
Abstract
Despite the improvements in the human embryonic stem cell (hESC) culture systems, very similar conditions to those used to maintain hESCs on mouse feeders are broadly applied to culture methods based on human feeders. Indeed, basic fibroblast growth factor (bFGF), a master hESC-sustaining factor, is still added in nearly all medium formulations for hESC propagation. Human foreskin fibroblasts (HFFs) and mesenchymal stem cells (MSCs) used as feeders have recently been reported to support hESC growth without exogenous bFGF. However, whether hESCs may be maintained undifferentiated without exogenous bFGF using media conditioned (CM) by human feeders remains elusive. We hypothesize that HFFs and hMSCs are likely to be functionally different and therefore the mechanisms by which HFF-CM and MSC-CM support undifferentiated growth of hESCs may differ. We have thus determined whether HFF-CM and/or MSC-CM sustain feeder-free undifferentiated growth of hESC without exogenous supplementation of bFGF. We report that hMSCs synthesize higher levels of endogenous bFGF than HFFs. Accordingly and in contrast to HFF-CM, MSC-CM produced without the addition of exogenous bFGF supports hESC pluripotency and culture homeostasis beyond 20 passages without the need of bFGF supplementation. hESCs maintained without exogenous bFGF in MSC-CM retained hESC morphology and expression of pluripotency surface markers and transcription factors, formed teratomas, and showed spontaneous and lineage-directed in vitro differentiation capacity. Our data indicate that MSC-CM, but not HFF-CM, provides microenvironment cues supporting feeder-free long-term maintenance of pluripotent hESCs and obviates the requirement of exogenous bFGF at any time.
Introduction
bFGF appears to be a key player in sustaining hESC pluripotency and it is added in nearly all available medium formulations for hESC procurement and expansion. In fact, despite the evolution of hESC culture systems, identical conditions used to maintain hESCs on MEFs have also been applied in expansion protocols using human feeders, with exogenous supplementation of bFGF still being required to sustain hESC undifferentiated growth. Typically, both feeders used for CM production and hESCs themselves during undifferentiated growth are supplemented with bFGF.10,11,15 Recently, Park et al.9,16 reported that several human feeders, including HPCs, HFFs, and MSCs, support long-term maintenance of hESCs without exogenous bFGF supplementation.9,16 However, it remains to be addressed whether hESCs can be maintained undifferentiated without exogenous bFGF supplementation in feeder-free systems using CM by human feeders. The development of feeder-free and exogenous bFGF-free hESC culture systems would highly facilitate hESC culture conditions by reducing exogenous components and would make hESC maintenance more affordable. Importantly, less complex hESC culture systems (as few exogenous components as possible) should also facilitate the study of key developmental signaling pathways contributing to preserve the balance between self-renewal and differentiation.
Here, we hypothesize that HFFs and MSCs are likely to be functionally different providing distinct microenvironment cues (cellular interactions and secreted factors) to hESCs. Consequently, the mechanisms by which HFF-CM and MSC-CM support undifferentiated growth of hESCs might also differ. To facilitate hESC culture conditions and to reduce exogenous compounds, we were prompted to analyze whether HFFs and/or MSCs secrete bFGF during CM production as this bFGF may partially contribute to preserving undifferentiated growth of hESC in feeder-free conditions without exogenous supplementation of bFGF during CM production, hESC growth or both. We found that hMSCs secrete significantly higher levels of bFGF than HFFs. Accordingly, in contrast to HFF-CM, MSC-CM produced in the absence of bFGF supports hESC pluripotency and culture homeostasis beyond 20 passages without the need of exogenous bFGF supplementation. hESCs maintained without bFGF supplementation in MSC-CM retained typical undifferentiated morphology, expression of pluripotency surface markers and transcription factors, and showed in vitro (spontaneous and directed) and in vivo differentiation capacity into tissues representing the three germ layers. Our data indicate that the MSC-CM provides specific microenvironment conditions supporting long-term feeder-free maintenance of undifferentiated hESCs without the requirement of exogenous bFGF.
