
Abstract
Cartilage injury represents one of the most significant clinical conditions. Implantation of expanded autologous chondrocytes from noninjured compartments of the joint is a typical strategy for repairing cartilage. However, two-dimensional culture causes dedifferentiation of chondrocytes, making them functionally inferior for cartilage repair. We hypothesized that functional exclusion of dedifferentiated chondrocytes can be achieved by the selective mapping of collagen molecules deposited by chondrogenic cells in a three-dimensional environment. Freshly isolated and in vitro expanded human fetal or adult articular chondrocytes were cultured in a thermoreversible hydrogel at density of 1×107 cells/mL for 24 h. Chondrocytes were released from the gel, stained with antibodies against collagen type 2 (COL II) or COL I or COL X and sorted by fluorescence activated cell sorting. Imaging flow cytometry, immunohistochemistry, quantitative polymerase chain reaction, and glycosaminoglycan (GAG) assays were performed to evaluate the differences between COL II domain forming and COL II domain-negative cells. Freshly dissected periarticular chondrocytes robustly formed domains that consisted of the extracellular matrix surrounding cells in the hydrogel as a capsule clearly detectable by imaging flow cytometry (ImageStream) and confocal microscopy. These domains were almost exclusively formed by COL II. In contrast to that, a significant percentage of freshly isolated growth plate pre-hypertrophic and hyperdrophic chondrocytes deposited matrix domains positive for COL II, COL I, and COL X. The proportion of the cells producing COL II domains decreased with the increased passage of in vitro expanded periarticular fetal or adult articular chondrocytes. Sorted COL II domain forming cells deposited much higher levels of COL II and GAGs in pellet assays than COL II domain-negative cells. COL II domain forming cells expressed chondrogenic genes at higher levels than negative cells. We report a novel method that allows separation of functionally active chondrogenic cells, which deposit high levels of COL II from functionally inferior dedifferentiated cells or hypertrophic chondrocytes producing COL X. This approach may significantly improve current strategies used for cartilage repair.
Introduction
R
Cells are usually sorted or separated based on their differences in cell surface antigens or cytoplasmic density. Cellular differences in surface antigens (CD markers) enable cell sorting by fluorescent-activated cell sorting (FACS) or magnetic-activated cell sorting. Cellular differences in cytoplasmic density help to separate different cells by density gradient centrifugation. CD marker expression profile of cultured, dedifferentiated chondrocytes has been compared to freshly isolated chondrocytes by several groups.8,9 A group of CD markers such as CD49c, CD49f, and CD44 have been proposed to predict the chondrogenic capacity of monocultured chondrocytes. 10 However, all previously published studies are based on a complex combination of CD markers, which has an indirect, partial correlation with the chondrogenic phenotype. Currently, cell sorting technologies have not been based on the detection of ECM molecules produced by the cells.
In this study, we proposed a new method of cell sorting that we have termed extracellular matrix domain (EMD) detection (EMDD), which allows for the enrichment of functionally active, COL II-producing chondrocytes and the exclusion of dedifferentiated and hypertrophic cells expressing COL X. EMDD is based on a thermoreversible gel system that creates a 3D culture environment for chondrocytes and permits the isolation of cells without disruption of the ECM capsule produced around secretory active chondrocytes. The goal of this methodology is to select cells that possess a functional phenotype predisposed to COL II synthesis, the predominant constituent of hyaline cartilage, rather than COL I synthesis more typical of the fibrocartilage ECM.
Methods
Cell culture and expansion
Human fetal tissues were obtained from Novogenix Laboratories, LLC following informed consent and elective termination. All donated material was anonymous and carried no personal identifiers. The developmental age was determined by ultrasound. Periarticular chondrocytes or growth plate fetal chondrocytes were isolated under the dissection microscope from 17-week-old human fetuses. Adult articular chondrocytes were isolated from full thickness cartilage dissected from knee biopsies of a patient undergoing total knee replacement. Isolation of chondrocytes was carried out as previously described.11,12 In short, cartilage was digested for 20–22 h in collagenase type II (0.15%) in the Dulbecco's Modified Eagle Medium (DMEM) supplemented with penicillin (100 U/mL) and streptomycin (100 mg/mL). Chondrocytes were then cultured in the chondrocyte proliferation medium (DMEM/F12 supplemented with 10% fetal bovine serum and 100 U/mL penicillin and 100 μg/mL streptomycin). The medium was refreshed every 3–4 days. Upon confluence, chondrocytes were trypsinized. Half of the cells were cryopreserved; the other half was replated. All chondrocytes (fetal or adult) used in this article have experienced at least one cycle of freeze-and-thaw. All reagents used for cell culture were purchased from Invitrogen, unless otherwise stated. Common chemicals were purchased from Sigma-Aldrich.
