
Abstract
Recent cartilage engineering efforts have focused on development of zonally organized tissue. However, there remains a need for protocols that differentiate progenitor populations into chondrocytes of zonal phenotype. Here, we evaluate the potential of coculture of bovine mesenchymal stem cells (MSCs) and zonal explants of bovine cartilage tissue to drive MSC differentiation to chondrocytes with the superficial zone phenotype. Two coculture systems were set up: one between alginate encapsulated MSCs and superficial zone cartilage explants, and one between MSCs and middle/deep zone cartilage explants. Chondrogenic and superficial zone markers were monitored over a 21-day differentiation period via gene and protein expression. A control conditioned media study was used to determine the impact of communication via soluble factors between cell populations during differentiation. At day 21, results show superficial zone explant coculture without transforming-growth factor β3 supplementation induces upregulation of chondrogenic gene expression markers SOX9 and type II collagen 3.4-fold and 11.4-fold, respectively, over standard chondrogenic control media. Further, coculture of MSCs and superficial zone explants can be used to upregulate mRNA expression of the superficial zone marker proteoglycan-4 in MSCs (1.75-fold over chondrogenic control at day 21), indicating the superficial zone chondrocyte phenotype. Gene expression data show middle/deep zone explant and MSC coculture did not induce the chondrogenesis observed in superficial zone explant coculture. Likewise, poor chondrogenesis was observed in all conditioned media groups. Results highlight the importance of superficial zone cartilage and cells in guiding stem cell fate and regulating differentiation of MSCs to chondrocytes of the superficial zone type.
Introduction
I
The superficial zone is typically the first to become degraded during diseases such as osteoarthritis. 9 Therefore, therapeutic interventions must either aim to prevent the loss of this layer at early disease stages, or to regenerate this layer in later disease stages. Growing attention has been focused on such regeneration of stratified repair tissue,10–12 however, cell source remains a major challenge for zonal cartilage engineering efforts. Bone marrow-derived mesenchymal stem cells (MSCs) are potential treatment populations for articular cartilage defects. Typically, MSCs are cultured in a micromass or hydrogel and delivered chondrogenic media supplemented with transforming-growth factor βs (TGF-βs) and dexamethasone to induce chondrogenesis. Methods for inducing zone-specific chondryctes are yet to be established, but growth factor delivery 13 and cues from the scaffold environment 11 show potential for guiding progenitor populations to zone-specific cells.
Here, we investigate a unique method of inducing zone-specific chondrocytes from MSCs: soluble signals derived from zonal cartilage tissue explants. While coculture of these two populations has been reported,14–19 results have been mixed, and there are no reports of zonal coculture models. A clear trend has not been established on the influence or the mechanisms of influence between these two cell populations. Some studies report that chondrocyte-secreted factors alone can influence differentiation of MSCs. For example, it was reported that cartilage explants secreted TGF-β for up to 14 days, and soluble factors derived from cartilage explants were able to upregulate SOX9 gene expression and type II collagen synthesis in MSCs. 14 Similar studies confirmed that soluble factors derived from chondrocytes were able to upregulate chondrogenic gene markers and matrix production by MSCs,15,16 and reduce hypertrophic markers in MCSs.14,17 However, the chondrogenesis induction reported via chondrocyte-conditioned media showed only moderate type II collagen gene upgregulation over control media and predominately type I collagen matrix accumulation. 16 Additional work reports direct contact culture of MSCs and chondrocytes enhanced MSC differentiation.18,19
Conflicting results are reported on the effects of MSCs and chondrocytes cultured directly together in micromass or in hydrogels. Some studies report a direct benefit of coculture on MSC chondrogeneis.18–21 However, from these studies it is not always clear which population is upregulated for gene and protein expression of chondrogenic markers.22–24 While the two cell populations definitely influence each other, from current reports it is unclear whether MSCs are primarily responsible for upregulating the chondrocyte phenotype in the chondrocyte population, or whether chondrocytes are primarily responsible for inducing chondrogenesis in the MSC population—or both.
In the present work, we aim to establish the use of zonal cartilage-derived soluble factors to drive differentiation of MSCs to chondrocytes of the superficial zone phenotype. To this end, we investigate a coculture model of MSCs and cartilage explants of both the superficial and middle/deep zones. A control conditioned media study was also performed to assess the impact of communication between cell populations. Due to the distinct phenotype and function of superficial zone chondrocytes, we hypothesize signals derived from this group will have a unique effect on the differentiation of MSCs. The goals of the presented work are to determine whether soluble signals from superficial zone cartilage explants can drive differentiation of MSCs to the superficial chondrocyte phenotype, what effects explants from the middle/deep zone have on MSC differentiation, and whether communication between cell populations impacts differentiation of MSCs.
