
Abstract
Large quantities of human pluripotent stem cells (hPSCs) needed for therapeutic applications can be grown in scalable suspended microcarrier cultures. These microcarriers are coated with animal or human extracellular matrix (ECM) proteins to promote cell growth and maintain pluripotency. However, the coating is costly for large-scale cultures and it presents safety risks. This study demonstrates that hPSCs can be propagated on noncoated positively charged cellulose microcarriers in a serum-free medium containing the ROCK inhibitor, (Y27632) or myosin inhibitor, Blebbistatin. In the presence of these two inhibitors, myosin phosphatase 1 and myosin light chain 2 were dephosphorylated suggesting that reduced myosin contractility is responsible for hPSC survival and growth on ECM coating-free microcarriers. Cells propagated on the noncoated microcarriers for 12 passages maintained their pluripotency and karyotype stability. Scalability was demonstrated by achieving a cell concentration of 2.3×106 cells/mL with 11.5-fold expansion (HES-3) in a 100-mL spinner flask. The differentiation capability of these cells toward three primary lineages is demonstrated via in vitro embryoid bodies and in vivo teratoma formations. Moreover, the directed differentiation to polysialylated neuronal cell adhesion molecule-positive (PSA-NCAM+) neural progenitors produced high cell concentrations (9.1±1.2×106 cells/mL) with a cell yield of 412±77 neural progenitor cells per seeded HES-3 and a PSA-NCAM expression level of 91±1.1%. This defined serum- and coating-free scalable microcarrier culturing system is a safer and less expensive method for generating large amounts of hPSCs for cell therapies.
Introduction
T
hPSCs are traditionally cultured on feeder cells (e.g., mouse embryonic fibroblasts), which provide a microenvironment for maintaining pluripotency. Over the years, hESC cultures have developed from coculturing with feeder layers to feeder-free Matrigel-coated cultures.4,5 However, Matrigel consists of undefined mouse-derived components, which cannot be used in regenerative medicine due to safety issues. With this in mind, research groups have been striving to establish new methods of culturing hPSCs in chemically defined and xeno-free conditions. Matrigel was replaced by defined extracellular matrix (ECM) proteins like mouse-derived
6
or human recombinant laminin,7,8 human plasma-derived
9
or recombinant vitronectin,
10
and ECM (bone sialoprotein, vitronectin)-derived peptides.
11
Interestingly, a study by Harb et al.
12
reported that the addition of Rho-associated kinase (ROCK) inhibitor, Y27632, into the serum-free medium (mTeSR1) enables the cultivation of hESC lines H1, BGN1, and H9 on poly-
Two-dimensional (2D) tissue culture platforms have been used for generation of hPSCs. However, in clinical applications, where the cell dosage as high as 5×109 cardiomyocytes is required to treat myocardial infarction, 15 mass production using the 2D platform is impractical. 16 To meet this requirement, microcarrier technology for hPSC expansion provides a large surface area for cells to grow in scalable, suspension bioreactors. hPSCs cultured on microcarriers grow as cell–microcarrier aggregates and not as monolayers. In the initial step of seeding, cells attach and spread on the microcarrier surface; thereafter, several microcarriers join together generating aggregates composed of microcarriers and cells. 17 Several research groups have demonstrated the feasibility of using this platform for culturing hPSCs in suspended cultures using ECM protein-coated microcarriers.17–23 The microcarrier platform is proven to be superior to the 2D tissue culture plates in terms of cell growth, robustness, and scalability without compromising pluripotency and karyotype stability of hPSCs. hPSCs cultured on microcarriers yielded between 2.2×106 and 6.1×106 cells/mL in stirred bioreactors with 15–20-fold expansion.17,19–21 These levels are significantly higher than hPSCs cultured on 2D tissue culture plates.19,21,24 Our group has developed a cylindrical microcarrier platform based on cellulose matrix derivatized with diethylaminoethyl (DEAE) tertiary amine positively charged groups (DE-53, Whatman).17,19 The platform enables the expansion of hPSCs to high cell densities (up to 6.1×106 cells/mL) as well as in situ neural differentiation sequentially. 21 Large quantities of polysialylated neuronal cell adhesion molecule-positive (PSA-NCAM+) neural progenitor cells (NPCs) of 8.4×106 cells/mL were generated with yields of 333–371 NPCs per seeded hPSC. 21
Similar to the 2D culture platform, Matrigel coating on microcarriers is essential for supporting long-term hPSC growth, while maintaining pluripotency.17,19 hPSCs cultured on Matrigel-coated microcarriers exhibited 1.9–18 times higher cell concentrations and higher pluripotency than noncoated ones. 19 Chen et al. examined seven different ECM proteins and identified laminin as a suitable candidate to replace Matrigel, for microcarrier coating. 19 A later study by Heng et al. demonstrated that laminin- or vitronectin-coated microcarriers can support hPSC propagation in the serum-free medium. 22 Both laminin- and vitronectin-coated microcarrier cultures exhibited similar cell growth profiles, consistently high expression (>90%) of pluripotent markers, maintenance of karyotypic normality, and retained the ability to differentiate into lineages of all three germ layers. 22
Using ECM proteins for microcarrier coating can be problematic since they may contain animal-derived components (Matrigel or mouse laminin) and are expensive (recombinant or plasma-purified human vitronectin, or recombinant human laminin (511, 521). In this study, we demonstrate for the first time that by adding either the ROCK inhibitor (Y27632) or Blebbistatin, hPSCs can be cultivated on uncoated positively charged DEAE cellulose microcarriers (DE-53) in a serum-free medium. Both hESCs and hiPSCs can be propagated in this system for multiple passages exhibiting high cell concentrations, while maintaining high levels of pluripotent markers and stable karyotypes. Differentiation capability was demonstrated by directed differentiation to NPCs. Scalability of the culture was demonstrated in 100-mL spinner flasks. We observed that dephosphorylation of myosin phosphatase 1 and myosin light chain 2 occurred in the presence of these two inhibitors, suggesting that reduced myosin contractility is responsible for hPSC growth on noncoated microcarriers.
This defined ECM-free, scalable microcarrier culturing system can serve as a safe and less expensive method for generating large quantities of hPSCs for clinical therapies.