3D culture in thermoreversible hydrogel or agarose
To assess the collagen expression signature of freshly isolated chondrogenic cells, either fetal periarticular, growth plate chondrocytes or adult articular chondrocytes were first resuspended in a thermoreversible hydrogel (Cosmo Bio [Tokyo, Japan]) on ice at a 1×107 cell/mL density (Fig. 1A). Next, hydrogels with cells were solidified by incubating at 37°C and cultured in the chondrogenic medium (the StemPro base medium with 50 μg/mL of ascorbic acid, 10 ng/mL of transforming growth factor-β3 (TGFβ3), and 100 U penicillin/mL and 100 μg/mL streptomycin) stimulating ECM production. Twenty-four hours later, hydrogels were liquefied by cooling plates on ice. Chondrocytes were released from hydrogels, washed with phosphate-buffered saline (PBS), and stained with antibodies against COL II, COL I, or COL X, respectively, or in combination (as indicated in figure legends), and analyzed by FACS. The entire procedure is summarized in Figure 1. To visualize ECM domains on histology sections, we cultured fetal periarticular chondrocytes (Passage 1) in an agarose gel for 7 days in the chondrogenic medium. The agarose hydrogel was used because of scientifically better quality of sections after paraffin embedding as compared to the thermoreversible hydrogel. Toluidine Blue staining was performed to show glycosaminoglycan (GAG) formation, and immunohistochemistry was carried out to indicate the expression of COL II.

Chondrocytes produce collagen-rich extracellular matrix (ECM) domains in hydrogels.
FACS and Imagestream analysis
Noncultured fetal periarticular and growth plate chondrocytes, adult articular chondrocytes or in vitro expanded periarticular (passage 1–6) chondrocytes were suspended in liquified thermoreversible hydrogel at a density of 10 million cells per mL on ice; 100 μL of gel was used for each sample. After the addition of cells and mixing, hydrogels were solidified by incubating at 37°C for 5 min. Hydrogel droplets with trapped cells were cultured in the chondrogenic medium for 24 h before sorting. After 24 h, culture plates were placed on ice, the hydrogel was liquefied, and recovered live cells incubated with rabbit antibodies against COL II, COL I, or COL X (Abcam) or the mouse antibody against CD29 followed by washing with PBS containing 1% bovine serum albumin (BSA). For dedifferentiation studies, cells were next incubated with secondary antibodies conjugated with Alexa-488 or −594 fluorochrome, and then washed with PBS containing 1% BSA. Control samples were processed without incubation with a primary antibody. For simultaneous detection of two collagens in matrix domains, primary antibodies were labeled with Zenon® Alexa 594 or Alexa488 IgG2a Labeling Kits (Invitrogen). Stained samples were sorted using a BD FACSAria or analyzed using the LSRII cytometer (BD Biosciences). Data files were exported and analyzed with FACSDiva software (BD Biosciences). The same cells were also analyzed using the ImageStream IS100 imaging flow cytometer (Amnis) and analyzed using IDEAS software (Amnis). Data were obtained from a minimum of 5000 cells. ImageStream is a combination of flow cytometry and fluorescence microscopy. On one hand, it can analyze a large number of cells based on their fluorescent labels, on the other hand, it also enables detailed morphometric analysis based on fluorescent images for individual cells. 13
RNA isolation and quantitative polymerase chain reaction
Gene expression analysis of cells in monolayer culture was performed as described.11,12 RNA samples were isolated with the RNeasy Micro Kit (Qiagen). For quantitative polymerase chain reaction (qPCR), 1 μg of total RNA was reverse transcribed into cDNA using the Omniscript RT kit (Qiagen). qPCR was performed on cDNA samples by using the Maxima SYBR Green/ROX qPCR Master Mix (Thermo Scientific). PCR reactions were carried out on ViiA™ 7 Real-Time PCR System (Life Technologies). For each reaction, a melting curve was generated to test primer dimer formation and nonspecific priming. The primers for real-time PCR are listed in Supplementary Table S1 (Supplementary Data are available online at
Pellet culture and chondrogenic differentiation
Passage 2 of fetal periarticular chondrocytes was sorted based on the expression of Col II domains. Fifty thousand sorted cells (either positive or negative) were seeded in one well of a round-bottom 96-well plate (nontissue culture treated). Cells were aggregated by centrifugation at 500 g for 5 min. Cell aggregates were cultured in the chondrogenic differentiation medium (the StemPro base medium supplemented with 50 μg/mL of ascorbic acid, 10 ng/mL of TGFβ3, and 100 U penicillin/mL and 100 μg/mL streptomycin) for 1 week. The medium was refreshed every other day.
