
Abstract
Human infrapatellar fat pad contains a source of mesenchymal stem cells (FPSCs) that potentially offer a novel population for the treatment of damaged or diseased articular cartilage. Existing cartilage repair strategies such as microfracture harness the presence of a low-oxygen microenvironment, fibrin clot formation at sites of microfracture, and elevations in growth factors in the damaged joint milieu. Bearing this in mind, the objective of this study was to determine the chondrogenic potential of diseased human FPSCs in a model system that recapitulates some of these features. In the first phase of the study, the role of transforming growth factor beta-3 (TGF-β3) and fibroblast growth factor-2 (FGF-2), in addition to an altered oxygen-tension environment, on the colony-forming unit-fibroblast (CFU-F) capacity and growth kinetics of human FPSCs during monolayer expansion was evaluated. The subsequent chondrogenic capacity of these cells was quantified in both normoxic (20%) and low- (5%) oxygen conditions. Expansion in FGF-2 was shown to reduce CFU-F numbers, but simultaneously increase both the colony size and the cell yield compared to standard expansion conditions. Supplementation with both FGF-2 and TGF-β3 significantly reduced cell-doubling time. Expansion in FGF-2, followed by differentiation at 5% oxygen tension, was observed to synergistically enhance subsequent sulfated glycosaminoglycan (sGAG) accumulation after chondrogenic induction. FPSCs expanded in FGF-2 were then encapsulated in either agarose or fibrin hydrogels in an attempt to engineer cartilaginous grafts. sGAG synthesis was higher in fibrin constructs, and was further enhanced by differentiation at 5% oxygen tension, accumulating 2.7% (ww) sGAG after 42 days in culture. These results indicate that FPSCs, a readily accessible cell population, form cartilage in an in vitro environment that recapitulates several key biological features of cartilage repair during microfracture and also point toward the potential utility of such cells when combined with fibrin hydrogel scaffolds.
Introduction
Therapeutic tissue-engineering applications require the reliable production of relatively large numbers of donor-specific cells 13 ; therefore, the identification of MSC expansion conditions that accelerate proliferation while maintaining multipotency is important for clinical applications. Supplementation of expansion media with various growth factors has been explored in an attempt to improve the isolation and expansion of progenitor cells isolated from different tissues.14–17 For example, fibroblast growth factor-2 (FGF-2) has been shown to enhance the proliferation of bone marrow-,13,18–21 adipose-, 22 periosteum-, 23 and IFP-24,25 derived MSCs and to delay the loss of chondrogenic potential in bone marrow-derived MSCs with passaging. 26 In the case of FPSCs, expansion in the presence of FGF-2 has also been shown to enhance their subsequent chondrogenic potential. 24 Various subtypes of transforming growth factor beta (TGF-β) have also been added to expansion media to enhance proliferation and subsequent chondrogenesis of chondrocytes and MSCs.15,27–29 Of note is the finding that chondrocytes expanded with FGF-2 and TGFβ1 displayed the lowest doubling times during expansion and subsequently generate cartilaginous tissues with the highest accumulation of glycosaminoglycans and collagen type II. 30 The in vitro maturation of cartilage tissue is also enhanced using a similar combination of growth factors. 31
Oxygen tension has also been shown to regulate MSC proliferation and differentiation. 32 Expansion of MSCs at a low oxygen tension has been shown to increase cell yield and reduce doubling times,33–36 as well as promoting the chondrogenic potential of cells isolated from bone marrow,37–39 adipose tissue, 40 and cartilage.41,42 Furthermore, enhanced chondrogenesis has been reported for MSCs undergoing differentiation at low oxygen tensions,38,43–45 although there are also reports of hypoxic conditions inhibiting chondrogenesis.48,49 For IFP-derived MSCs, chondrogenic differentiation at 5% oxygen has been shown to increase expression of hypoxia-inducible transcription factor 2α, reduce cell proliferation, and enhance matrix synthesis. 50 The functional properties of cartilaginous tissues engineered using IFP-derived MSCs are also improved in a low-oxygen environment. 51
Clearly, the proliferation and differentiation capacities of MSCs isolated from multiple tissue types are strongly dependent on the local environment; however, the expansion and differentiation conditions required to engineer truly functional cartilaginous grafts using human MSCs remain to be elucidated. Spontaneous regenerative events in the body may provide clues as to how we should engineer such tissues. Specifically, existing cartilage repair strategies such as microfracture, although imperfect, harness the presence of a low-oxygen microenvironment, 52 fibrin deposition at sites of microfracture, 53 and elevations in growth factors 54 in the damaged joint environment to induce both the proliferation and differentiation of MSCs. Bearing this in mind, the objective of this study was to determine the proliferation and differentiation potential of human FPSCs in a model system mimicking aspects of this environment. The first phase of the study assessed the role of TGF-β3 and FGF-2, in addition to an altered oxygen tension environment, on the colony-forming unit-fibroblast (CFU-F) capacity and growth kinetics of human FPSCs during monolayer expansion. The subsequent chondrogenic capacity of these cells was then quantified in normoxic (20%) and low- (5%) oxygen conditions in a pellet culture system. Having identified appropriate expansion conditions, we next sought to determine the effect of a biomaterial substrate (agarose and fibrin hydrogels) and biochemical cues in either low- (5%) or normoxic (20%) oxygen environments on the development of cartilaginous tissues engineered using human FPSCs. We hypothesised that in vitro conditions mimicking aspects of the regenerative environment of microfracture, specifically encapsulation in fibrin hydrogels and maintenance in low-oxygen conditions, would facilitate the development of cartilaginous tissue engineered using FPSCs.