Materials and Methods
hESC culture and preparation of CM
H9 and AND1 hESC lines were grown feeder-free for 20 passages on matrigel flasks (BD Biosciences) in MSC-CM or HFF-CM with or without bFGF supplementation (8 ng/mL). The medium was changed daily; the hESC cultures were also visualized daily by phase-contrast microscopy and split every 6–7 days with collagenase IV. The basal medium used to prepare CM was 80% Knockout-Dulbecco's modified Eagle's medium (DMEM), 20% KO Serum Replacement, 1% NEAA, 1 mM
Reverse transcriptase-polymerase chain reaction for ESC factors and bFGF
Total RNA was extracted from hESCs, MSCs, and HFFs using Trizol (Invitrogen) followed by DNase treatment (Invitrogen). cDNA synthesis was done with 0.5 μg of total RNA using the First-Strand cDNA Synthesis Kit (Amersham). The cDNA was used for end-point and quantitative polymerase chain reaction (qPCR). The expression of the ESC factors Nanog, Oct3/4, Rex-1, and Sox-2 was assessed by end-point reverse transcriptase polymerase chain reaction (RT-PCR). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as normalizer. The primers used were Nanog: 5′-TGCAGTTCCAGCCAAATTCTC-3′ and 5′-CCTAGTGGTCTGCTGTATTACATTAAGG-3′; Oct3/4: 5′-TCTGCAGAAAGAACTCGAGCAA-3′ and 5′-AGATGGT CGTTTGGCTGAACAC-3′; Rex-1: 5′-CAGATCCTAAACA GCTCGCAGAAT-3′ and 5′-GCGTACGCAAATTAAAGTC CAGA-3′; Sox-2: 5′-CCCCCGGCGGCAATAGCA-3′ and 5′-TCGGCGCCGGGGAGATACAT-3′; and GAPDH: 5′-GAA GGTGAAGGTCGGAGTC-3′ and 5′-GAAGATGGTGATGG GATTTC-3′. The conditions used for end-point PCR were as follows: 5 min at 94°C, 35 cycles of 30 s at 94°C followed by 50 s at 60°C and 50 s at 72°C, and a final extension of 10 min at 72°C.
For the bFGF qPCR, two sets of different primers were used: primer set A (5′-GTGTGTGCTAACCGTTACCT-3′ and 5′-GCTCTTAGCAGACATTGGAAG-3′) and primer set B (5′-CTTCCTGCGCATCCACCCCG-3′ and 5′-AGCCAGG TAACGGTTAGCACACA-3′). qPCR conditions were as follows: 3 min at 95°C, and 40 cycles of 10 s at 95°C followed by 20 s at 60°C. FGF2 mRNA expression was normalized against GAPDH. Real-time PCR was performed using the Mx3005 Real-Time PCR system (Stratagene) and the SYBR Green QPCR Master Mix (Agilent Technologies).
Enzyme-linked immunosorbent assay detection of bFGF in CM
MSCs and HFFs were grown in Ad-DMEM supplemented with 10% FCS and 2 mM
Flow cytometry characterization of hESCs