Histology and immunohistochemistry
Three cell aggregates formed by COL II domain-positive or -negative chondrocytes were fixed in 10% formalin for 15 min, dehydrated with ethanol, and embedded in paraffin using routine procedures. A microtome (Leica) was used to cut 5-μm sections. Slides were then deparaffinized and stained for sulfated GAG with Toluidine Blue. For immunohistochemistry, paraffin sections were processed using routine techniques. Rabbit antibodies against COL I and COL II (both Abcam) were used, followed by incubation with horseradish peroxidase-conjugated secondary antibodies against rabbit immunoglobulin G (Vector Laboratories). Antibodies were then visualized with Peroxidase substrate Kit DAB (Vector Laboratories). Images were acquired using either the confocal TCS SP2 AOBS laser-scanning microscope system (Leica Microsystems, Inc.) with 63×(1.4 numerical aperture [NA]) oil-immersion objectives or using the Zeiss AxioVision software version 4.8 Carl Zeiss Microscope (Carl Zeiss) equipped with ApoTome.2: modules for Axio Imager.2 and Axio Observer with 10×, 20×, 40×(1.3 NA), and 63×(1.4 NA) oil-immersion objectives.
Quantitative GAG and DNA assay
Cell aggregates (n=3) were washed with PBS and stored at −80°C for 16–20 h. Subsequently, they were digested in a buffer (1 mg/mL proteinase K in the Tris/ethylenediaminetetraaceticacid (EDTA) buffer (pH 7.6) containing 18.5 μg/mL iodoacetamide and 1 μg/mL pepstatin A) for more than 16 h at 56°C. The GAG content was spectrophotometrically determined with 1,9-dimethylmethylene blue chloride staining in the phosphate buffer with ethylenediaminetetraacetic acid buffer (14.2 g/L Na2HPO4 and 3.72 g/L Na2EDTA, pH 6.5) using an FLUO star Omega microplate reader (BMG LABTECH) at an absorbance of 520 nm with chondroitin sulfate as a standard. Total DNA amounts in aggregates were determined by a CyQuant DNA Kit (Molecular Probes).
Statistical analysis
All experiments were repeated at least three times. GAG and DNA quantifications were examined for statistical significance with the paired Student's t test. For statistical analysis of qPCR data, the dCt for positive and negative cells were subject to a simple t-test. p Values of<0.05 were considered as statistically significant.
Results
Freshly isolated chondrocytes form ECM domains in thermoreversible hydrogel system
Freshly dissected fetal periarticular chondrocytes robustly formed domains that consisted of the ECM surrounding cells in the hydrogel as a capsule-like structure. These domains were almost exclusively formed by COL II (Fig. 2). In contrast to that, a significant percentage of freshly isolated growth plate chondrocytes deposited matrix domains positive for COL II, COL I, and COL X. This is consistent with the expression signature of hypertrophic cartilage cells (Fig. 1C).

Visualization of the ECM domains.