Materials and Methods
Cell isolation and expansion
Ethics approval for the study was obtained from the institutional review board of the Mater Misericordiae University Hospital with IFPs being obtained from seven patients with knee osteoarthritis (OA) at joint arthroplasty. The IFPs were washed thoroughly in phosphate-buffered saline (PBS), and tissue specimens were finely diced with care taken to exclude fibrotic regions. Samples were weighed and incubated with 0.25% (w/v) collagenase (Worthington Biochemical) for 4 h at 37°C under constant rotation. After tissue digestion, cells were washed in PBS, filtered through a 40-μm nylon cell strainer, and centrifuged at 650 g for 5 min. Supernatant was discarded to remove floating adipocytes, and cells were resuspended in a standard expansion medium (EM) consisting of a high-glucose Dulbecco's modified Eagle's medium (DMEM) containing 10% foetal bovine serum and antibiotics (100 units/mL penicillin and 100 μg/mL streptomycin) (all from GIBCO, Biosciences). After counting in 3% final acetic acid (to lyse blood cells) and Trypan blue, cells were seeded at a density of 5×103 live cells/cm2 in T-75 flasks (Sarstedt). Further groups were expanded in an EM supplemented with FGF-2 (5 ng/mL), TGF-β3 (0.5 ng/mL), or both (all from ProSpec-Tany TechnoGene, Ltd.). The concentration of growth factors utilized here was motivated by previous studies.15,24,28,30
All groups were expanded at either 5% or 20% oxygen. Nonadherent cells were removed at the first medium change after 72h. After colony formation, during passage 0 (P0) (8–10 days), cells were detached with 0.05% trypsin–EDTA (Sigma-Aldrich) and subcultured at a density of 5×103 cells/cm2 (P1). Cultures were expanded to passage two (P2) at a seeding density of 5×103 cells/cm2. Complete medium exchanges were performed twice weekly.
CFU-F assay
For the CFU-F assay, freshly isolated cells from four donors were plated in 58-cm2 petri dishes at a density of 135 cells/cm2 with EM and the various growth factor formulations. Triplicate dishes were plated for all conditions. After 14 days, cells were fixed with 2% paraformaldehyde (PFA), stained with 1% crystal violet (Sigma-Aldrich), and colony numbers (>50 cells) counted. The total colony number was used to determine the total number of population doublings that occurred during P0 and hence the population doubling time. For the purposes of comparison between groups, the colony size was defined as the average diameter of the 10 largest colonies formed.
In vitro chondrogenesis in pellet culture
P2 cells (250,000) from four donors were centrifuged in 1.5-mL microtubes by centrifugation at 650 g for 5 min. Pellets were cultured for 21 days in 1 mL of serum-free medium consisting of high glucose DMEM supplemented with a standard antibiotic mixture (100 units/mL penicillin and 100 μg/mL streptomycin; Invitrogen), 100 μg/mL sodium pyruvate, 40 μg/mL
Engineering cartilaginous grafts in fibrin and agarose hydrogels
IFPs were obtained from three different patients with knee OA at joint arthroplastly. FPSCs were isolated as described above, and expanded at 20% oxygen in a medium supplemented with 5 ng/mL FGF-2. FPSCs from all three donors were pooled before hydrogel encapsulation to create a superlot. In the first study, expanded cells were suspended in either 2% agarose (Type VII; Sigma) or fibrin hydrogels at a density of 20 million cells/mL. Fibrinogen (Sigma-Aldrich) was dissolved in aprotinin solution (Nordic Pharma). Cells were mixed with this solution, and thrombin was added to crosslink the gel. The solution was immediately injected into an agarose mold to yield fibrin cylindrical hydrogels (50 mg/mL fibrinogen, 5000 KIU/mL aprotinin, and 2.5 U/mL thrombin final concentrations) with the same dimensions as the agarose hydrogels. Construct cylinders of dimensions Ø5×2 mm were produced and maintained in a chemically defined chondrogenic medium supplemented with TGF-β3 (10 ng/mL) at either 20% or 5% oxygen. In a second study, fibrin hydrogels were prepared in the same manner and cultured in a standard (STD) medium formulation with or without the addition of 5% fetal bovine serum (FBS) (STD + 5% FBS), and cultured at either 20% or 5% oxygen. Constructs were maintained in culture for 42 days.
Biochemical analysis
Pellets (n=4 per experimental group for each of the four donors) and hydrogels (n=4 per experimental group using FPSCs from the superlot) were digested in papain (125 μg/mL) in 0.1 M sodium acetate, 5 mM cysteine HCl, and 0.05 M EDTA, pH 6.0 (all from Sigma-Aldrich), at 60°C for 18 h. Total DNA content was measured using a Quant-iT™ PicoGreen® dsDNA kit (Molecular Probes, Biosciences) with a lambda DNA standard. The proteoglycan content was estimated by quantifying the amount of sulfated glycosaminoglycan (sGAG) in digested pellets using the dimethylmethylene blue dye-binding assay (Blyscan, Biocolor Ltd.), with a chondroitin sulfate standard.