Flow cytometry analysis was carried out as previously described.15,21 For cell surface markers, hESCs were dissociated with 0.5% trypsin and the single-cell suspension was stained (2–5×105cells/mL) with TRA-1-81-FITC, TRA-1-60-PE, SSEA-4-FITC, and SSEA-3-PE (BD). For intracellular staining, hESCs were fixed and permeabilized using the Fix&Perm reagent (Caltag Laboratories)22,23 before OCT-4 staining (FITC-conjugated MoAb from BD). The cells were then washed and stained with 7-AAD (Immunotech) for 15 min on ice. As negative control, an isotype-matched nonspecific mouse Ig was analyzed. Cells were analyzed using a FACSCanto flow cytometer. Supplementary Figure S1 (Supplementary Data are available online at
Embryoid body development
hESCs were transferred to low-attachment plates and permitted to differentiate spontaneously through embryoid body (EB) formation in KO-DMEM+20% FCS+1%
Hematopoietic differentiation from hESCs
Confluent hESCs maintained in MSC-CM with or without bFGF supplementation were treated with collagenase IV, scraped off the Matrigel, and transferred to low-attachment plates (Corning) to allow human EB (hEB) formation by overnight incubation in the differentiation medium. The medium was changed the next day with an identical differentiation medium supplemented with hematopoietic growth factors.25–27 hEBs were dissociated at days 10, 15, and 22 of development, and the single-cell suspension achieved by gentle pipetting was stained with anti-CD34-FITC, anti-CD31-PE, anti-CD45-APC (Miltenyi Biotyec), and 7-AAD. Live cells (7-AAD-) were analyzed using a FACS-Canto flow cytometer. Hemogenic precursors were identified as CD31+ CD34+CD45−, blood cells as CD45+, and primitive blood cells as CD45+CD34+ (Fig. 5C).25,27–29 Human clonogenic progenitor assays colony forming units (CFUs) were carried out by plating 50,000 cells from day 15 hEBs into methylcellulose H4230 (Stem Cell Technologies) supplemented with 50 ng/mL SCF, 3 units/mL erythopietin, 10 ng/mL granulo-monocyte colony stimulating factor, and 10 ng/mL interleukin-3. Colonies were scored at day 14 of the CFU assay using standard criteria.26,27,30
In vivo teratoma formation