Decrease of COL II producing cells reflects the dedifferentiation of chondrocytes
Morphology of chondrocytes expanded in two-dimensional (2D) for up to six passages is shown in Supplementary Figure S1. From passage 1 to 6, chondrocytes turned into fibroblast-like cells with a flat and spindle-like appearance different from their original small, polygonal shape. The most dramatic change in morphology happened between passage 3 and passage 4. qPCR confirmed that the expression of chondrogenic genes dropped rapidly during the 2D expansion (Supplementary Fig. S2). In agreement with morphological findings, the level of tested chondrogenic genes also dropped quickly from passage 1 to passage 2. Less than 1% of expanded periarticular chondrocytes expressed COL X after six passages (not shown), and therefore, the next studies focussed on COL I and II. We next tested if dedifferentiation of chondrocytes correlates with the formation of COL II domains in hydrogel. Up to 90% of cells showed robust COL II-rich matrix domain formation at passage 0–1, but this potential progressively decreased with the passage number (Fig. 3A, B). As shown in Figure 3C, the ratio of fetal chondrocytes producing COL I-rich domains markedly increased (from 1–5% to 60–70%) after in vitro expansion for 6 passages (Fig. 3B). It is important to mention that fetal synovial or bone marrow-derived stromal cells obtained from the same fetuses produced no COL II or COL I matrix domains with a 24–48-h interval.
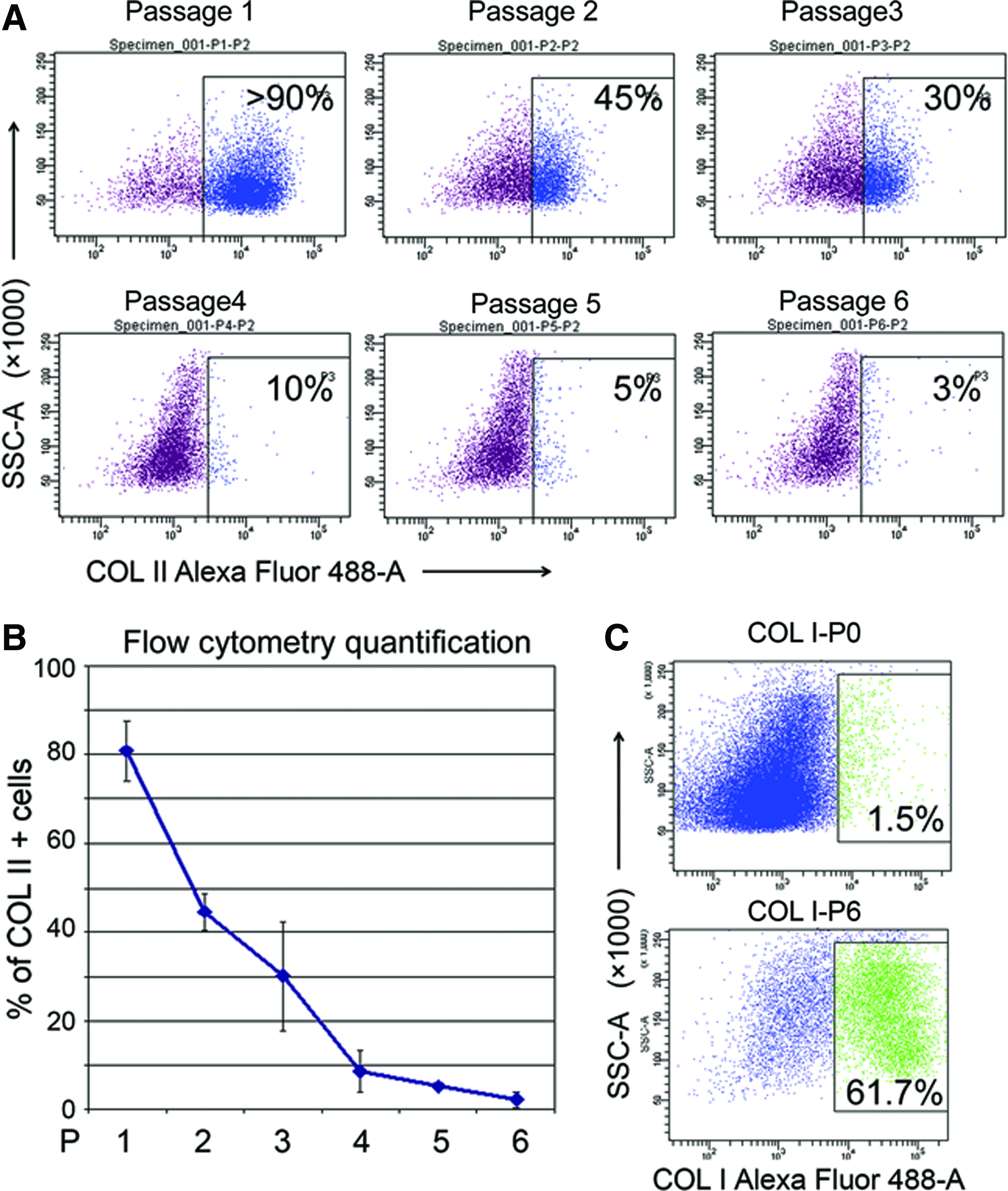
The ratio of COL II producing cells decreases as chondrocytes dedifferentiate.