Histology
Pellets and hydrogels (n=2 per group) were fixed in 4% PFA (Sigma-Aldrich), wax-embedded, and sectioned at 5 μm. Sections were stained for sGAG with 1% alcian blue 8GX (Sigma-Aldrich) in 0.1 M HCl, and for collagen with picrosirius red.
The deposition of collagen types I, II, and X was identified through immunohistochemistry. Briefly, sections were rinsed with PBS, quenched of peroxidase activity for 20 min, and treated with chondroitinase ABC (Sigma; 0.25 units/mL) in a humidified environment at 37°C for 1 h to enhance permeability of the extracellular matrix by removal of chondroitin sulfate. Slides were again rinsed with PBS and blocked with 10% goat serum for 30 min. Sections were incubated with collagen type I (ab6308, 1:400; 5.7 mg/mL), collagen type II (ab3092, 1:100; 1 mg/mL), or collagen type X (ab49945, 1:200; 1.4 mg/mL) primary antibody (mouse monoclonal; Abcam) for 1 h at room temperature. After washing in PBS, the secondary antibody for type I and type II collagen (anti-mouse IgG biotin antibody produced in goat) (concentration 1 g/L) binding was applied for 1 h. Color was developed using the Vectastain ABC reagent (Vectastain ABC kit; Vector Laboratories) for 45-min and 5-min exposure to peroxidase DAB substrate kit (Vector laboratories). Slides were dehydrated through ethanol and xylene and mounted with a Vectamount medium (Vector Laboratories). Positive and negative controls (cartilage and ligament) were included in the immunohistochemistry staining protocol for each batch.
Statistical analysis
GraphPad Prism (Version 4.3) software was used for all statistical analyses. Numerical and graphical results are reported in the form of mean±standard error from the mean. Groups were compared with two-way analysis of variance with Bonferroni post-tests, with oxygen tension and growth factor formulation as the independent variables.
Results
Expansion in FGF-2 alters the colony-forming unit capacity of diseased human FPSCs
Colonies generated from FPSCs expanded in FGF-2 generally appeared larger than those expanded in a standard EM, particularly in lower-oxygen environments (Fig. 1). When quantified, significant donor variability in CFU-F numbers was observed (Fig. 2). Both growth factor supplementation (p<0.001) and oxygen tension (p<0.005) had an overall effect on the CFU-F efficiency. Supplementation of EM with FGF-2 gave rise to significantly lower colony numbers compared to standard expansion conditions (3/4 donors). In general, expansion in the presence of TGF-β3 did not significantly affect colony numbers compared to the standard EM (3/4 donors).