Materials and Methods
Cell culture
The human embryonic stem cell line HES-3 (ES Cell International) and H7 (WiCell Research Institute, Inc.) and hiPSCs (IMR90; kindly provided by James Thomson 25 ) were used in this study. All cultures were maintained on Matrigel (BD Bioscience)-coated tissue culture plates (Becton Dickson Lab ware). The serum-free medium (SFM), STEMPRO® hESC SFM (Life Technologies) was used for propagation of HES-3, whereas hESC H7 and hiPSCs (IMR90) were maintained on mTeSR™1 (STEMCELL Technologies). Details on the coating procedure and subculturing are described in a previous publication. 26
Cultivation of hPSCs on microcarriers
Positively charged cellulose-based cylindrical microcarriers (DE-53; Whatman) were used in this study. Details on Matrigel coating of the microcarriers and cultivation of hPSCs on microcarriers in static and spinner cultures were described previously.19,21,27 Briefly, for microcarrier coating, 5 mg (4.2×104 particles/mg and 6.8 cm2/mg) of microcarriers were mixed with 1 mL of a diluted Matrigel solution (1:30) in an ice-cool knockout (KO) medium and incubated overnight in 4°C on an orbital shaker. 19 For stationary cultures, hPSC cultures were enzymatically harvested from confluent Matrigel-coated tissue culture plates (Accutase; PAA Laboratories) for HES-3 or 1 mg/mL Dispase (STEMCELL Technologies) for H7 and hiPSCs (IMR90). Accutase-treated cells generate single-cell suspension, whereas Dispase treatment resulted in small cell aggregates (Ø 200–300 μm). These cells were seeded on 4 mg/mL microcarriers (Matrigel-coated or noncoated) in six-well plates (Ultra-Low-attachment plate; Corning) at 2×105 cells/mL (7.35×103 cells/cm2 and 1.2 cells/particle). The plates were placed on an orbital shaker in a 37°C humidified incubator for 2 h at 110 rpm to support cell attachment to microcarriers. Afterward, they were kept in a static condition for 7 days. Subculturing of microcarrier culture was done via gentle mechanical breakage of microcarrier–cell aggregates into uniform size (Ø 200–300 μm) with repetitive pipetting. 27
The scalability of microcarrier culture was demonstrated using 100-mL spinner flasks (Bellco Cat. No. 1965-00100). The mechanically dissociated static microcarrier culture was seeded at viable cell concentrations of 4×105 cells/mL to the spinner flask having 200 mg of DE-53 microcarriers in a 25 mL medium. After seeding, the spinner flask culture was agitated at 25 rpm for 2 h. After 1 day in static condition, the culture volume was increased to 50 mL as the final working volume and continuous agitation was applied at 25 rpm.
For obtaining single-cell suspension, 5 mL of the cell–microcarrier aggregate culture was washed two times with phosphate-buffered saline (PBS) by decantation. Then, 0.5 mL of TrypLE™ Express (Life Technologies) was added. After 5 min of incubation with occasional mixing, 3 mL of PBS was added and cells were gently dislodged from the microcarriers by pipetting. The single cells were separated from microcarriers via sieving through a 40-μm cell strainer (BD Biosciences). Cell viability of more than 90% and harvesting efficiency of more than 70% were achieved.
For both stationary and spinner culture, 80% of the medium was exchanged daily by letting the aggregates settle and removing the media. Viable cell counts were determined using NucleoCounter (NC-100; ChemoMetec A/S). ROCK inhibitor, Y27632 (Calbiochem) and Blebbistatin (Sigma-Aldrich) were added daily at the specified concentrations to the noncoated microcarrier cultures.
Analysis of pluripotency
To evaluate the expression level of pluripotent markers (Oct4, Tra-1-60, and mAb8428), hPSCs on microcarriers were dissociated into single cells as described above. Thereafter, cells were immunostained and analyzed for expression of pluripotent markers using flow cytometry as described previously.17,19 The mAb84 antibody recognizes an extracellular domain Podocalyxin-like protein 1, which is expressed only in undifferentiated hPSCs. 28
For immunostaining, microcarrier–cell aggregates were plated on a Matrigel-coated tissue culture dish for 1 day to allow cell attachment. After fixing with 4% paraformaldehyde, they were stained with 4′,6-diamidino-2-phenylindole and mouse primary antibodies for Oct4. Alexa-fluor® 488-conjugated F(ab′)2 fragments of goat anti-mouse immunoglobulin G (Life Technologies) were used as the secondary antibodies. Immunoflurescence was visualized using a fluorescent microscope (IX-71; Olympus).
hPSC differentiation
For spontaneous differentiation, cell–microcarrier aggregates were allowed to form embryoid bodies in differentiation media according to Chen et al. 19 RNA from the differentiated cells was extracted and analyzed for pluripotent (Oct4 and Nanog), mesoderm (Nkx2.5 and Hand1), endoderm (AFP and GATA6), and ectoderm (Nestin and Pax6) marker expression. Detailed protocol and list of primers can be found in previous studies.19,21 Immunostaining of the embryoid bodies with α-smooth muscle actin (SMA, mesoderm), β-III Tubulin (ectoderm), and alpha-fetoprotein (AFP, endoderm) antibodies was described earlier by Chan et al. 29
For teratoma formation, hPSC–microcarrier cultures were mechanically dissociated by pipetting and seeded on Matrigel-coated tissue culture plates in either STEMPRO® hESC SFM or mTeSR™1 depending on hPSC lines. After 7 days, 4–5 million cells were harvested mechanically using STEMPRO® EZPassage™ Tool (Life Technologies) and injected into SCID mice as described in Choo et al. 30 The dissected tumor was embedded in paraffin, sectioned, and stained with hematoxylin–eosin (H&E) for histological analysis.
