
Abstract
Currently, stem cell research faces a major bottleneck related to the low efficiency of methods to produce large quantities of human embryonic stem cells (ESC) for use in clinical trials. Most culture media currently employed for human ESC cultivation contain animal compounds, and cells are grown in static flasks. Besides the immediate contamination with nonhuman compounds, cell expansion in flasks tends to be laborious and nonefficient. Here we cultured human ESC in stirred microcarrier (MC) systems using an animal/human-component-free medium, to overcome both issues. The method developed to culture cells on suspended beads combined the use of polymeric MCs in stirred vessels with an optimized culture medium free of supplements of animal and human origin. This approach generated approximately 160 million cells within 6 days, which were shown to remain pluripotent. The process developed herein provides a step forward toward therapy due to the economic advantages in the production of human ESC and to their consequent low immunogenic potential.
Introduction
Despite more than a decade has elapsed since the first human ESC derivation, 2 expansion of these cells is still performed in a heterogeneous manner using conventional static flasks. Use of such culture techniques limits the yield and reduces the overall quality of expanded cells. Suspension culture using microcarriers (MCs) can, therefore, be an attractive alternative for growing both murine13–17 and human18,19 stem cells. MC beads provide high surface–volume ratios and enable the use of homogeneous stirred culture systems, which result in cell concentrations two to fourfold higher than those obtained with static plates.18,19
Although improvements in microcarrier-based cell culture have been described for ESC, the published reports employ media and/or matrices containing animal-derived compounds,17–21 making the resulting cultured ESC unsuitable for cell therapy.
Indeed, the great majority of ESC works use knockout™ serum replacement (KSR) as a medium supplement, 22 which is a serum substitute comprised of salts, aminoacids, and animal proteins. 23 Because human ESC can incorporate the nonhuman sugar Neu5Gc, 24 cells expanded in such media are prone to trigger a strong immune response in patients. 25 Using new media that do not contain animal compounds are highly desirable.
Even though some groups were able to develop ESC culture protocols in which cells are grown as floating aggregates,26,27 these protocols showed low efficiency and reproducibility 28 and/or relied on expensive reagents, such as ROCK-inhibitors 29 or on high concentrations of growth factors. 30 Therefore, MCs seem to be the best alternative available for culturing ESC in homogeneous, scalable stirred culture systems.
In the present study, a xeno-free process to culture human ESC in stirred MC systems was developed, using a novel, chemically defined, animal/human-component-free medium—the BRASTEM medium. We describe, for the first time, the culture of Neu5Gc-free ESC on MCs with the BRASTEM medium, a highly efficient and scalable platform.
Materials and Methods
Cell line and culture media
Undifferentiated H9 human ESC (WiCell Research Institute) were routinely cultured at 37°C and 5% CO2 in 100-mm tissue culture plates (TPP) on inactivated mouse embryonic fibroblasts (MEFs) (Hygeia Biotech). The KSR medium consisted of the high-glucose Dulbecco's Modified Eagle Medium (DMEM)/F12 supplemented with additional 2 mM of
Two other culture media evaluated in stirred MC cultures were xeno-free. The first medium was identical to the one described above, but employing knockout serum replacement xeno-free (Life Technologies), and was called KSR-XF. The second xeno-free medium, hereby entitled in this work as BRASTEM, consisted of a novel ready-to-use formulation that just needs supplementation of basic FGF-2 (Life Technologies) to a final concentration of 8 ng/mL. This medium was recently developed using a sequential experimental design and validated at the Federal University of Rio de Janeiro and at the University of California at San Diego.31,32
To avoid the need for feeder cells or for MC coating in stirred cultures, all culture media (before FGF-2 supplementation) were conditioned by inactivated MEFs in cell culture plates before use in human ESC culture. After conditioning for 24 h at 37°C and 5% CO2, media were harvested, supplemented with FGF-2, and added to stirred flasks.
For the formation of embryoid bodies (EBs), H9 cells were maintained in stirred cultures, as described above, treated with TrypLE (Life Technologies) for 1 min, scraped out, and transferred to 60-mm nonadherent dishes in a medium containing high-glucose DMEM/F12 supplemented with additional 2 mM of
Screening of different MCs
Eight commercially available MCs (Table 1) were evaluated for their ability to promote adhesion of H9 cells. All MCs were prepared according to the corresponding manufacturer's instructions. Adhesion tests were performed in noncoated tissue culture six-well plates (TPP), which are known to not support the adhesion of hES. To each well, 2.5 mL of the KSR medium and an amount of MCs corresponding to a total surface area of 20 cm2were added (for each MC the ratio of surface area per mass was taken from the manufacturer's data and used to calculate the mass of beads to add). Cells were then inoculated at 0.1×106 cells/well. Plates were incubated at 37°C and 5% CO2, and after 72 h, bright field photomicrographs of each plate were taken. Supernatant was then discarded, cultured cells were detached using TrypLE (Life Technologies) for 3 min, pipette five to ten times to release the MC, and counted with an automatic cell counter (NucleoCounter, NC-100; Chemometech).