In vivo protocols were approved by the University of Granada Council on Animal Care and Experimentation. In vivo pluripotency was assessed as previously described. 31 Briefly, hESCs were harvested and innoculated beneath the testicular capsule of 8-week-old NSG male mice (The Jackson Lab). Teratoma growth was checked by palpation every week. Mice were sacrificed within a week after tumors were clearly palpable or ∼12 weeks after implantation, if injected cells failed to form teratoma. Tumors were fixed and embedded in paraffin, and sections were stained with hematoxylin and eosin. 31
Results
MSCs express and secrete higher levels of bFGF than HFFs
Many attempts have been conducted to develop culture systems based on human feeders7,8,32–37 and human feeder-CM such as HFF-CM14,24,25 and MSC-CM.15,27 Importantly, human feeders have very recently been reported for the first time to sustain undifferentiated hESCs without exogenous bFGF supplementation. However, whether hESCs can be equally propagated without exogenous bFGF using HFF-CM and/or MSC-CM remains to be addressed. Based on this, we were prompted to analyze whether HFFs and hMSCs used to produce CM express and release bFGF, a master hESC-supportive factor. Human MSCs were found to express more than threefold higher levels of bFGF mRNA than HFFs (Fig. 1A). Accordingly, MSCs secreted more than threefold higher levels of bFGF than HFFs (8.5±0.5 vs. 2.7±0.4 pg/mL), as detected by ELISA (Fig. 1B). This observation suggests that HFFs and hMSCs may be functionally distinct, providing differential microenvironment cues to hESCs and therefore the mechanisms by which HFF-CM and MSC-CM support undifferentiated growth of hESCs might also differ.

Mesenchymal stem cells (MSCs) synthesize higher levels of basic fibroblast growth factor (bFGF) than human foreskin fibroblasts (HFFs).
bFGF supplementation solely during CM production suffices for maintenance of undifferentiated hESCs in MSC-CM, but not in HFF-CM
Typically, both feeders during the CM production and hESCs themselves during undifferentiated growth are supplemented with bFGF.10,38,39 Here, an initial experimental strategy was designed whereby both MSC-CM and HFF-CM were produced in the presence of bFGF and used to feed, over a 20-week period, two hESC lines (H9 and AND1) in the presence (+/+) or absence (+/−) of exogenous bFGF (Fig. 2A). In line with our previous observations, 15 both hESC lines were successfully maintained pluripotent using both MSC-CM and HFF-CM (produced in the presence of bFGF) upon supplementation of the