CFU-F assay (donor 1). Similar results were observed for all donors. CFU-F, colony-forming unit fibroblast; FGF-2, fibroblast growth factor-2; STD, standard; TGF-β3, transforming growth factor beta-3. Color images available online at
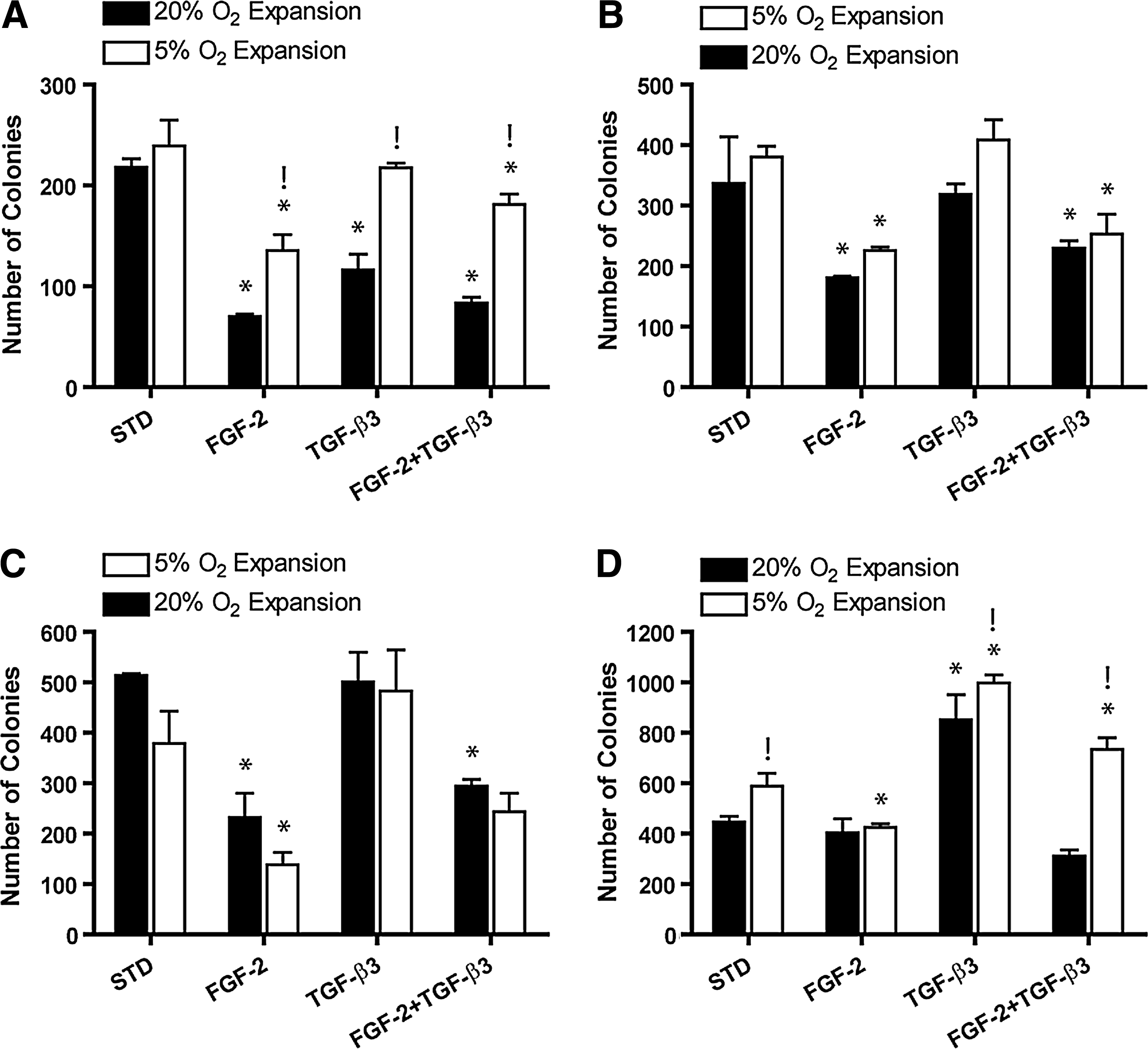
Total number of colonies determined from CFU-F assay.
Both growth factor supplementation (p<0.0001) and oxygen tension (p<0.005) had an overall effect on the colony size (all donors pooled), with significantly larger colonies observed in the FGF-2-supplemented medium at both oxygen tensions compared to the standard EM (p<0.05) (Fig. 3A). Smaller colonies were observed in the TGF-β3-only group at 5% oxygen compared to the standard EM (p<0.05). Colony size in the FGF-2 group was significantly greater at 5% oxygen compared to 20% oxygen (p<0.01).

Oxygen tension did not significantly affect the cell-doubling time during passage 0 (Fig. 3B). There was a greater number of population doublings during P0 in the FGF-2-supplemented medium compared to the standard expansion medium at 20% oxygen (p<0.05) (Fig. 3C). Supplementation with both FGF-2 and TGF-β3 was observed to significantly reduce cell-doubling times compared to the standard expansion medium (p<0.05) (Fig. 3B). The number of population doublings during P0 was also greater for cells expanded in the presence of both FGF-2 and TGF-β3 (Fig. 3C).
Expansion in FGF-2 followed by differentiation in a low-oxygen environment synergistically enhances the chondrogenic potential of diseased human FPSCs
Expansion in the presence of FGF-2 significantly enhanced sGAG production (sGAG/DNA) for FPSCs undergoing chondrogenic differentiation in pellet culture compared to cells expanded in a standard EM (4.3±0.08 μg/μg vs. 2.3±0.013 μg/μg; p<0.001) (Fig. 4). This result was also observed for MSCs expanded at 5% oxygen (data not shown). Expansion in the presence of FGF-2, followed by differentiation at 5% oxygen, resulted in the highest levels of sGAG production (5.9±0.34 μg/μg; p<0.05). Expansion in the presence of TGF-β3, or both FGF-2 and TGF-β3, did not significantly improve sGAG accumulation within pellets compared to standard expansion conditions (data not shown). All pellets stained positive for Alcian blue and picrosirius red, with pellets expanded in the presence of FGF-2 generally appearing larger and more intensely stained, particularly around the periphery of pellets (Fig. 5). Pellets stained positively for type II collagen, indicating the formation of a cartilage-like extracellular matrix.