Evaluation of different culture media in stirred MC cultures
With the aim of developing a scalable xeno-free process for human ESC expansion, the MC considered the most effective in promoting adhesion of H9 cells (Hillex II; SoloHill) was used to evaluate the cell growth and maintenance of pluripotency of cells cultured in the conditioned KSR medium, conditioned KSR-XF and BRASTEM xeno-free media.
Cells were first grown in the xeno-free media in TPP for at least two passages to adapt them before inoculation of the stirred flasks. Stirred cultures were carried out in 125-mL spinner flasks (Techne) with a 60-mL working volume, at a MC concentration of 5 g/L, at 37°C, 5% CO2, and 40 rpm. Before inoculation, MCs were preincubated for at least 30 min in each conditioned medium. For inoculation, cells routinely grown as adherent monolayers were treated with warm Accutase (Millipore) for 2 min, and then released with a cell scrapper to obtain homogeneous clumps. To avoid inaccurate counts due to clumps, the automated counter equipment Nucleo Counter (NC-100; Chemometech) was used. Counts are performed after cells are lysed, clumps are broken, and nuclei are released into a homogenous solution. Later, samples are loaded into a cassette, while nuclei are automatically stained with propidium iodide. Cell counts are performed in an unbiased way. After counting, cells were inoculated at a concentration of 0.1×106 cells/mL.
To guarantee a good adhesion of cells to the MC beads, a protocol for intermittent agitation over 72 h was developed and adopted in this work. This protocol consisted of: during the first 24 h, intermittent agitation with 5 min stirring per each interval of 60 min, using the half working volume (30 mL); during the next 24 h (second day), intermittent agitation with 5 min stirring per each interval of 30 min, using the total working volume (60 mL); during the next 24 h (third day), intermittent agitation with 5 min stirring per each interval of 10 min; and continuous agitation from the fourth day on.
Daily, 50% of the culture supernatant was exchanged for a fresh conditioned medium containing 16 ng/mL FGF-2, to keep a final concentration of 8 ng/mL, assuming degradation of FGF-2 in the spinner flask. Before each medium exchange, a 1-mL sample was taken from each spinner flask and centrifuged at 200 g for 5 min. The supernatant was used for glucose and lactate determination, whereas the sedimented pellet was used for cell counting using a NucleoCounter (NC-100; Chemometech). All cell culture experiments were carried out, at least, as triplicates.
Immunocytochemistry
Instead of following conventional immunocytochemistry protocols, briefly, cells on the MCs were fixed with 4% paraformaldehyde and embedded in Tissue Tek® O.C.T™ (Japan), after which 10-μm sections were prepared with a cryostat. This strategy was adopted to reduce background of Hilex-II MCs due to auto-fluorescence. Immunocytochemistry was performed as described previously. 19 Primary antibodies targeting Oct-3/4 (Santa Cruz Biotechnology, Inc.; mouse, 1:100), Sox-2 (Chemicon-Temecula; rabbit, 1:100), TRA-1-81 (Chemicon-Temecula; mouse, 1:100), and SSEA-4 (Chemicon-Temecula; mouse, 1:100) were used, whereas the fluorophore-conjugated secondary antibodies were goat anti-mouse Alexa Fluor 488 green (Molecular Probes; 1:400) and goat anti-rabbit Alexa Fluor 546 orange (Invitrogen; 1:400 dilution). 4,6-diamino-2-phenylindole (DAPI) was used for nuclear staining.
Flow cytometry
Cells were harvested from MCs using TrypLE (Life Technologies), and then fixed in 2% paraformaldehyde and 0.3% Triton X-100 (Sigma-Aldrich) and immunolabeled using the same primary antibodies as for immunocytochemistry (against Oct-3/4, Sox-2, TRA-1-81, and SSEA-4). Goat anti-mouse Alexa Fluor 488 green (Molecular Probes; 1:400) was used as a secondary antibody. Cells were analyzed on a FACS Calibur™ flow cytometer (BD).
For Neu5Gc quantification, a kit (Gc-free basic kit) containing an anti-Neu5Gc antibody, an isotype control, and specific blockers was purchased (Gc-Free), and tests were performed as recommended by the manufacturer. Cells were analyzed on a fluorescence-activated cell sorting (FACS) Calibur flow cytometer (BD).
For both cases, to avoid cell debris and clumps and before analysis, cell populations were gated according to forward scatter and side scatter (data not shown). At least 10,000 counts were performed within the gated region before the percentages of each positive population were obtained.