Directed differentiation of HES-3 on microcarriers to NPCs was carried out according to Bardy et al. 21 Briefly, 107 HES-3 cells expanded in spinner stirred cultures on ROCK inhibitor-treated noncoated microcarriers or Matrigel-coated microcarriers were transferred to another 100-mL spinner flask containing additional 4 mg/mL of DE-53 microcarriers and 50 mL of the KO medium (without addition of ROCK inhibitor or Blebbistatin). The cultures were differentiated for 20 days in stirring conditions (25 rpm), During this period, Noggin treatment and medium exchanges were done as described by Bardy et al. 21 Determination of PSA-NCAM expression was done by flow cytometry according to Bardy et al. 21
Western blot analysis
Protein extraction from 2D and microcarrier–hPSC cultures was done using a cell lysis buffer (Cell Signaling Technology Cat. no. 9803S) supplemented with 1 mM phenylmethanesulfonylfluoride, 1×PhosSTOP (Roche) and 1×protease inhibitor cocktail (Calbiochem Cat. no. 539134) according to the manufacturer's protocol. For microcarrier cultures, cell lysis was carried out in situ without enzymatic cell dissociation from microcarriers. Soluble proteins were separated from microcarriers and cell debris via centrifugation (13,000 rpm) at 4°C. Western blots were carried out as described previously31,32 using rabbit anti-MLC2 (Cell Signaling Technology), mouse anti-pMLC2 (Ser19; Cell Signaling Technology), rabbit anti-MYPT1 (Millipore) and rabbit anti-pMYPT1 (Thr 696; Millipore) antibodies. The anti-actin antibody was used as a loading control. The protein expression level was quantified by measuring the intensity of the band using ImageJ 1.43u. Lysates of HEK293 cultures were used as the positive control, which was recommended by the antibody provider (Cell Signaling Technology).
Karyotype analysis
hPSCs cultured on microcarriers were plated on Matrigel-coated 6-cm tissue culture plate for 5 days in SFM. The harvested cultures were sent for karyotype analysis as described in Chen et al. 19 Conventional cytogenetic analysis was done using the standard G-banding technique and 20 metaphases were analyzed.
Statistical analysis
Paired and unpaired Student's t-tests were used for statistical analysis according to experimental conditions. Error bars in the graphs represent standard errors calculated from three or more replicates. Asterisks indicate statistical significance of p-value<0.05.
Results
ROCK inhibitor enables the cultivation of hPSCs on positively charged microcarriers without ECM protein coating
The ECM protein coating on growth surface is essential for sustainable cultivation of hPSCs in 2D culture as well as suspended microcarrier culture.19,21,22 In a previous publication by Chen et al., 19 it was shown that ECM (Matrigel or laminin) coatings on a variety of microcarriers, including DE-53, are essential for hESC growth. Cell concentrations of 0.8–1.1×106 cells/mL with high expression levels of pluripotent markers were obtained during the long-term cultivation on ECM-coated microcarriers. 19 However, on the noncoated microcarriers, low cell growth with rapidly decreased expression of pluripotent markers was observed. 19
Attempts to generate aggregate culture without microcarriers was unsuccessful; HES-3 cells seeded at a concentration of 2×105 cells/mL (with or without ROCK inhibitor at a static or agitated condition) did not show any significant cell expansion over 7 days, emphasizing the need for attachment to microcarriers for initiation of cell growth (data not shown).
Thus, to achieve growth in cell–microcarrier aggregate cultures without addition of the ECM, we tested the possibility of propagating hESCs on noncoated microcarriers in the presence of the ROCK inhibitor. HES-3 were seeded at a cell density of 2×105 cells/mL on 4 mg/mL noncoated-positive charged DE-53 microcarriers in StemPro hESC SFM containing 0.25–10 μM of the ROCK inhibitor, Y27632. Cell growth and pluripotency were monitored for five passages. Matrigel-coated and noncoated DE-53 microcarrier (without ROCK inhibitor) cultures were used as positive and negative controls. In the negative control of noncoated DE-53 microcarriers, hESC cultures without ROCK inhibitor obtained low cell growth (Fig. 1A) and viable culture was maintained for only two passages (data not shown).
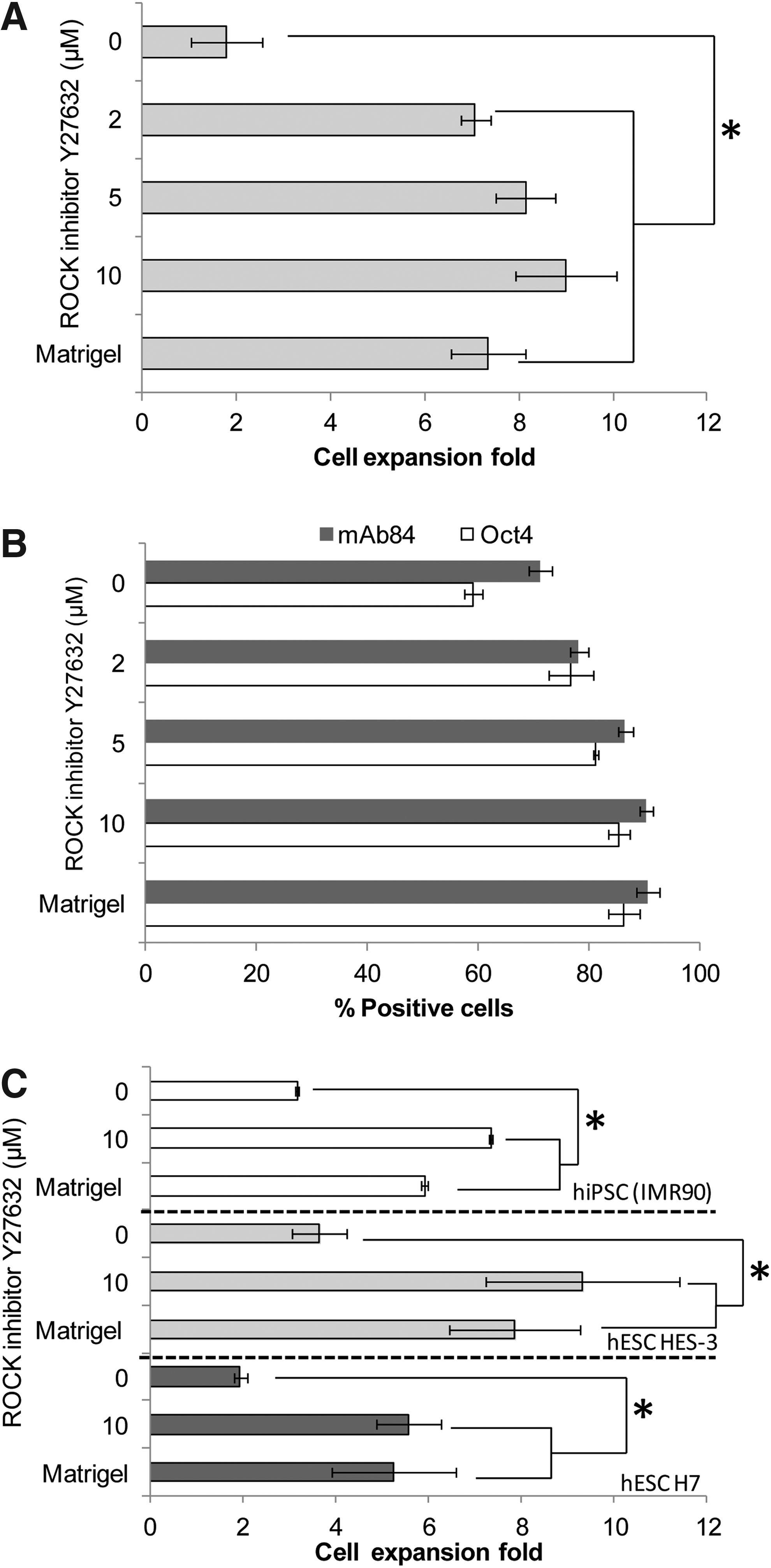
ROCK inhibitor supports human pluripotent stem cell (hPSC) growth on noncoated microcarriers. Cells were seeded at a concentration of 2.0×105 cells/mL on noncoated DE-53 microcarriers and cultured for five consecutive passages in a static condition. Matrigel-coated microcarrier cultures were used as the positive control without ROCK inhibitor treatment.