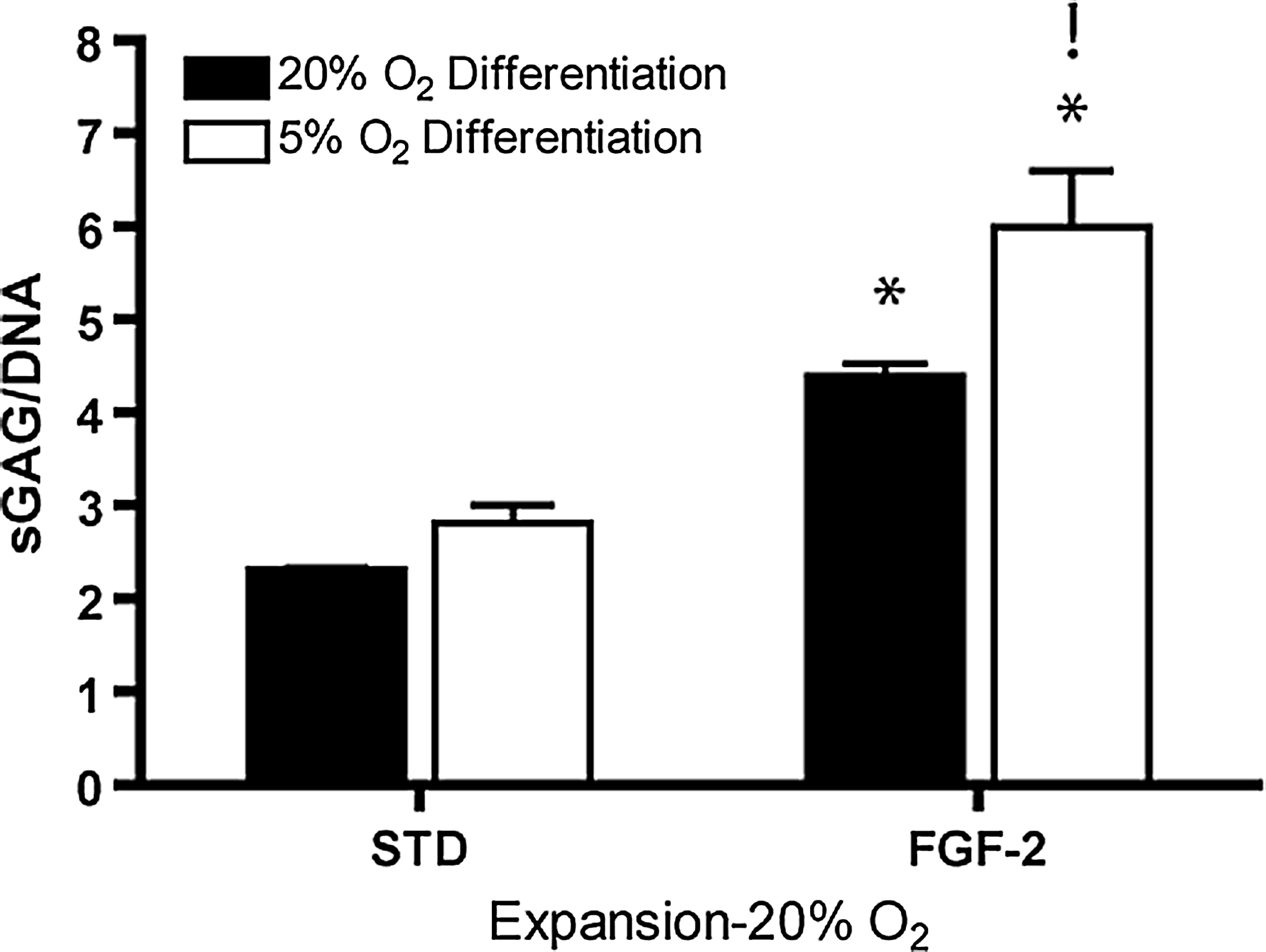
Sulfated glycosaminoglycan (sGAG)/DNA of pellets after 21 days of culture, which had been expanded at 20% oxygen tension and differentiated at either 20% or 5% oxygen tension. (*) Significance compared to STD medium formulation (p<0.001). (!) Significance compared to the same medium formulation (p<0.05).
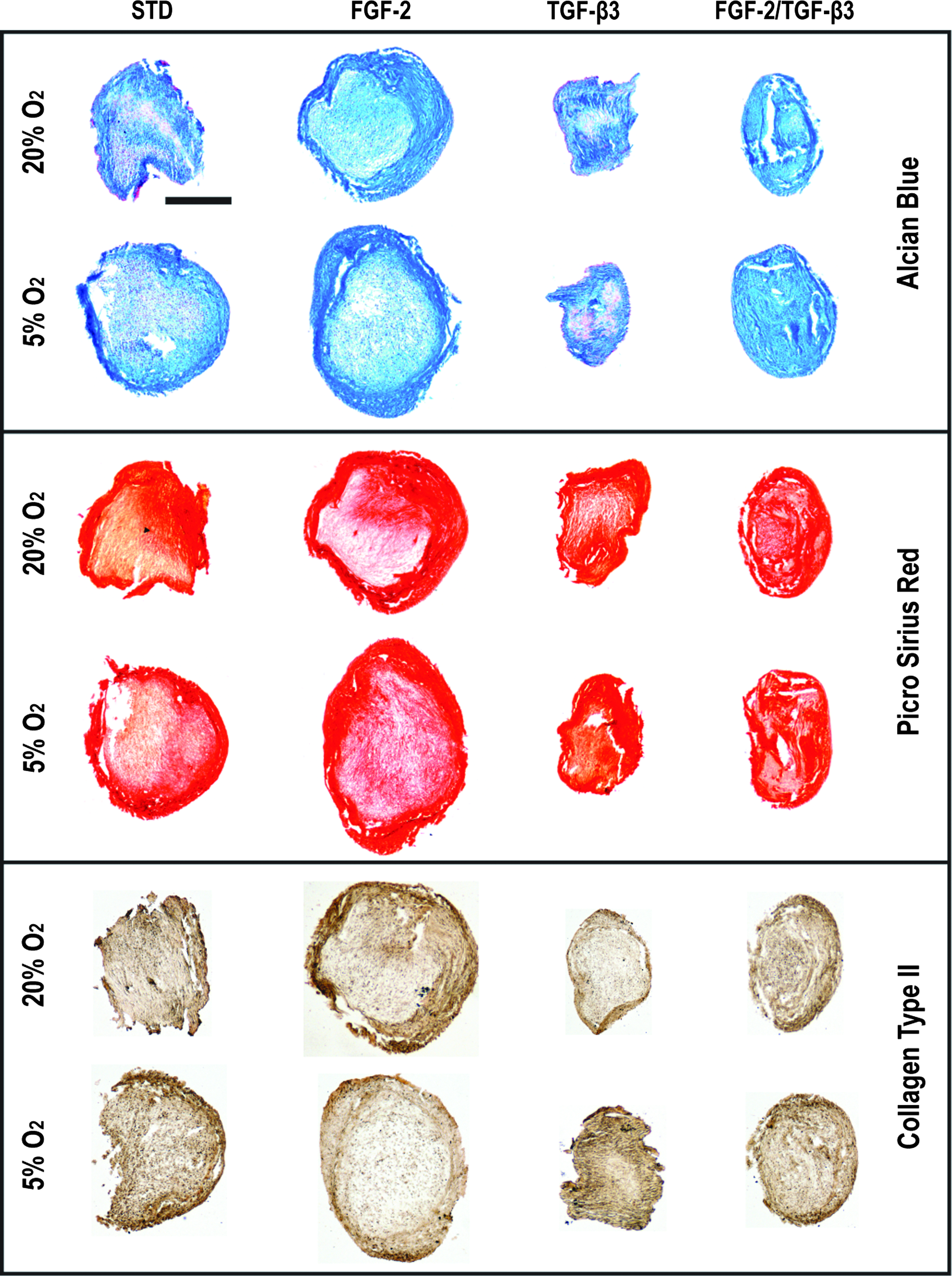
Picrosirius red, Alcian blue, and type II collagen staining of pellets (day 21) expanded at 20% O2 and differentiated at either 5% or 20% O2. Color images available online at
Encapsulation in fibrin hydrogels and maintenance in a low-oxygen environment support the development of cartilaginous grafts engineered using diseased human FPSCs
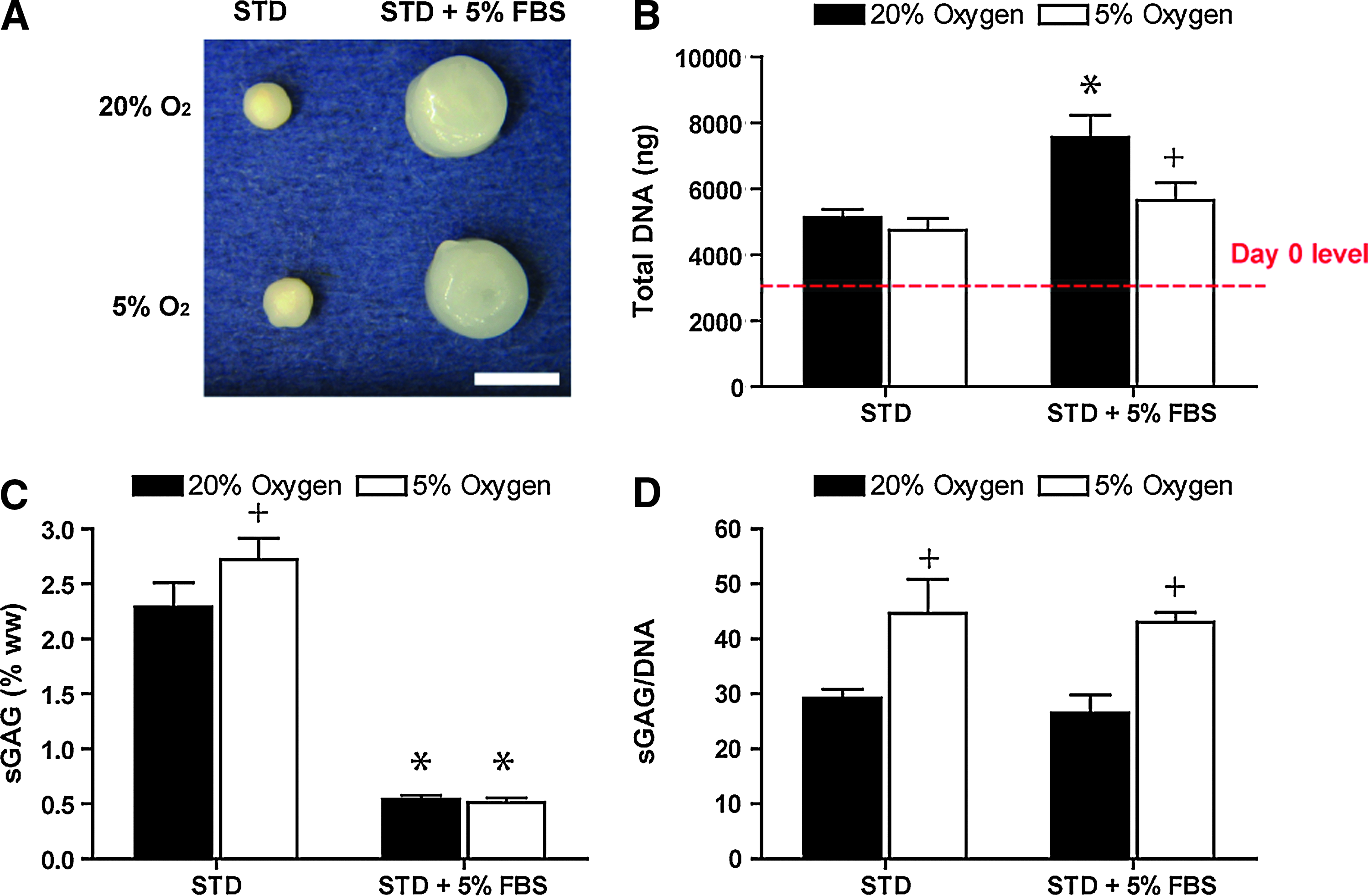
Having identified expansion and differentiation conditions that promoted robust chondrogenesis of FPSCs in pellet culture, we next explored the use of cells expanded in FGF-2 and encapsulated into either fibrin or agarose hydrogels for engineering functional cartilaginous grafts. After 42 days in culture, there was a significant decrease in DNA content in agarose hydrogels, but an increase for fibrin hydrogels over day-0 levels. In addition, sGAG production was higher in fibrin hydrogels (Fig. 6), and was further enhanced by differentiation in low-oxygen conditions; however, significant contraction of fibrin hydrogels occurred after 42 days of culture (Fig. 7). Fibrin constructs also stained positively for sulfated proteoglycan and type II collagen, but only weakly for type I and X collagen (Fig. 8). Supplementation with 5% FBS prevented contraction of fibrin constructs (Fig 7A) and had no effect on total sGAG production (Fig. 7D). Due to the significant change in volume and therefore decrease in mass of hydrogels not supplemented with FBS, the sGAG content as a percentage of wet weight for this group was approximately four- to fivefold higher than hydrogels maintained in the presence of 5% FBS (Fig. 7C). Maintenance at 5% O2 increased sGAG production for both these groups (Fig. 7D).


Histological evaluation of agarose and fibrin hydrogels at day 42. Alcian blue staining for sulfated proteoglycan, picrosirius red for total collagen, and immunohistochemical staining for collagen types I, II, and X. Scale bar is 1 mm. FBS, fetal bovine serum. Color images available online at