Scanning electron microscopy
Samples of cells attached to MCs were washed with phosphate-buffered saline (PBS) at 37°C with a mixture of 2% glutaraldehyde (Sigma-Aldrich) and 4% formaldehyde (Sigma-Aldrich) in PBS at pH 7.4 for 1 h and washed three times with PBS. The samples were then dehydrated in a graded ethanol series and dried with hexamethyldisilazane (Sigma-Aldrich). Finally, the samples were sputter-coated with a thin layer of gold (Emitech; K550) and examined in a scanning electron microscope (JSM6460LV; JEOL, Japan) at 15 kV.
Glucose and lactate concentrations
Glucose and lactate concentrations were determined in supernatants from spinner samples using a YSI-2700 bioanalyzer (Yellow Springs Instruments). Specific lactate production and glucose consumption rates were calculated from the slope of the plot of cumulative concentration of lactate or glucose against the integral of viable cells. 33 Lactate-to-glucose yield (Ylac/glc) was determined by dividing the specific lactate production rate by the specific glucose consumption rate.
Western blot
For Western blot (WB) detection of Neu5Gc, a kit (Gc-free basic kit; GcFree) was used according to the manufacturer's instructions. Briefly, samples and a positive control (fetal bovine serum) were applied to a sodium dodecyl sulfate–polyacrylamide electrophoresis gel, and then transferred to a polyvinylidene fluoride membrane and blocked with the blocking solution provided by the kit. To determine the presence of Neu5Gc, the membrane was then exposed to an affinity-purified polyclonal chicken immunoglobulin Y (IgY) antibody specific for Neu5Gc and compared to an identical membrane exposed to an equivalent amount of nonspecific chicken IgY provided in the commercial kit.
RNA isolation and reverse transcription polymerase chain reaction assay
Total RNA was isolated using the Trizol reagent (Life Technologies) and digested using the DNA-free™ Ambion Kit (Life Technologies), following the manufacturer's instructions. Complementary DNA was generated from 1 μg of total RNA using the First Strand cDNA Synthesis kit (GE Healthcare Life Sciences), according to the manufacturer's protocol. Each RT-PCR reaction was carried out for 35 cycles using the GoTaq® Green Master Mix (Promega), 250 nM of each primer (Forward and Reverse), and 0.5 μL of cDNA for a final volume of 10 μL. cDNA samples were amplified by RT-PCR using the following primers and annealing temperatures (A.T.): hAFP (F: 5′-AGA GTT GCT AAA GGA TAC CAG GA-3′/R: 5′-AGG CCA ATA GTT TGT CCT CAC-3′/A.T.: 55°C), hFOXA2 (F: 5′-ACG CAA GGG AGA AGA AAT CCA-3′/R: 5′-TTT TAC ACC GAG TCA CTC ACA-3′/A.T.: 55°C), hSOX17 (F: 5′-GTG AAT CTC CCC GAC AGC-3′/R: 5′-TGT TTT GGG ACA CAT TCA AAG C-3′/A.T.: 56°C), hCK8 (F: 5′-AGC AGC AGA AGA CGG CTC GAA-3′/R: 5′-GAA GTT GAT CTC GTC GGT CAG C-3′/A.T.: 58°C), hCK18 (F: 5′-CAC ACA GTC TGC TGA GGT TGG AG-3′/R: 5′-CTG CTG TCC AAG GCA TCA CCA-3′/A.T.: 58°C), hBRACHYURY (F: 5′-TCT ACA TCC ACC CCG ACT-3′/R: 5′-GCT GTC TCC GGG TTC CTC-3′/A.T.: 58°C), hMSX1 (F: 5′-CCC TGG TGC TGT ACC CC-3′/R: 5′-GGT CCC TTC AAC CTA CCT T-3′/A.T.: 56°C), hGFAP (F: 5′-CCC TGG ACA TCG AGA TCG C-3′/R: 5′-TGT GCT CCT GCT TGG ACT C-3′/A.T.: 58°C), hMAP2 (F: 5′-CCT TTG AGA ACA CGA CAC AAC GAA-3′/R: 5′-GGA ATC CAT TGG CGC TTC-3′/A.T.: 55°C), hPAX6a (F: 5′-AGA AAG AGT TTG AGA GAA CCC AT-3′/R: 5′-TCA TGT GTG TCT GCA TAT GTG G-3′/A.T.: 58°C), hGAPDH (F: 5′-TTC GAC AGT CAG CCG CAT C-3′/R: 5′-GAC TCC ACG ACG TAC TCA GC-3′/A.T.: 58°C).
Statistical analyses
Data are presented as means±standard error from at least three independent experiments. Statistical analyses were performed using the Student t-test for the case 2 groups were compared. For multiples groups being analyzed at the same time, analysis of variance followed by the Tukey's test was performed. All p-values lower than 0.05 were considered statistically significant.