CM with exogenous bFGF during hESC growth (+/+ conditions) (Figs. 2 and 3). However, when hESCs were maintained without supplementation of exogenous bFGF (+/− conditions) significant differences were found in the potential of MSC-CM and HFF-CM to support undifferentiated hESCs. Noticeably, bFGF supplementation solely during CM production (+/− conditions) sufficed for long-term feeder-free maintenance of undifferentiated pluripotent hESCs in MSC-CM, but not in HFF-CM (Figs. 2 and 3). H9 and AND1 hESC lines maintained for 20 passages in MSC-CM in the absence of exogenous bFGF (+/−) retained typical hESC morphology and continued to express identical levels of the ESC-transcription factors Nanog, Oct4, Rex-1, and Sox2 (Fig. 2B) and the ESC surface markers SSEA-4, SSEA-3, Tra-1-60, and Tra-1-81 (Fig. 2C). The gold-standard pluripotency assay is the ability to form in vivo teratomas upon injection into immune-deficient mice. Both H9 and AND1 lines grown in MSC-CM +/− formed teratomas with an efficiency (84%) and latency (51±30 days) similar to those reported for the same hESC lines maintained in MSC-CM +/+ (Fig. 3A). These teratomas contained tissues representing the three germ layers (Fig. 3B), thus demonstrating the pluripotency of hESC lines maintained in MSC-CM +/−. In contrast, hESC lines maintained in HFF-CM in the absence of exogenous bFGF (+/−) displayed reduced expression levels of the ESC transcription factors Nanog, Sox2, and Rex1 (Fig. 2B) and the ESC surface markers SSEA-4, SSEA-3, Tra-1-60, and Tra-1-81 (Fig. 2C). More importantly, these hESCs expanded in HFF-CM in the absence of exogenous bFGF (+/−) failed to form teratomas after a 12-week period, indicative of loss of pluripotency potential (Fig. 3). Taken together, these data indicate that hESCs can be successfully maintained undifferentiated without exogenous supplementation of bFGF in MSC-CM, but not in HFF-CM, both produced in the presence of bFGF.
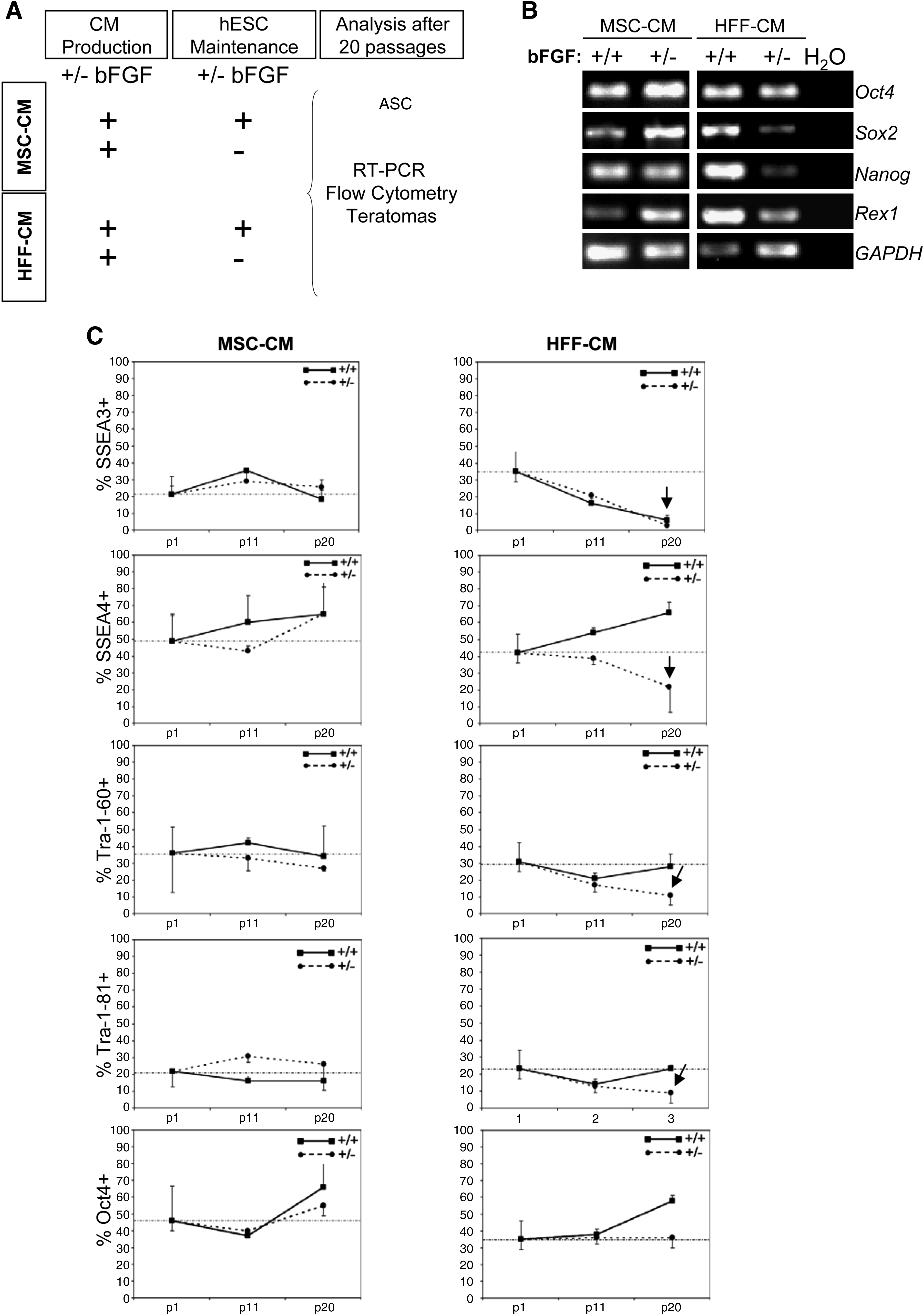
bFGF supplementation exclusively during conditioned media (CM) production suffices for long-term feeder-free maintenance of undifferentiated human embryonic stem cells (hESCs) in MSC-CM, but not in HFF-CM.
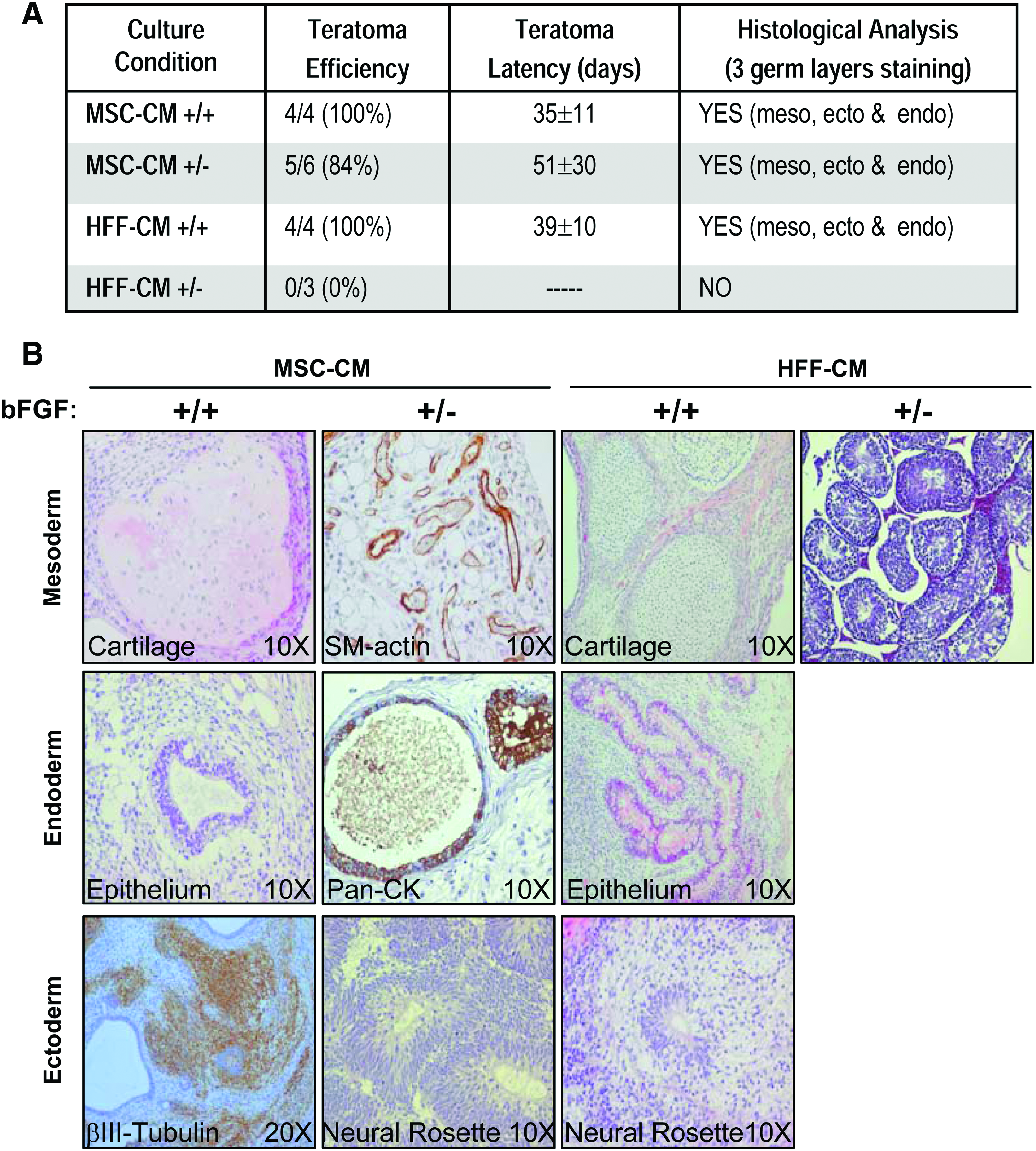
hESCs maintained without bFGF supplementation in MSC-CM, produced in the presence of bFGF, retain the capacity to form teratomas.
