of Medicine) for his insightful comments and polishing the article. This work was financially supported by the Natural Science Foundation of China (30800232 and 81171475), and Shanghai Science and Technology Committee Project (10QH1402400).
Disclosure Statement
No competing financial interests exist.