Results
Screening of MCs
Eight commercially available MCs (Table 1) were evaluated for their ability to promote adhesion of H9 cells. The recovery of cells adhered after incubation for 72 h is shown in Figure 1A. The yield of cells that could be recovered from beads was taken as the response for these experiments, since the performance of a larger scale stem cell expansion process will depend on the amount of cells that can be harvested for final application.

Screening of MCs using H9 cells grown in the KSR medium using noncoated plates. Inoculum 0.1×106 cells/well. Yield of cells recovered from each MC after 72 h
This comparative analysis of MCs was motivated by the fact that most protocols for cell expansion on MCs found in literature use beads coated with Matrigel™,18,20,34 or with collagen, 19 hampering their use for obtaining cells for human therapy. Moreover, lag phases reported in literature were long, for example, 8 days in the report by Fernandes et al., 19 decreasing process cost–effectiveness.
In this work, MCs coated with animal collagen (Cytodex-3, Collagen, and FACT) were among those promoting the best adhesion, as expressed by the total amount of cells that could be recovered (0.040±0.003, 0.046±0.001, and 0.039±0.004 million cells per well, respectively). Use of the MC coated with recombinant human fibronectin (Pronectin F) resulted in a poorer cell yield (0.035±0.003 million cells per well), and most noncoated MCs displayed an even worse performance. Use of the Hillex II MCs enabled the recovery of the highest amount of cells (0.069±0.004 million cells per well), thereby qualifying this MC as the one with the best performance among the MCs tested in this work (p<0.001, Tukey's test). H9 cells incubated with Hillex II MCs for 3 days were able to form three-dimensional-like colonies (Fig. 1I) that were not observed upon incubation with the other MCs (Fig. 1B–H).
The present results suggest that protein-coated MCs in general promote a good cell attachment, but that a strong positively charged surface, such as the triethylammonium groups on Hillex II MCs, can promote even better cell adhesion, which is in accordance with previous reports. 30
Evaluation of xeno-free media in stirred MC cultures
With the aim of developing a xeno-free protocol for human ESC expansion, two xeno-free media (KSR-XF and BRASTEM) were evaluated in comparison with the standard KSR medium in stirred MC cultures. Due to the best performance shown by Hillex II, this MC was used in these experiments.
Using the KSR medium, cultures of H9 cells with Hillex II (Fig. 2) using the adhesion protocol described in the present work showed a shorter lag phase (6 days) than the 8 days observed in our previous study using Cytodex-3. 19 Maximum cell concentration achieved in the present work (1.72±0.14×106 cells/mL) was similar to that obtained in the previous work with Cytodex-3, but it was achieved quicker, in 264 h, probably due to the combination of the new adhesion protocol and the better adhesion profile provided by the Hillex II MCs. The specific growth rate observed during the exponential growth phase in this work was 0.0230±0.0012 h−1 (R 2 >0.95), resulting in a doubling time during the exponential phase of 30.3±1.5 h.

H9 cells growth kinetics on Hillex II MCs (-♦-) BRASTEM medium, maximum cell concentration 2.78±0.13×106 cells/mL; (-▪-) KSR medium, maximum cell concentration 1.70±0.04×106 cells/mL; (-▴-) KSR-XF medium, maximum cell concentration 1.72±0.14×106 cells/mL. Error bars were obtained from at least three independent experiments (n≥3). KSR XF, knockout serum replacement xeno-free.
The KSR-XF medium was prepared in the same manner as the KSR medium, but using the xeno-free version of KSR commercialized by the same manufacturer. Thus, as expected, a similar growth profile was obtained in both media (Fig. 2), with H9 cells cultured in the KSR-XF medium achieving 1.70±0.04×106 cells/mL. However, the novel xeno-free, animal/human-component-free medium (BRASTEM) developed by Marinho 31 and Muotri et al. 32 provided a significantly better performance. Adaptation of cells to this medium was significantly superior, and the lag phase was reduced to 2 days. Cells grew faster, resulting in a doubling time of 25.3±1.1 h in the exponential growth phase. The maximum cell concentration achieved (2.78±0.13×106 cells/mL) was significantly higher (p<0.05) than in all other experiments, and was reached in a much shorter period (6 days).
To further evaluate cell growth on MCs in the different media, scanning electron micrographs of samples withdrawn after 24 h of culture were taken. As shown in Figure 3A, cells grown in the BRASTEM medium showed a more evenly distributed colonization of the MCs than in KSR and KSR-XF media (Fig. 3C, E). Furthermore, in both KSR-based media there was a clear tendency to the formation of large clumps of beads, whereas in BRASTEM media cells proliferated also on isolated beads. When taken together, parameters related to cell expansion and productivity (Table 2) showed a clearly superior performance of cells grown in the BRASTEM medium over the two KSR-based media. The advantages of the BRASTEM medium were further corroborated by its cost, which was estimated to be approximately 35% and 54% lower than those of KSR and KSR-XF media, respectively, taking into consideration the medium and supplement prices in the United States of America in 2012 (data not shown).