Cultured adult articular chondrocytes also showed the ability to form EMD in the thermoreversible gel. However, only a small subset (∼10) of chondrocytes from adult articular cartilage deposit ECM domains can be detected by COL II antibody staining (Supplementary Fig. S3) after in vitro expansion (passage 3).
Chondrocytes that maintain their phenotype can be separated from dedifferentiated cells based on matrix domain
We next tested if heterogeneous in vitro expanded periarticular chondrocytes could be separated on the basis of their collagen deposition signature. First, to visualize ECM domains on histological sections, chondrocytes were cultured in an agarose gel for 7 days before histological assessment. COL II antibody-based labeling was used to detect ECM domains surrounding chondrocytes (Fig. 3A). GAGs were also deposited into ECM domains by some of the gel-trapped chondrocytes (Fig. 3A), indicating functional heterogeneity of the cultured cells. ImageStream analysis and high-resolution confocal microscopy were carried out next to confirm the specific morphological extracellular appearance of matrix domains produced by chondrocytes cultured in the thermoreversible hydrogel (Fig. 3B, C, respectively).
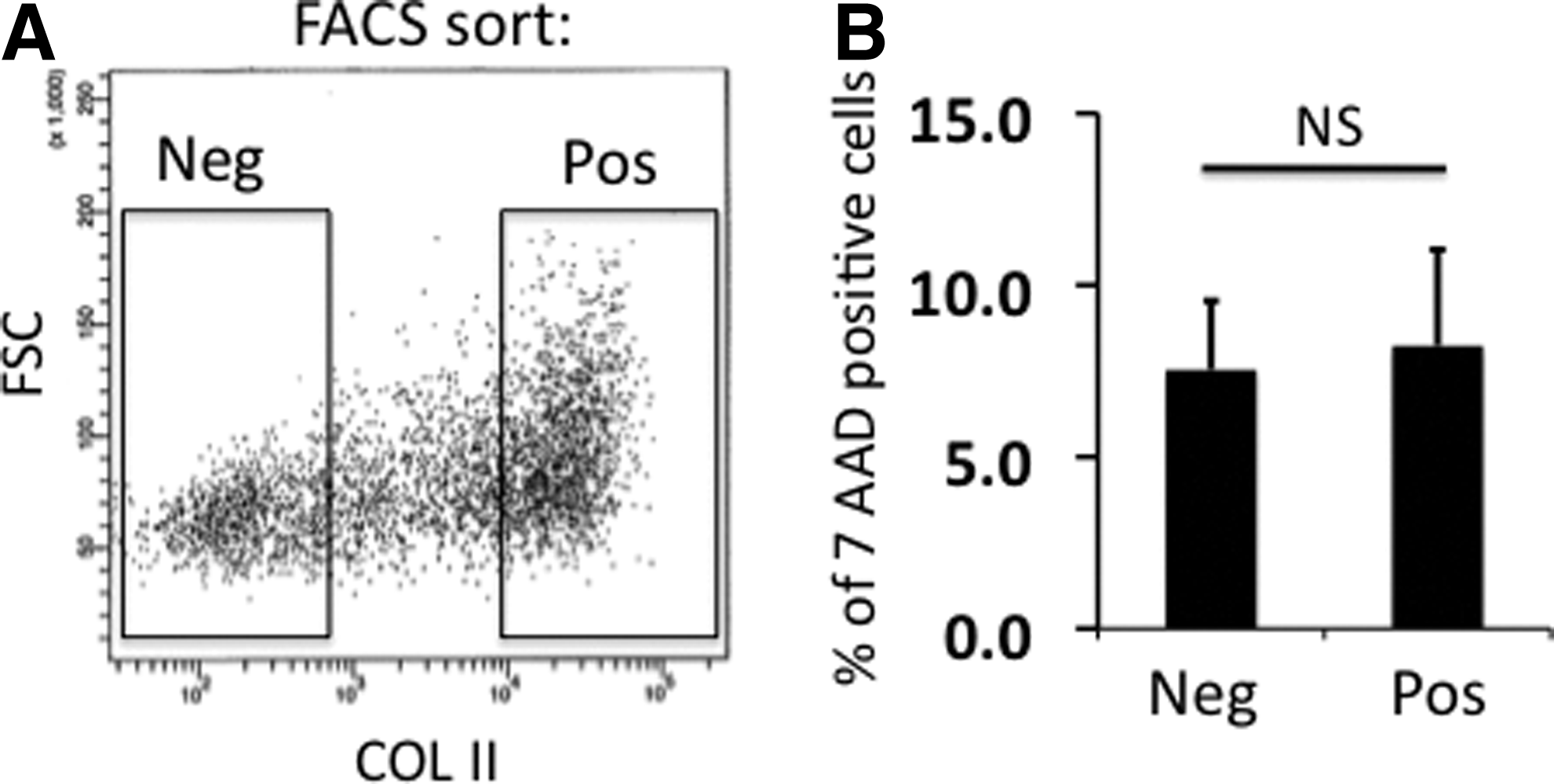
Next, FACS analysis was performed to sort COL II matrix domain producing chondrocytes after 2 passages in 2D culture (Fig. 4A). No difference in viability was observed between these two subsets as detected by the quantitative viability test (Fig. 4B). Isolated COL II domain-positive or COL II-negative cells were seeded to form aggregates, respectively (Fig. 5A). Immunohistochemical assessment of cell aggregates showed that COL II domain forming cells almost exclusively deposited COL II proteins, while COL II-negative cells produced COL I (Fig. 5A). qPCR data from 3D culture of sorted cells also confirmed higher expression of the chondrogenic genes in COL II domain-positive cells (Fig. 5B), while COL I expression was much higher in COL II domain-negative cells, indicating that they represent dedifferentiated chondrocytes (Fig. 5C). Although immunostaining of cell aggregates with Toluidine Blue showed