In the presence of ROCK inhibitor (2–10 μM), cells were able to attach to DE-53 microcarriers, and expanded for at least sevenfold, while maintaining steady cell growth of 1.4–1.8×106 cells/mL and high levels of pluripotency (Oct4>77% and mAb84>78%). Cultures with ROCK inhibitor at concentrations of 5 and 10 μM showed comparable expression levels of pluripotency markers as Matrigel-coated microcarrier cultures (Fig. 1B, p-value=0.21). Short-term treatment with ROCK inhibitor (10 μM, 1–2 days after cell seeding) of HES-3 DE-53 microcarriers (noncoated) resulted in a decrease in pluripotency markers (Oct4 56–71%, mAb84 41–48%, and Tra-1-60 61–69% after two passages). ROCK inhibitor-treated Matrigel-coated DE-53 microcarrier cultures achieved similar levels of cell growth and expression of the pluripotent marker (Tra-1-60) as nontreated ones (Supplementary Fig. S1; Supplementary Data are available online at
To verify the long-term robustness of the ROCK inhibitor-based microcarrier culture, HES-3, H7, and hiPS (IMR90) cells were propagated for 12–15 passages. All cultures achieved average cell concentrations of 1.29×106 cells/mL and high expression levels of pluripotent markers, which were comparable to Matrigel-coated microcarrier cultures (p-value>0.05) (Fig. 2). hPSC microcarrier culture treated with ROCK inhibitor exhibited spherical shape cell–microcarrier aggregates, whereas Matrigel-coated microcarrier culture has more irregular structures (Fig. 3A, B). The cell–microcarrier aggregates were composed of 1–10 hESC layers separated by microcarriers (H&E staining, data not shown). This structure suggests that the formation of the aggregates was initiated by cell attachment on microcarriers (cell attachment of 95±1% was achieved in 15 min,) followed by joining of several small aggregates to form larger ones. After 7 days, cell–microcarrier aggregates had reached an average diameter of 520±240 μm.

Long-term cultivation of three hPSC lines on ROCK inhibitor-treated noncoated microcarrier cultures as compared to Matrigel-coated ones. In each passage, 2×105 cells/mL were seeded on 4 mg/mL fresh microcarriers for 7 days. Cell concentrations and pluripotent markers were measured on day 7. Error bars indicate standard errors.
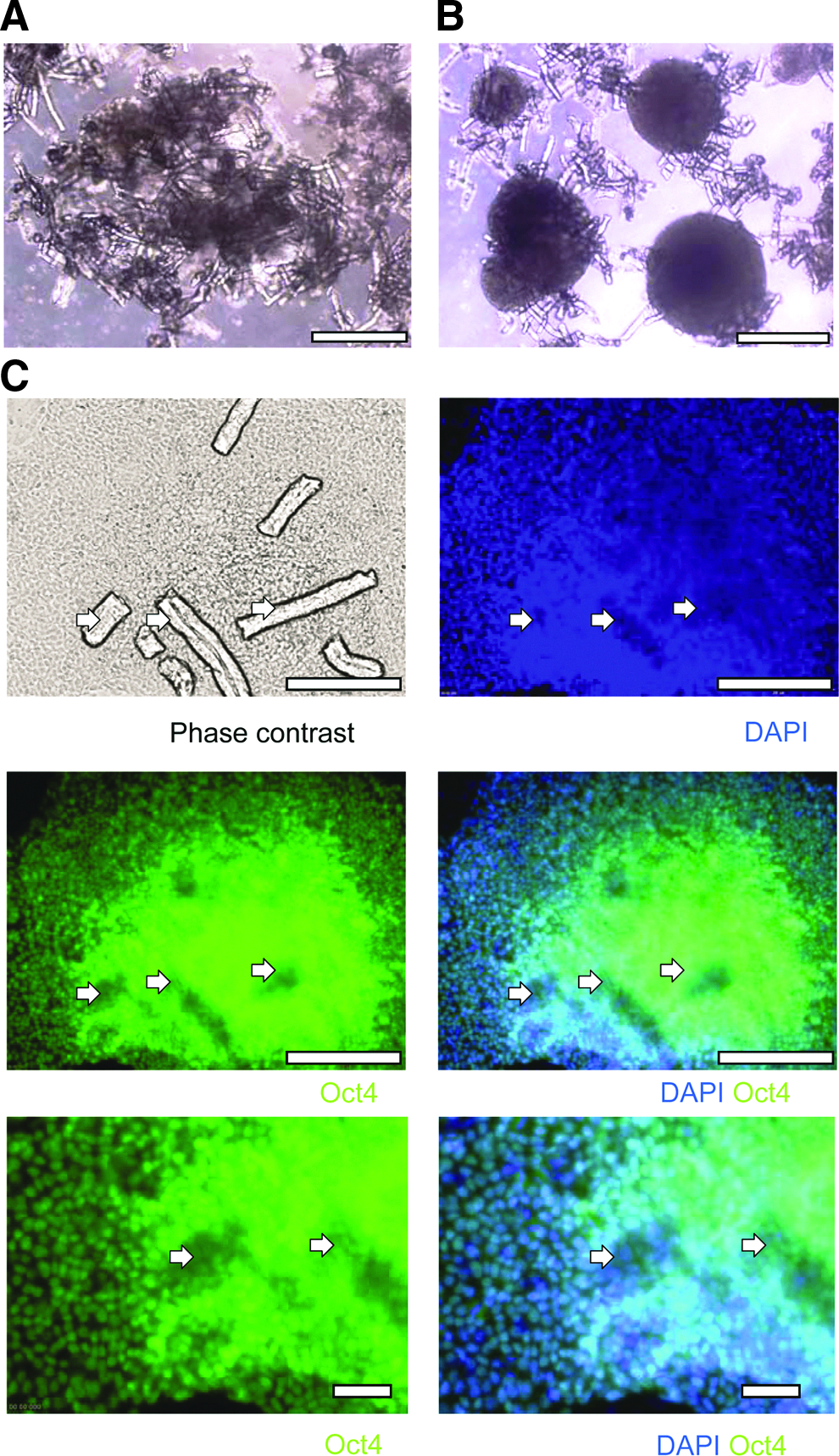
Immunostaining and morphology of ROCK inhibitor-treated cell–microcarrier aggregates obtained from static cultures (7 days). Phase-contrast image of HES-3 cells cultured on
The different morphology of the aggregates can be attributed to the inhibition of ROCK, which is known to be involved in regulating the shape and movement of cells by acting on stress fibers. 33 Immunostaining of pluripotency markers of the ROCK inhibitor-treated noncoated HES-3 microcarrier–cell aggregates show positive expression of the pluripotent marker Oct4 (Fig. 3C). The karyotype of HES-3 cells treated with the ROCK inhibitor remained normal for at least 15 passages (Supplementary Fig S2). Similar results were obtained using hESC (H7) and hiPSC (IMR90) lines demonstrating the universality of the system (Supplementary Fig. S2).