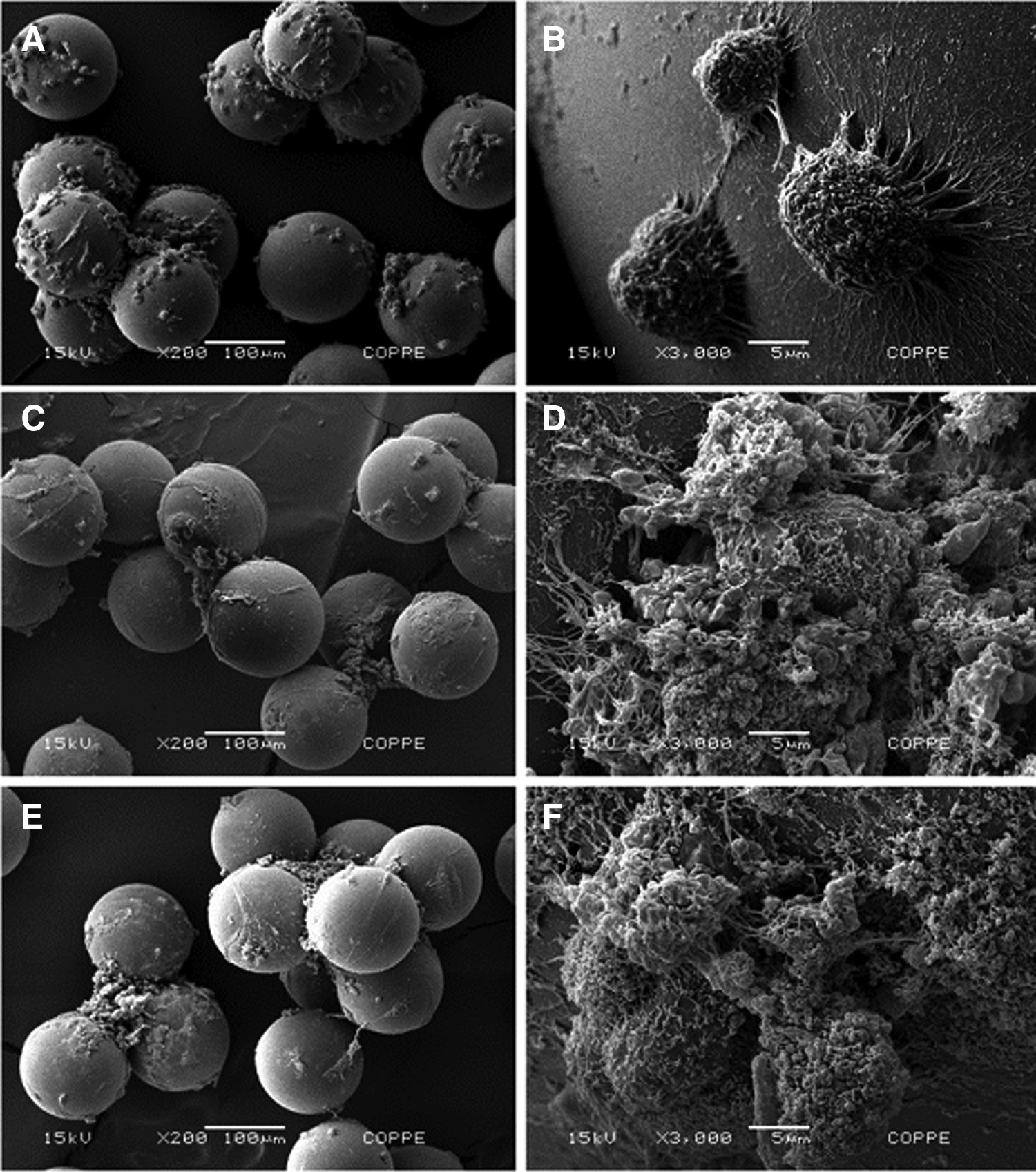
Scanning electron microscopy of H9 cells adhered to Hillex II MCs cultured for 24 h in: BRASTEM medium at 200×
KSR, knockout serum replacement; KSR XF, knockout serum replacement xeno-free.
To evaluate cell metabolism under the different culture conditions, glucose and lactate concentrations were determined (Table 2). Cultures with the BRASTEM medium showed a lactate-to-glucose ratio higher (1.69 mol/mol) than cultures with KSR-based media, but still in the range observed by other authors, for human ESC cultured with the serum-free media mTeSR1 and StemPRO. 35 The larger lactate-to-glucose ratios indicate a high-glucose transport rate and a high flux through the glycolytic pathway, resulting in a substantial glucose consumption and lactate production. Lactate accumulation could trigger the need for automatic pH control when scaling-up cultures in bioreactors.