Maintenance of hESCs in MSC-CM does not require bFGF supplementation either during CM production or during hESC growth
We next wanted to address to what extent bFGF added to MSC cultures during CM production may be responsible for the ability of MSC-CM to support hESC growth without exogenous bFGF supplementation. It is plausible that (i) the bFGF used to produce MSC-CM remains bioactive on hESCs during propagation or that (ii) additional hESC-supportive factors are produced by MSCs in response to bFGF during CM preparation. Additionally, the ability of MSC-CM to support hESC growth without exogenous bFGF supplementation may be attributed either to the levels of bFGF synthesized by the MSCs themselves or to other MSC-synthesized soluble hESC-supportive factors.
To study this, MSC-CM produced without the addition of bFGF was used to feed, over a 20-week period, H9 and AND1 hESCs in the presence (−/+) or absence (−/−) of exogenous bFGF (Fig. 4A). Strikingly, hESCs were successfully maintained undifferentiated in MSC-CM produced in the absence of bFGF without further supplementation of bFGF during hESC growth (−/−). hESCs grown in MSC-CM without bFGF supplementation during both CM production and hESC growth (−/−) retained after 20 passages the expression of the ESC factors Nanog, Oct4, Sox2, and Rex1 (Fig. 4B) and the hESC surface markers SSEA-4, SSEA-3, Tra-1-60, Tra-1-81, and Oct-4 (Fig. 4C). Similarly, hESCs maintained in MSC-CM −/− formed teratomas containing a variety of tissues representing the three germ layers with 100% efficiency and latency (56 days vs. 49 days) very similar to those hESCs grown in MSC-CM −/+ (Fig. 5A), indicating that hESC lines propagated in MSC-CM −/− retain pluripotency. These data indicate that prolonged propagation of hESCs feeder-free in MSC-CM completely obviates the requirement of exogenous bFGF supplementation.

Maintenance of hESCs in MSC-CM does not require bFGF supplementation either during CM production or during hESC growth.