Inhibition of myosin II is associated with hPSC growth on noncoated microcarriers
ROCK affects several downstream targets of Rho and is mainly involved in regulating the shape and motility of cells by acting on the actin cytoskeleton regulation. 33 The formation of stress fibers is generated by phosphorylation of myosin light chain 2 (MLC2) that is induced by ROCK. Due to this phosphorylation, the actin binding of myosin II is enhanced and initiates the contraction of the cytoskeleton actin fibers. 33 To verify that cell growth on noncoated microcarriers is related to inhibition of myosin, we tested Blebbistatin, which directly inhibits myosin II heavy chain,13,34,35 for supporting hPSC growth on noncoated microcarriers. Phosphorylation levels of MLC2 and myosin phosphatase (MYPT1, which is located upstream to MLC2) were measured in ROCK inhibitor- and Blebbistatin-treated microcarrier cultures and compared to untreated cultures (Matrigel coated).
HES-3 cells were seeded on noncoated microcarriers in STEMPRO® hESC SFM in the presence of 10 μM of Blebbistatin in parallel to control cultures that contain 10 μM of ROCK inhibitor. Cell growth was maintained in both systems (12 passages) achieving an average cell concentration of 1.06±0.10×106 and 1.32±0.11×106 cells/mL for Blebbistatin- and ROCK inhibitor-treated cultures, respectively (Fig. 4A). Expression levels of pluripotent markers, Oct4, mAb84, and Tra-1-60, were similar in both microcarrier cultures (p-value>0.05) (Fig. 4A).

ROCK inhibitor and Blebbistatin treatment decrease phosphorylation of MYPT1 and MLC2 and enable human embryonic stem cell (hESC) growth on noncoated microcarriers.
We next analyzed the effect of ROCK inhibitor and Blebbistatin on the phosphorylation of MYPT1 and MLC2 in microcarrier cultures. Western blot analysis show that hESCs (HES-3 and H7) propagated on noncoated microcarriers in the presence of ROCK inhibitor or Blebbistatin (Fig. 4) exhibited lower levels of phosphorylated MYPT1 (pMYPT1) and MLC2 (pMLC2) when compared to Matrigel-coated microcarrier cultures. Thus, it appears that the reduction in actin–myosin contractility via ROCK inhibitor or Blebbistatin treatments lead to the survival and active growth of hPSCs on noncoated microcarriers.
Expansion of hPSCs in ROCK inhibitor-treated stirred spinner flask microcarrier culture
To demonstrate the scalability of the ROCK inhibitor-treated noncoated microcarrier culturing system, we propagated HES-3 and H7 cells on Matrigel-coated and noncoated DE-53 microcarrier spinner cultures and compared their cell growth and pluripotency. Matrigel-coated and ROCK inhibitor-treated microcarrier cultures of both hESC lines exhibited similar growth profiles (Fig. 5A). After 1 day lag phase, HES-3 cells grow exponentially showing similar doubling times of 27.0 and 27.7 h and maximal viable cell density of 2.3×106 and 2.5×106 cells/mL (p-value=0.075>0.05) for ROCK inhibitor-treated and Matrigel-coated microcarrier cultures, respectively. H7 cells exhibited also similar doubling time and maximal cell density (Matrigel-coated microcarrier: 32.5 h and 3.33×106 cells/mL; ROCK inhibitor-treated microcarrier culture: 32.2 h and 3.36×106 cells/mL). ROCK inhibitor-treated hPSC microcarrier cultures (HES-3 and H7) appeared to have higher expression levels of pluripotent markers (although not significant) than Matrigel-coated nontreated ones (Oct4 67–94% vs. 42–80%, p-value=0.34), mAb84 (91–98% vs. 63–80%, p-value=0.04), and Tra-1-60 (91–98% vs. 59–88%, p-value=0.12). Moreover, a minor peak adjacent to the negative control appears only in the flow cytometry histograms of the Matrigel-coated microcarrier culture indicating the presence of a small population of differentiating cells in this culture (Fig. 5B). These results can imply that ROCK inhibitor-treated hPSCs undergo structural cytoskeleton changes, which may have reduced the propensity of the cells to differentiate spontaneously, induced by fluidic shear stress. 36 Further work is needed to substantiate this assumption. Similar to static microcarrier cultures, ROCK inhibitor-treated spinner flask culture exhibited more spherical cell–microcarrier aggregates than those in Matrigel-coated spinner flask culture. In both cultures, the cylindrical DE-53 microcarriers were embedded in the aggregates between the cell layers (Fig. 5C, D). Cells harvested from these cultures exhibited high levels of viability (>90%) and H&E staining of the sectioned cell–microcarrier aggregates showed intact cells at the periphery and center of the aggregates without visible necrosis (Data not shown).