Discussion
Identifying environmental conditions that enhance the proliferation and chondrogenic potential of MSCs is an essential prerequisite for tissue engineering and regenerative medicine therapies targeting damaged and diseased articular cartilage. In this study, it was found that expansion of diseased human FPSCs in the presence of FGF-2 increases the cell yield and enhances their chondrogenic potential. As evident by the lower number of colonies formed in a medium supplemented with FGF-2, and the subsequent enhanced sGAG synthesis rates of the progeny of these progenitors, it would appear that expansion in the presence of this growth factor appears to help select and/or expand a more proliferative and chondrogenic subpopulation from the heterogeneous cell population isolated from the IFP. Furthermore, chondrogenic differentiation of FPSCs expanded in the presence of FGF-2 was further enhanced by subsequent maintenance in a low-oxygen environment. Having identified appropriate expansion conditions, we sought to identify differentiation conditions that might facilitate the development of cartilaginous grafts engineered using human FPSCs. We hypothesized that recapitulating aspects of the in vivo regenerative environment known to promote chondrogenesis of MSCs, specifically fibrin deposition, a low-oxygen environment, and the presence of elevated levels of growth factors, would facilitate the development of grafts engineered using FPSCs. This was indeed the case with fibrin hydrogels found to better support the development of a cartilaginous tissue compared to agarose hydrogels using human FPSCs, demonstrating both the importance of the local substrate for tissue-engineering applications and for the potential utility of FPSCs for such strategies.
As has been observed for MSCs isolated from other tissues such as bone marrow, 55 some donor variability was also observed in the colony-forming efficiency of cells isolated from the IFP. The colony-forming efficiency was also affected by growth factor supplementation. The exact mechanism for this is unclear, although it suggests that FGF-2 may also play a role in preventing a subpopulation of otherwise