To compare the three media also in terms of their ability to maintain pluripotency, cells harvested in the last day of culture had the expression of pluripotency markers evaluated by both flow cytometry and immunocytochemistry. Both nuclear (OCT-4 and SOX-2) and surface (SSEA-4 and TRA1-81) markers were observed (Fig. 4A–C) in slices of MCs colonized with cells, indicating that pluripotency of cells was maintained in cell–bead aggregates cultured with the BRASTEM medium.
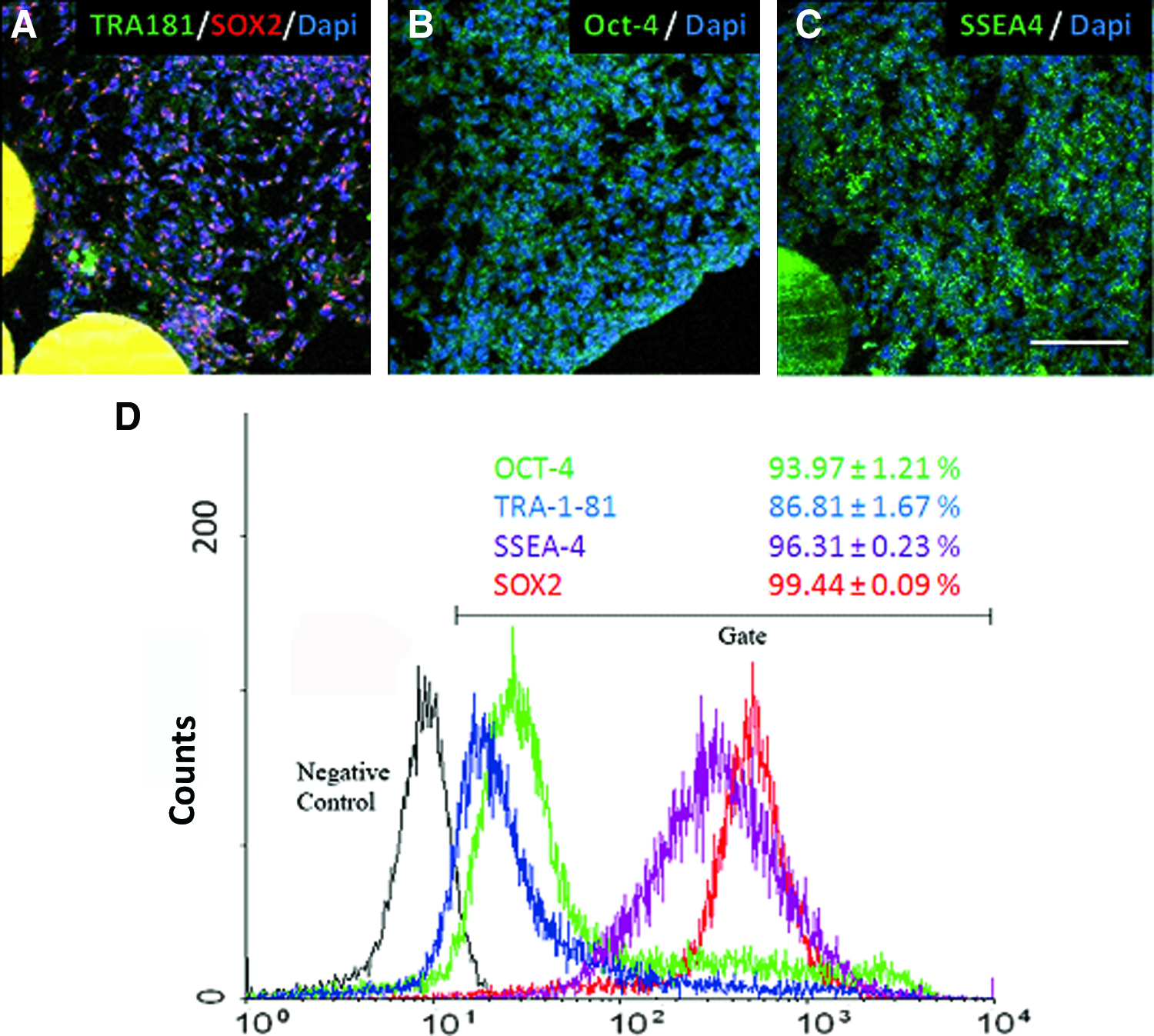
Immunocytochemistry of H9 cells cultured 6 days in spinner flasks attached to Hillex II MCs in the BRASTEM medium. Pluripotency markers were evenly detected from the borders of the colonies to the inner region, closer to MCs: TRA-1-81, green and SOX-2, red
Flow cytometry analysis of BRASTEM cultures (Fig. 4D) quantitatively confirmed a large population of cells positive for pluripotency markers (93.97±1.21% for OCT-4, 99.44±0.09% for SOX-2, 96.31±0.23% for SSEA-4, and 86.81±1.67% for TRA1-81), indicating that this novel animal/human-component-free medium does not induce ES differentiation, which was an issue in earlier reports employing the KSR-based medium.18,30
To confirm the ability of H9 cells cultured with the BRASTEM stirred culture system to differentiate, these cells were grown as EBs. After 14 days in suspension, H9 cells formed spherical EBs (Supplementary Fig. S1A; Supplementary Data are available online at
Reduction of nonhuman sialic acid incorporation
Nonhuman components are usually present in ESC standard media22,23 and their presence was confirmed in the present work by analyzing samples of fresh and MEF-conditioned KSR medium for Neu5Gc (Fig. 5A, lanes 4 and 5). As expected, Neu5Gc could not be detected in fresh samples of KSR-XF and BRASTEM xeno-free media (Fig. 5A, lanes 2 and 6, respectively). Even when these media were conditioned by murine MEFs (Fig. 5A, lanes 3 and 7, respectively), Neu5Gc contamination was significantly lower than in analogous samples of cultures grown in the KSR medium, suggesting that the main source for Neu5Gc might be the medium rather than feeder cells.