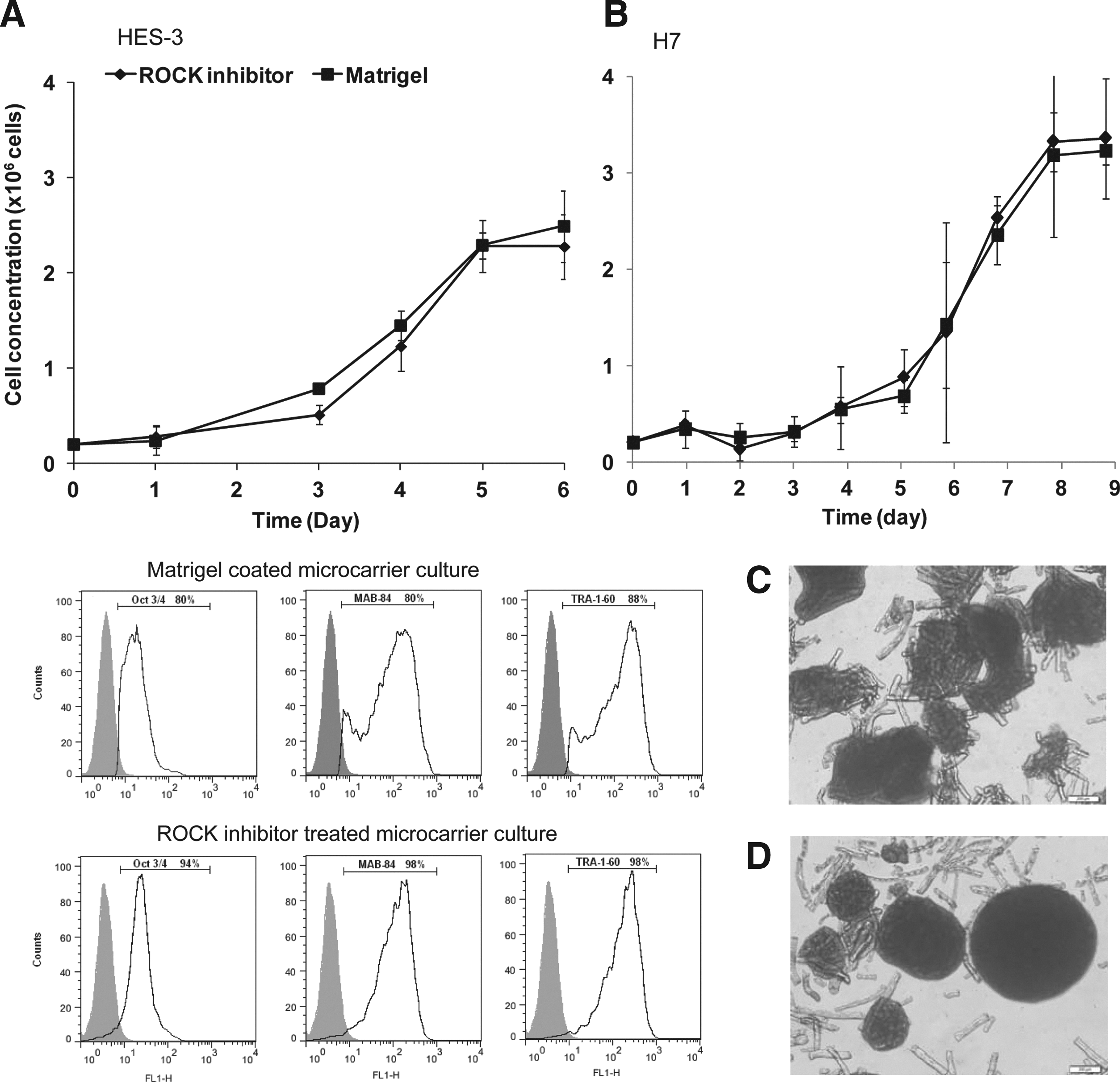
Comparison of cell growth, pluripotent marker expression, and morphology of hPSCs cultured on Matrigel-coated and noncoated ROCK inhibitor-treated DE-53 microcarrier spinner flask cultures.
Embryoid body culture and teratoma generation in SCID mice were done to evaluate the differentiation capability of the ROCK inhibitor-treated microcarrier cultures. Increased transcription levels of representative genes from three germ lineages and a decrease in pluripotent markers (Oct4 and Nanog) were demonstrated in embryoid body (cell–microcarrier aggregates) cultures (Fig. 6A). Immunostaining of differentiated embryoid bodies plated on gelatin-coated tissue culture plates also showed positive expression of representative endoderm (AFP), ectoderm (β-III Tubulin), and mesoderm (SMA) proteins (Fig. 6B). Teratoma generated in SCID mice contained tissues from the three germ lineages, namely, rosettes of neural epithelium, gut-like epithelium, and cartilage (Fig. 6C).