colony-forming cells from either adhering or expanding in monolayer culture. Previous studies have also observed significant variability with human MSC populations in terms of proliferative capacity and subsequent multipotentiality, which do not appear to be a function of age or gender.56,57 Serum effects and/or laboratory procedure and protocols may also at least partially contribute to such variability. 13 The colony size was also affected by expansion conditions, with dramatically larger colonies observed in groups supplemented with FGF-2 at 5% oxygen tension. This larger colony size is indicative of more rapid cell proliferation, although cell-doubling times for FPSCs expanded in FGF-2 were not found to be statistically lower than standard expansion conditions when data were pooled from all donors (Fig. 3B). An alternative explanation for the smaller colony size observed in standard expansion conditions is that the higher number of colonies that were formed results in contact inhibition, thereby limiting cell proliferation. The cell-doubling time was significantly lower for FPSCs expanded in the presence of both FGF-2 and TGF-β3, demonstrating that these growth factors act synergistically to promote more rapid proliferation. The number of population doublings occurring before reaching confluence during passage 0 (the cell yield) was also higher for FPSCs supplemented with FGF-2 or FGF-2 and TGF-β3, which was possibly due to a smaller cell size as has been observed for bone marrow-derived MSCs expanded in the presence of FGF-2.13,26
In agreement with previous studies using human FPSCs, expansion in the presence of FGF-2 enhanced subsequent chondrogenesis in a pellet culture system. 24 The finding that low-oxygen conditions during pellet culture also improve chondrogenesis of FPSCs is also in agreement with the same authors. 50 The results of our study also suggest that these two factors can act synergistically to enhance chondrogenesis of FPSCs, with expansion in FGF-2 appearing to be a more potent prochondrogenic factor than differentiation at a low-oxygen tension. Importantly, for tissue-engineering applications, the benefits of differentiation in a low-oxygen environment were also observed after encapsulation into fibrin hydrogels.