a minimal difference in GAG deposition (Supplementary Fig. S4), quantitative GAG production analysis normalized to DNA levels indicated that COL II-positive cells produced more GAG than negative cells, further indicating that COL II domain forming cells are functionally superior (Fig. 5D). To further confirm the differences of aggregate sizes formed by sorted EMD-positive and -negative cells, we quantified the diameters of aggregates. As shown in Figure 5E, aggregates formed by positive cells were significantly bigger. Finally, the expression levels of COL II isoforms A and B were determined by qPCR in the EMD-positive and -negative populations sorted by FACS. Positive cells expressed significantly higher levels of both isoforms than negative cells (Fig. 5F). The same data with different calculations indicated that both positive and negative cells express more COL II isoform A than isoform B (Fig. 5G).
Functional sorting of COL II matrix domain producing chondrocytes.

Morphological and functional analysis of COL II domain- positive and COL II domain-negative chondrocytes. Fetal periarticular chondrocytes (passage 2) were sorted as shown in Figure 4(A) and seeded in 96-well plates at 50,000 cells per well density and cultured for 7 days.
Discussion
In this study, we used a thermoreversible hydrogel to permit ECM domain formation by cultured chondrocytes and then separated COL II producing cells by FACS. COL II domain-positive or -negative cells demonstrated very distinct characteristics in regard to morphology, histological appearance, and gene expression, suggesting that cells retaining their chondrogenic properties can be enriched based on their production of COL II. In particular, the data clearly demonstrate the superiority of sorting using COL II domain generation rather than GAG matrix domains since COL I-positive cells still produced substantial GAG after culture in aggregates under conditions suitable for re-expression, while failing to recover substantial COL II production. The phenotypically normal COL II-positive cells can be defined/enriched by sorting COL II producing cells that may be more phenotypically stable upon implantation.