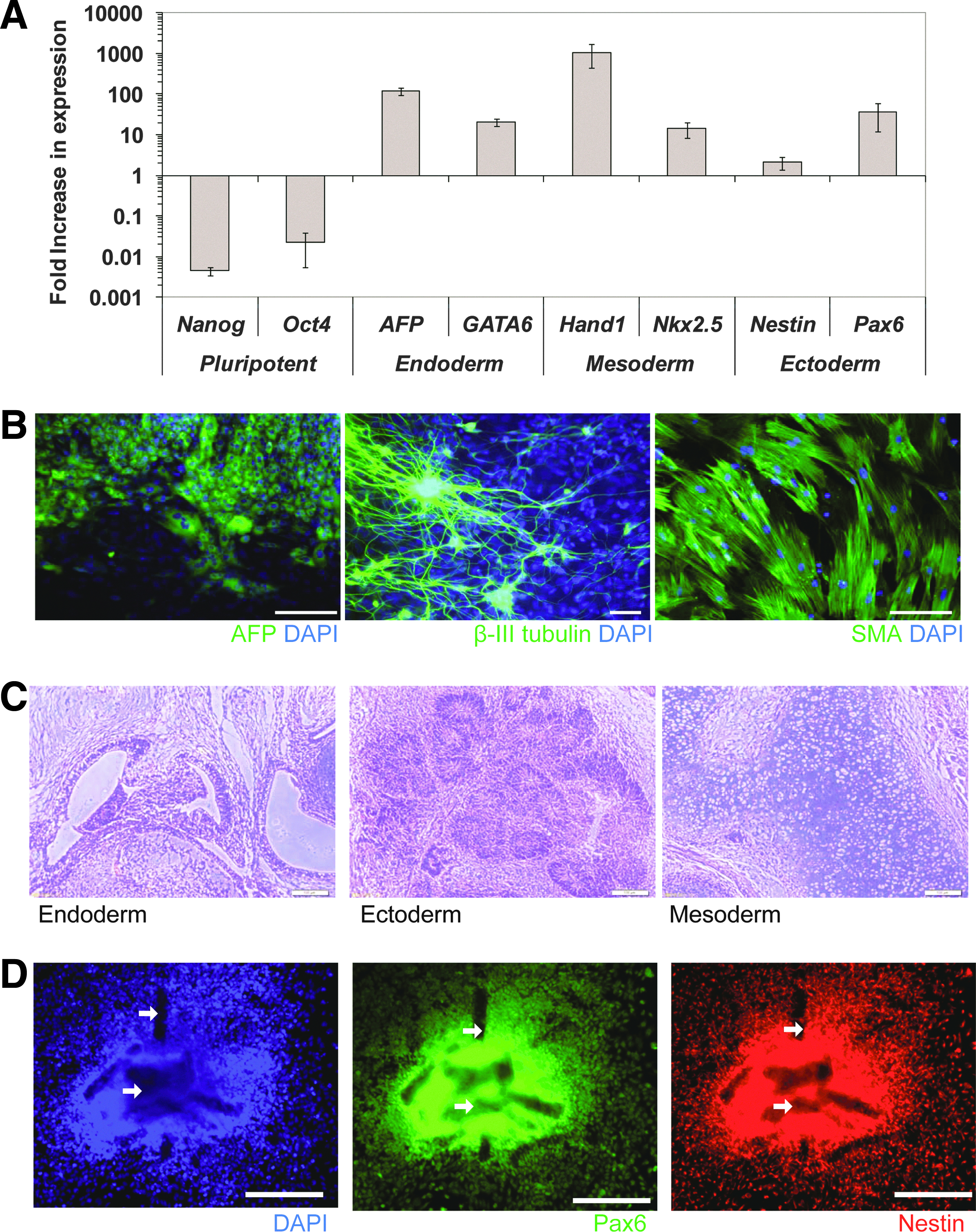
ROCK inhibitor-treated HES-3 microcarrier cultures have differentiation capabilities.
Noncoated, ROCK inhibitor-treated HES-3 microcarrier spinner flask cultures were differentiated to NPCs using the protocol described by Bardy et al. 21 After 16–20 days of differentiation in stirred spinner culture, highest levels of PSA-NCAM-positive NPCs were obtained (90.9±1.1% PSA-NCAM-positive, viable cell density of 9.1±1.2×106 cells/mL, and yield of 412±77 NPCs per seeded HES-3). These yields are comparable to the highest ones obtained from a parallel Matrigel-coated microcarrier culture at day 16 (94.5±2.8% PSA-NCAM-positive cells, viable cell density of 9.3±0.6×106 cells/mL, and yield of 368±43 NPCs per seeded HES-3) (Supplementary Fig. S3). These cells also expressed typical NPC markers (PAX6 and Nestin) when replated on Matrigel-coated plates (Fig. 6D).
Discussion
Microcarrier technology has proven to be a scalable culturing platform for hPSC expansion and subsequent differentiation to definitive endoderm, neural progenitors, and cardiomyocytes.17–19,21,23,37,38 Similar to 2D culture, for hPSC cultivation, Matrigel coating has been shown to be essential for growth on microcarriers.17,19 The complexity of Matrigel composition and its animal origin may pose a challenge in applying the hPSC microcarrier expansion platform for clinical applications. Studies have reported alternatively defined matrices like laminin (mouse and human),6,8,22,39,40 human vitronectin (plasma purified or recombinant),9,10,22 and fibronectin 40 can replace Matrigel coating for supporting hPSC cultivation. However, these ECM proteins may not be suitable for large-scale production as they either contain components of animal origin or are expensive to produce and purify.
In this study, we have shown that hPSCs (hESCs and hiPSCs) can be cultured on noncoated microcarrier cultures for at least 12 passages simply by adding ROCK inhibitor (Y27632) or Blebbistatin. We also demonstrate that cell growth, expression levels of pluripotency markers, and scalability of ROCK inhibitor- and Blebbistatin-treated microcarrier cultures are comparable to those cultured on Matrigel-coated microcarriers. Both in vitro and in vivo spontaneous differentiation procedures were able to generate cells of the three germ layers. Moreover, efficient differentiation to NPCs was demonstrated in stirred cultures. We have found that long-term exposure of hPSCs to ROCK inhibitor has no adverse effect on cell growth, pluripotency, karyotype stability, and directed differentiation to neural progenitors, which is consistent with similar findings in other studies.14,41–44 The use of ROCK inhibitor eliminates the need of ECM coatings on microcarriers for hPSC cultivation. This is an important step toward the development of a defined serum-free hPSC expansion platform for both research and clinical applications.