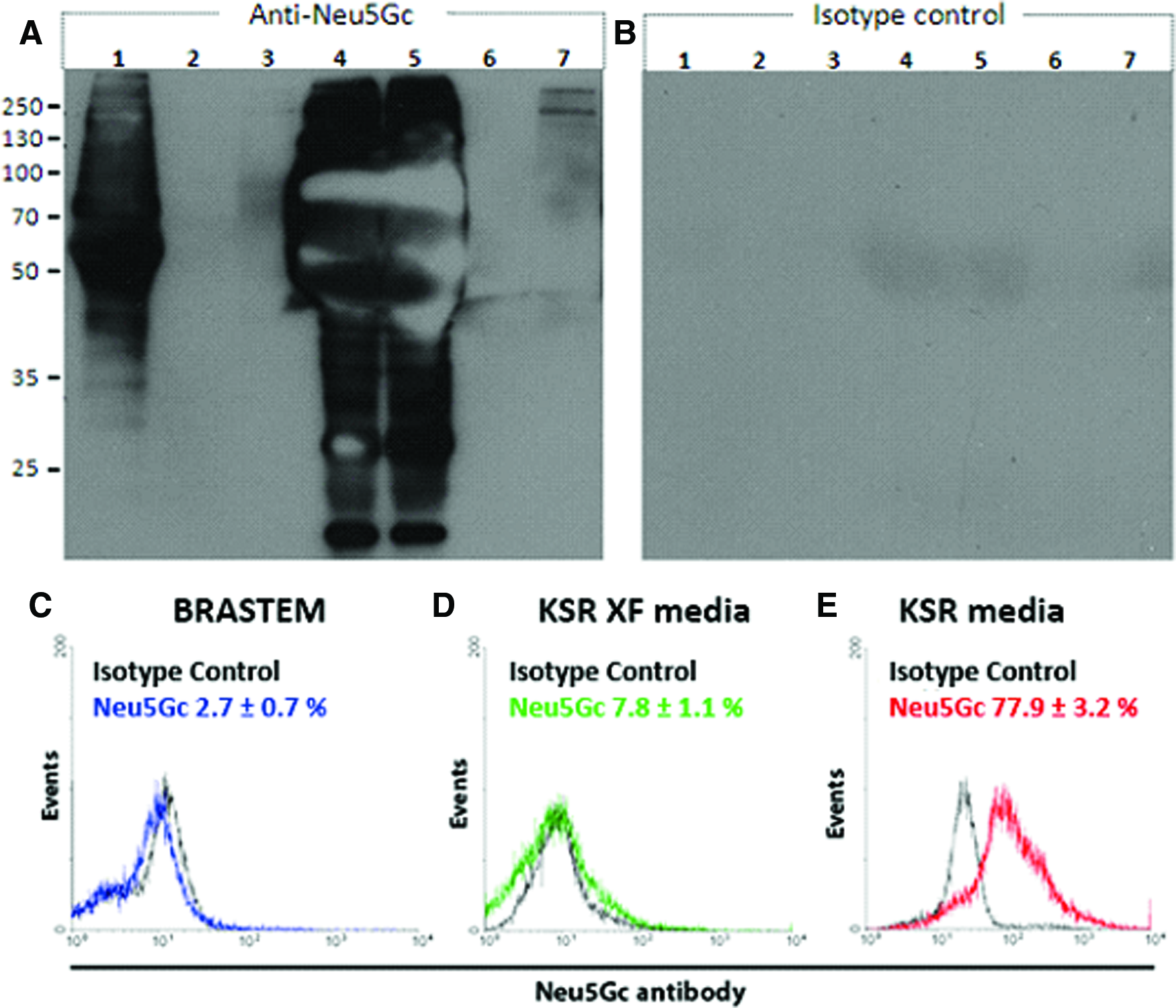
WB for Neu5Gc antibodies.