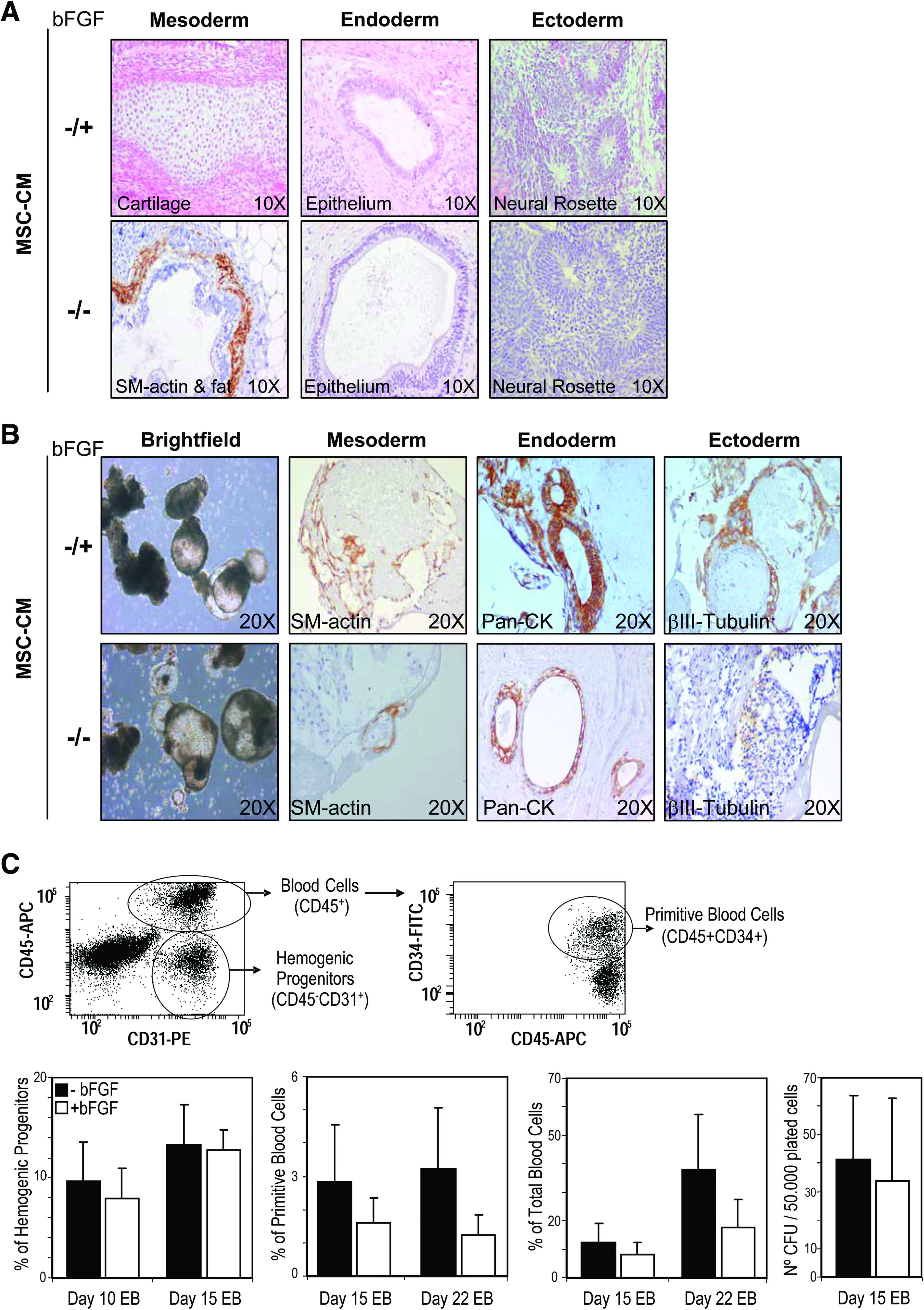
hESCs maintained in MSC-CM without any bFGF supplementation retain the capacity to undergo spontaneous and directed differentiation in vitro and in vivo.
hESCs maintained in MSC-CM without bFGF retain in vitro spontaneous and lineage-directed differentiation potential
Improvements and standardization of hESC culture conditions must warrant that the differentiation potential of the hESCs is not compromised. We thus analyzed whether the removal of bFGF negatively affects the spontaneous or directed in vitro differentiation potential of hESCs maintained in MSC-CM. hESCs maintained in MSC-CM either with or without bFGF showed identical in vitro EB spontaneous differentiation into tissues representing the three germ layers: mesoderm (SM-actin+ cells), endoderm (Pan-CK+ cells), and ectoderm (β-III-Tubulin+ cells) (Fig. 5B). To further determine whether the absence of bFGF during hESC propagation in MSC-CM compromises lineage-directed differentiation, both H9 and AND1 hESC lines were induced to differentiate toward hematopoietic lineage through the formation of hEBs in the presence of the BMP4 and hematopoietic growth factros.25–27 We analyzed the emergence of hemogenic progenitors (CD45-CD31+), primitive (CD45+CD34+) and mature (CD45+CD34−) blood cells, and CFU potential at different time points during differentiating hEBs (days 10, 15, and 22) (Fig. 5C). As it can be shown in Figure 5C, hESCs maintained in MSC-CM without bFGF supplementation displayed similar or higher hematopoietic potential than those grown in the presence of bFGF (Fig. 5C), highlighting that the removal of exogenous bFGF in hESC cultures maintained in MSC-CM does not seem to compromise the differentiation potential of hESCs.