Encapsulation of human FPSCs in agarose hydrogels has been shown to suppress their chondrogenic potential compared to pellet culture. 58 Given the well-documented advantages of hydrogel encapsulation for tissue-engineering applications, we sought to explore the use of alternative hydrogel substrates as a means to generate cartilaginous grafts. Fibrin hydrogel-based biomaterials offer high biocompatibility and provide great promise as a scaffold material for cartilage repair. 59 Indeed, fibrin hydrogels perhaps best mimic the body's own regenerative substrate, which under low-oxygen conditions facilities cartilage tissue formation. In this study, we observed improved chondrogenesis of human FPSCs in fibrin compared to agarose hydrogels, which was further enhanced under low-oxygen conditions. This provides support for our hypothesis that recapitulating aspects of the microfracture environment, namely the use of a fibrin substrate and a low-oxygen microenvironment, is a promising approach for cartilage tissue engineering. Of course, the microfracture technique generally leads to the formation of mechanically inferior fibrocartilaginous tissue compared to normal cartilage. This may be driven, at least in part, by the challenging inflammatory and mechanical environment within a damaged joint that can promote a more fibrous phenotype at the expense of a chondrogenic phenotype.60–62 In vitro studies have demonstrated the importance of biophysical cues in determining the fate of chondrogenic cells, 63 with tensile loading having been shown to promote type I collagen synthesis, 64 while alternative cues such as dynamic compression65–69 and hydrostatic pressure70–72 have been shown to enhance sGAG and type II collagen synthesis and suppress markers of hypertrophy.73,74 The results of computational models point to a complex mechanical environment in vivo with spatially varying signals that may drive MSC differentiation down undesired pathways. 60 An inherent advantage of in vitro tissue engineering is that these factors can be controlled using bioreactors, allowing engineered tissues to be mechanically primed before implantation.29,58,67,75–79 Inflammatory cytokines present in damaged or diseased joints may also alter MSC proliferation kinetics and potentially inhibit chondrogenesis. Various inflammatory cytokines have been shown to influence the proliferative potential of MSCs.80–82 Catabolic factors such as interleukin-1α and/or tumor necrosis factor-α have been shown to suppress chondrogenesis of MSCs.83–86 Understanding how oxygen, mechanics, various cytokines, and substrate interact to regulate cell fate is central to regenerating hyaline cartilage using MSCs.
At this stage, no definitive explanation can be provided for why fibrin provides a more chondrogenic environment than agarose hydrogels for human FPSCs. Somewhat surprisingly, we have recently observed the opposite result using immature porcine bone marrow-derived MSCs, where agarose better supported chondrogenesis compared to fibrin hydrogels. 74 The cell-mediated contraction observed in fibrin hydrogels, which in turn supports greater cell–cell interactions, is hypothesized as one potential mechanism for this improvement in chondrogenesis using diseased human FPSCs. Cells cannot directly adhere to agarose, thereby limiting contraction, migration, proliferation, and hence cell–cell interactions that appear critical for supporting chondrogenesis of adult MSCs. A potential limitation of the use of fibrin for tissue-engineering applications is its inherent capacity to undergo contraction when cells are cultured on it, rendering the final geometry of the engineered graft unpredictable. In this work, we demonstrated that supplementation of 5% FBS during culture inhibits contraction of these hydrogels. The exact mechanism by which FBS prevents this is unclear, although it may be due to FPSCs proliferating extensively on the surface of the fibrin hydrogels in the presence of serum, thereby providing a supporting cell-and-tissue layer that resists contraction.
In conclusion, the colony-forming efficiency and growth kinetics of human IFP-derived stem cells depend on the local oxygen tension and growth factors in which the cells are expanded. Expansion in the presence of FGF-2 appears to help select for, and/or preferentially expand, a subpopulation of the total mononuclear cell fraction within the IFP, the progeny of which are more chondrogenic than cells expanded in standard conditions. Recapitulating aspects of the in vivo regenerative environment, specifically encapsulation in fibrin and maintenance in a low-oxygen environment, facilitated the development of cartilaginous grafts engineered using FPSCs expanded in the presence of FGF-2. These results support the future use of FPSCs in combination with fibrin hydrogels as a therapy to treat damaged articular cartilage.
Footnotes
Acknowledgments
DJK is funded by the Science Foundation Ireland under the President of Ireland Young Researcher Award (08/Y15/B1336) and by a European Research Council Starter Award (StemRepair—Project number 258463).
Disclosure Statement
The authors have nothing to disclose.