To test the hypothesis that the medium accounts for most of the contamination with Neu5Gc, human ESC previously cultured in the KSR medium were expanded in spinner flasks with Hillex II MCs using KSR, KSR-XF, and BRASTEM media (all of them previously conditioned by MEF) and analyzed by flow cytometry for the presence of Neu5Gc. Flow cytometry data (Fig. 5C) showed a significantly lower occurrence of Neu5Gc in cells grown with the xeno-free media (7.79±1.15% and 2.75±0.67% for KSR-XF and BRASTEM media, respectively), when compared to the KSR medium (77.94±3.25%). These results indicate that xeno-free media are able to significantly decontaminate cells that have previously been in contact with Neu5Gc within as little as one-passage. This finding represents a major breakthrough in terms of regulatory-friendly cell production for human therapy, since Neu5Gc is known to cause immunogenic responses. 25
Discussion
Fulfillment of the biotechnological and industrial potential of stem cells depends on the availability of large amounts of high-quality cells to be used for drug screening, in vivo animal testing, and human cell therapy. However, developing scalable and economically feasible processes for in vitro human ESC expansion remains a challenge. 36 Recently, published articles in the field have proposed optimized protocols; however, many of them were based on murine ESC.37,38
The use of Hillex II MCs established in this work provides an efficient way of expanding cells in stirred, scalable systems using an animal/human-component-free support for cell adhesion. In contrast to other protocols, which reported the need to coat MCs, for example, with Matrigel,18,21 it was shown in this work that for Hillex II MCs no coating with commercial matrices or animal-derived proteins is needed to culture ESC. This represents an advantage in terms of process costs and regulatory issues.
Compared to the protocol described herein, previous articles reported an initial adhesion phase consisting of either identical agitation periods (i.e., 5 min) combined to longer (e.g., 6 h) resting periods 30 or simply a long 24-h low-speed stirring period. 18 We initially observed that a 24-h adhesion phase could not satisfactorily promote cell attachment to MCs, and further verified that good cell adhesion was only achieved when stirring/resting cycles were maintained for 72 h (data not shown). We hypothesized that extracellular matrix produced over the first 72 h on culture protect cells from continuous shear-forces, and consequently improves growth. Human ESC produce extracellular matrix while on MCs. Around 72 h, a complete layer of extracellular matrix is observed above those cells (Supplementary Fig. S2). This extracellular matrix may behave as a shield against the forces generated by the continuous agitation. The consequences of a faster adaptation period include the reduction of the lag phase when compared to data described on our previous work using KSR media. 19
The use of serum-free culture media had already been described,38–41 but optimization of specific formulations of medium components was never pursued. In our work, the use of a novel optimized, animal/human-component-free medium formulation31,32 yielded a doubling time of approximately 25 h, with a threefold increase in cell productivity, from 0.006±0.001×106cells/h (KSR-based media) to 0.019±0.001×106cells/h (BRASTEM medium), resulting in a final concentration of 2.78±0.13×106cells/mL (Table 2). Nonetheless, variables, such as agitation speed, MC concentration, should be taken into account to reduce cell agglomeration. 42 Since the BRASTEM medium was the one which achieved the highest concentration of cells and therefore increasing the probability of clump formation, it is reasonable to expect that improvements in culture conditions may have a better impact on the BRASTEM system than others, making it even more attractive.
Even though cells proliferated at high rates, they were capable of maintaining the expression of pluripotency markers at high levels during all the period of growth. Immunocytochemistry observations were corroborated by quantitative flow cytometry analysis, showing expression of four pluripotency markers, in more than 93% of the cells for OCT-4, SOX-2, and SSEA-4, and more than 86% of the cells for TRA1-81.
Considering the high cell growth achieved and the maintenance of pluripotency observed, our data confirm stirred MC systems as the currently best alternative for scaling-up production of human ESC. The use of the xeno-free, chemically defined, animal/human-component-free medium BRASTEM was reported here for the first time and showed that its use in combination with the xeno-free Hillex II MCs provides a promising regulatory-friendly way to produce large amounts of low-immunogenic human ESC. Using the system for long-term maintenance (serial passages) needs further evaluation. However, by culturing human ESC with BRASTEM into a small spinner flask system, 160 million of noncontaminated cells were obtained after less than a week, making the system feasible for short-term expansions need in future clinical trials.
Footnotes
Acknowledgments
The authors acknowledge funding from the Brazilian public agencies CAPES, FAPERJ, CNPq, FINEP, and BNDES. Special thanks also to Dr. Sylvie Devalle (National Laboratory for Embryonic Stem Cell Research, Brazil) for reviewing the article and Dr. Marcia Sader (Metallurgical and Materials Engineering—COPPE/UFRJ) for assistance with scanning electron microscopy.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.