Discussion
Our data indicate that MSC-CM, but not HFF-CM, provides microenvironment conditions supporting feeder-free long-term maintenance of undifferentiated hESCs and obviates the requirement of exogenous bFGF both during CM production and during hESC growth. To the best of our knowledge, this is the first time that hESCs are successfully maintained undifferentiated without losing self-renewal or pluripotency features for prolonged periods of time in a feeder-free culture system without exogenous bFGF supplementation. hESCs maintained in MSC-CM without bFGF supplementation retained fully spontaneous and hematopoietic differentiation potential, demonstrating that the removal of exogenous bFGF in hESC cultures maintained in MSC-CM does not impair the potential of hESCs to differentiate toward certain lineages. This further validates our feeder-free, bFGF-free culture system because improvements and standardization of hESC culture conditions must warrant that the differentiation capacity of the hESCs is not compromised.
Importantly, hESCs are equally propagated and display very similar differentiation performance in MSC-CM regardless of the supplementation of exogenous bFGF. This demonstrates that neither carryover/residual bFGF added to MSCs during MSC-CM production remains bioactive on hESCs during propagation, nor additional hESC-supportive factors are produced by MSCs in response to bFGF during MSC-CM preparation. bFGF represents a master human pluripotent stem cell (hPSC)-sustaining factor that is included in nearly all medium formulations for hPSC propagation. Despite the significantly higher levels of bFGF secreted by MSCs as compared to HFFs, it is still tempting to speculate that the ability of MSC-CM to support hESC growth without exogenous bFGF supplementation may be attributed not exclusively to the levels of bFGF synthesized by the MSCs, but also to additional soluble, MSC-synthesized hESC-supportive factors. Optimization of hESC-supportive media is a daunting challenge. For instance, it has been reported that TeSR, a chemically defined medium for hESC culture, is composed of at least 70 components. 12 Because of the combinatorial complexity, future prospective proteomic studies should aim at deciphering the MSC secretome. This should provide very valuable information about the specific components (and their concentrations) sustaining hESC performance in the absence of bFGF.
Apart from the differences in bFGF production, HFFs and MSCs are expected to provide different feeder cell-hESC interactions and secretome to hESC, which may, in turn, activate or inhibit a combination of signaling pathways and mechanisms important for the regulation of hESC homeostasis and pluripotency. For instance, in the absence of exogenous bFGF, HFFs used as feeders seem to perform different from HFF-CM. Thus, Park et al.9,16 recently reported that HFFs used as feeders permit the maintenance of hESCs without extrinsic bFGF, while our data indicate that in the absence of bFGF HFF-CM is unable to sustain long-term feeder-free propagation of hESCs. This suggests that when HFFs feeders are used for hESC growth, mechanisms activated through HFFs-hESC physical contact (and not HFF-secreted factors) seem responsible for hESC maintenance. In striking contrast with HFFs, we have shown here that in the absence of exogenous bFGF, MSC-CM performs equally to MSCs used as feeders, as reported by Park et al.9,16 and Cortes et al. 21 This suggests that when MSCs are used for hESC culture, pluripotency-regulating mechanisms activated by MSC-released soluble factors may be more relevant for hESC growth than those relying on MSCs-hESC interaction.
Footnotes
Acknowledgments
This work was funded by the CICE-FEDER (P08-CTS-3678) de la Junta de Andalucía to P.M., the FIS/FEDER (PI10/00449) to P.M. and (PI11/00119) to C.B. the MICINN (PLE-2009-0111) to P.M., and the Fundación AECC-Junta Provincial de Albacete (CI-2011) to P.M. C.B. and P.J.R. were supported by the Instituto de Salud Carlos III (CP07/00059 and CP09/0063, respectively). V.R.-M. was supported by a Marie Curie fellowship (PIIF-GA-2009-236430). I.G.-A. was supported by the MICINN (PTA-2009-01804-I).
Disclosure Statement
Authors disclose neither potential conflict of interest nor competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.