This study demonstrates for the first time functional cell sorting based on ECM production of two different markers by multiplex sorting. Current techniques for cell sorting are generally based on principles that employ either separation based on the cytoplasmic density or surface antigen detection. Density gradient centrifugation has long been used for rough separation of cells, although it is only applicable for separation of very distinct cell populations with obvious differences in cellular density. 15 On the other hand, isolation of cells by CD markers is more broadly applicable. This principle is based on differences in cell surface antigen expression, which is used for immunophenotyping of the cells. CD markers have numerous physiological functions, often acting as receptors or ligands in specific cell signaling pathways. Some CD markers also function as adhesion molecules. Although cell sorting by CD markers has greatly advanced cell biology, the link between surface antigens and cell function is often indirect and empirical. Our study provides an important enhancement of current technologies used for cell sorting.
Previously, studies indicated that some articular chondrocytes undergo hypertrophy and deposit COL X in long-term cultures. 7 Although no significant deposition of COLX by cultured periarticular chondrocytes was observed in this study, detection of COLX depositing cell subset was clearly observed after plating freshly isolated hypertrophic chondrocytes in hydrogels. Detection and labeling of the COLX depositing cells offer a unique tool for the isolation of highly purified hypertrophic chondrocytes.
Matrix-induced autologous chondrocyte implantation (MACI) is a very popular treatment for cartilage defects. 16 Like the classical procedure of ACI, it includes arthroscopic excision of biopsies from low-weight-bearing areas of healthy cartilage, isolation and expansion of chondrocytes in the laboratory, and implantation of chondrocytes into the cartilage defect. However, instead of injection into defects as a cell suspension, chondrocytes were seeded on a bilayer of porcine-derived type I/type III collagen after in vitro expansion. One major reason that this procedure may fail is that dedifferentiated chondrocytes are introduced into the defects together with cartilage matrix-producing cells. Our method can be used to exclude dedifferentiated cells by sorting out chondrocytes producing COL II domains. With enrichment of COL II producing cells that are implanted into cartilage defects, the efficacy of ACI and MACI could potentially be improved. EMDD can be used as a sensitive readout for factors that regulate matrix protein production in chondrocytes and other cell types. Similar to other antibodies previously used in clinical practice, antibodies against COL II can be manufactured in good manufacturing practice (GMP) facilities. Monoclonal antibodies have been approved by FDA for many therapeutic applications, especially for oncology. 17 To improve the performance of ACI or MACI, in vitro expanded chondrocytes can be sorted using GMP grade antibodies against COL II, or potentially, other ECM (COL II, I, VI, aggrecan, etc.) and implanted to patients.
Several significant improvements in cartilage tissue engineering have been reported recently. 16 Coculture of chondrocytes with mesenchymal stromal cells, mechanical stimulation of chondrogenic cultures, and control of metabolic parameters allowed producing cartilage tissue with better mechanical and biological characteristics with relatively small chondrocyte numbers.18–21 In this study, we report a novel method that allows enrichment for functionally active chondrogenic cells, which may further advance existing cartilage tissue-engineering strategies.
It is important to mention that EMDD can be not only used to sort chondrocytes, but in principle, any cell type that produces ECM in hydrogels. Based on the direct link between ECM production and biological processes such as inflammation, tissue repair, and metastatic cancer invasion, EMDD provides a useful tool to study cell behaviors in such biological events.
In conclusion, our data show that live chondrogenic cells can be separated on the basis of their ECM production using functional cell sorting based on the methodology we called EMDD. This approach can be carried out in a xenogen-free environment and allows for functional enrichment of chondrogenic cells producing high levels of COL II and may lead to significant improvement of current therapeutic approaches in the area of cartilage restoration.
Footnotes
Acknowledgments
The authors are grateful to Mrs. Ann George from Childrens Hospital Los Angeles for technical support on Imagestream analysis and to Ms. Felicia Codrea and Jessica Scholes from UCLA FACS Core for their help with sample analysis and sorting.
Funding
This project is funded by the NIH NIAMS K01AR061415 grant to D.E. and NIH/National Center for Advancing Translational Science (NCATS) UCLA CTSI Grant Number UL1TR000124.
Disclosure Statement
The authors declare no financial support or any other benefits from commercial sources received for the work reported on in the manuscript. D.E. and L.W. have submitted a patent disclosure to the UCLA office of intellectual property.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.