ROCK inhibitor, Y27632, has been used widely in a variety of applications that involved hPSC dissociation.14,32,35 It has been shown that addition of ROCK inhibitor to dissociated hPSCs can improve the cell survival rate and plating efficiency.14,32,35 Other studies also showed improved recovery of cryopreserved hPSCs or its differentiated progenies when ROCK inhibitor was added. 45 Several groups have tried to address the underlining mechanism of ROCK inhibitor in preventing cell dissociation-induced apoptosis. It has been shown that ROCK inhibitor does not prevent the apoptosis directly. Prevention of apoptosis was attributed to ROCK-dependent myosin II hyperactivation.13,32,35,46 We were not successful in achieving stable hESC aggregate cultures with ROCK inhibitor alone. However, several groups have utilized ROCK inhibitor to culture hPSCs as cell aggregates without microcarriers.38,47–51 These aggregate cultures have been reported to reach comparable cell concentrations of 1.0–1.9×106 cells/mL (2.5–17.7-fold expansion).48,49,51 Even though these cell aggregate cultures have advantages in cost reduction and simpler downstream processing as compared to microcarrier cultures, they have several limitations that still need to be addressed. In cell aggregate culture, where the whole aggregates are comprised of cells only, the lack of proper monitoring can lead to excessive multilayered cells and result in limited nutrient diffusion. Whereas in microcarrier culture, such excessive multilayered cells would not occur when adequate microcarriers are provided in the culture. As shown in Figures 3 and 5, cell–microcarrier aggregates had layer thickness of up to 10 cells with microcarriers embedded in the microcarrier–cell aggregates. Hence, nutrient deficiency due to diffusion limitation is significantly lower in microcarrier culture than aggregate culture. In previous publications, we have demonstrated that hESC microcarrier culture achieve higher cell yields than 2D cultures (1.7–1.9-fold) and over 90% viability indicating no nutrient diffusion limitation.17,21 Moreover, there is no need for manual manipulation of the culture during the 7 days of cell growth and the culture can be passaged efficiently with or without enzymatic treatment at wide range of seeding densities. 27 In aggregate cultures, frequent intervention and stringent controls are needed, cells are seeded at controlled low density (cell aggregate), and manual dissociation is needed to control the aggregate size.49–52 Moreover, during culture passaging, significant cell death occurs due to enzyme disaggregation treatment.49–52
hPSCs have net negative surface charges and attach efficiently to positive charged cellulose microcarriers. 19 This cell attachment is adequate to support hPSC spreading and growth for the short term (two passages) on noncoated microcarriers, but not for long-term growth and maintenance of pluripotency.17,19,21 In this study, we show that hPSCs can grow on these noncoated microcarriers in the presence of ROCK inhibitor and Blebbistatin. These cultures exhibit significant lower expression levels of pMYPT1 and pMLC2 proteins (as compared to Matrigel-coated microcarrier cultures), indicating a reduction in myosin II-mediated cell contractility. The involvement of the ROCK–Myosin II signaling pathway in supporting hPSC growth on noncoated microcarriers was validated by using myosin II inhibitor (Blebbistatin). We postulate that these inhibitors mediated cytoskeletal reorganization, in which, myosin II is the key regulator that enables hPSCs to maintain growth and pluripotency on noncoated microcarriers.
ROCK inhibitor-mediated cytoskeletal reorganization affects spatial organization (i.e., the microenvironment) of hPSCs on microcarriers via altering cell–cell and cell–ECM interactions similar to those observed in 2D hPSC cultures. This microenvironment has been widely known to determine the fate of hPSCs.53,54 The increase in cell–cell interaction via E-cadherin modulation has been reported to improve hPSC survival. 35 Addition of ROCK inhibitor to hPSC colonies was shown to increase Oct4 expression, which was attributed presumably to the reduction in actomyosin tension within hPSC colonies.54,55 Pakzad et al. 44 showed that the addition of ROCK inhibitor to hPSC culture exhibited upregulation of integrins (αV, α6, and β1) associated with pluripotency of hPSCs.44,56 The changes in integrin organization led to enhanced cell adhesion to the ECM and disposition of ECM proteins that support self-renewal of hPSCs.44,56 In our study, a high level of cell attachment is achieved by the positively charged surface of the DE-53 microcarriers, in which, 85–93% cell attachment was observed in 15 min (data not shown). This suggested that in the case of positively charged microcarrier cultures (DE-53), ROCK inhibitor treatment-mediated integrin expression might not play a significant role in hPSC attachment efficiency to the DE-53 microcarriers. There are many studies and reviews addressing the issue of the effect of ROCK inhibitor treatment on cytoskeletal reorganization, hPSC growth, pluripotency, and differentiation propensity.32,42,57–63 However, up until now, the direct correlation between ROCK inhibitor-mediated cytoskeletal reorganization and the mode of hPSC growth has not been elucidated convincingly. More extensive basic research is needed to clarify these issues.
Agitation stress has been shown to influence differentiation commitment in hPSCs. Leung et al. showed that hESCs (HES-3) propagated on Matrigel-coated DE-53 microcarriers in agitated spinner flasks exhibited lower pluripotency marker expressions when compared to static microcarrier cultures. 36 A later study by Chen et al. showed the impact of ECM microcarrier coating on pluripotency of hPSCs in agitated spinner flasks. hESCs propagated on laminin-coated microcarrier spinner flasks were more prone to differentiation than Matrige- coated ones. 19 Similar observations of shear-induced differentiation were also found with mesenchymal stem cells (MSCs). Datta et al. demonstrated synergistic effects of fluidic shear and ECM coating on scaffolds on osteogenic differentiation of rat MSCs. 64 Arnsdorf et al. also found that exposure of murine MSCs to oscillatory flow induced increased cytoskeletal tension and expressions of differentiation markers. 31 Both the cytoskeletal changes and differentiation could be curtailed by addition of ROCK inhibitor or Blebbistatin. 65 Teramura et al. showed similar effects using hiPSC, ROCK inhibitor treatment prevented downregulation of pluripotent markers caused by cyclic tensile forces. 66 In this study, we have shown that ROCK inhibitor treatment reduces agitation shear-induced spontaneous differentiation observed previously in ECM-coated microcarrier spinner flask cultures. 19 As shown in Figure 5, both hESC lines (HES-3 and H7) treated with ROCK inhibitor displayed higher expression levels of pluripotency markers when compared to untreated hESCs on Matrigel-coated microcarriers. Based on this observation, it seems that cytoskeletal reorganization resulting from ROCK inhibitor treatment allows hPSC microcarrier cultures to resist fluidic shear-induced spontaneous differentiation.
In conclusion, we have demonstrated culturing of hPSCs on noncoated microcarriers in a defined serum-free medium. The elimination of ECM coatings from the microcarrier surface will facilitate the translation of the hPSC microcarrier expansion platform into clinical applications. As discussed above, we have shown that probably due to ROCK inhibitor-mediated cytoskeleton changes, hPSCs can be propagated on noncoated microcarriers. However, further study is needed to address the complexity of mechanotransduction pathways and their impact on hPSC microcarrier culture.
Footnotes
Acknowledgments
The authors would like to acknowledge Dr. Andre Choo (Bioprocessing Technology Institute, Singapore) for the teratoma assay and Dr. Kah Yong Tan (Bioprocessing Technology Institute, Singapore) for his critical review of the manuscript. This work was generously supported by the Agency for Science, Technology and Research (A*STAR), Singapore.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.