Materials and Methods
Superficial and middle/deep zone explant section isolation
Zonal chondrocyte explants were isolated according to previously published laboratory protocols.25,26 Briefly, cartilage plugs (4 mm diameter, 3–6 mm height) were harvested from the femoral condyles of 20-week-old calves using a Sklar Tru-Punch disposable biopsy punch (Sklar Instruments). The top 10% (0.3–0.6 mm) was taken as the superficial zone, and remaining tissue up was defined as the middle and deep tissue zones.
Bone marrow stromal cell isolation
Primary bovine bone marrow tissue was harvested from the tibia of 3-week-old calves. The tissue was suspended in growth media (Minimal Essential Medium α [MEMα], (Gibco/Invitrogen) supplemented with 0.1% penicillin/streptomycin antibiotics and 0.2 mM of ascorbic acid), filtered through a 70 μm mesh, and centrifuged to isolate the cell population. This population was then enriched for MSCs via plastic adhesion by plating in monolayer and culture in growth media with 10% fetal bovine serum. Then, two passages cells were trypsinized (Gibco/Invitrogen) and counted using Trypan blue staining and a hemacytometer.
Cell encapsulation and coculture
About 2.0% w/v alginate was prepared from alginic acid sodium salt from brown algae (Sigma-Aldrich) and was mixed with isolated MSCs and injected through an 18-gauge syringe into continuously stirred 0.1 M calcium chloride (Sigma-Aldrich). The resulting cellular density was ∼100,000 per bead, and each spherical bead had a diameter of ∼5 mm (∼2×106 cells/mL). Alginate concentration and cell density were chosen based on previous results from the lab indicating high cell viability. 27 Control beads were cultured in serum-free chondrogenic media made of high-glucose MEMα (Gibco/Invitrogen)+110 μg/mL sodium pyruvate, 40 μg/mL proline, 50 μg/mL ascorbate 2-phosphate, 0.1 μM dexamethasone, 1% ITS+premix (BD Biosciences), and 10 ng/mL TGF-β3 treatment (R&D Systems). Coculture groups were set up with either superficial or middle/deep explants chips in the bottom of a six-well plate, a transwell membrane placed in the well, and alginate beads containing MSCs on top of the membrane. Each coculture well contained five alginate beads, minced tissue from three explants (see section “Superficial and middle/deep zone explant section isolation”), and 10 mL of media. At each time point the five alginate beads in a well were pooled and RNA was isolated. See Figure 1A for illustration of coculture experimental setup. Each group cocultured with superficial or middle/deep explants chips was delivered chondrogenic media with or without 10 ng/mL TGF-β3, and media was changed every 48 h. At days 1, 7, and 21 MSCs were isolated from the alginate beads by delivery of 0.1 M ethylenediaminetetraaceticacid (EDTA) for 20 min at 37°C. The solution was then centrifuged to form a cell pellet, which was resuspended in phosphate-buffered saline and used for RNA extraction. At days 7 and 21 beads were fixed for staining.

Conditioned media incubation and delivery
To control communication between cell populations a conditioned media study was set up with the corresponding experimental groups. This experiment was performed to compare differences in the ability versus inability of soluble factors from each cell population, chondrocyte and MSC, to influence the other cell type. In the coculture experiment soluble factors from the MSCs can access the cartilage and vice versa, while in the conditioned media experiment only soluble factors from the cartilage can access the MSCs. In addition, the cartilage cells have no ability to sense the presence of a proximal progenitor population. Zonal explants chips and alginate encapsulated MSCs were cultured in separate wells. Growth factor-free chondrogenic media was delivered to zonal explant chips and allowed to incubate for 48 h, after which the media was collected, relevant groups were supplemented with TGF-β3, and it was delivered to alginate encapsulated MSCs. Throughout the study fresh growth factor free chondrogenic media was delivered to the explants every 48 h, and the media that was explant conditioned for 48 h was delivered to the alginate encapsulated MSCs. See Figure 1B for illustration of conditioned media experimental set up. At days 1, 7, and 21, cells were isolated for RNA extraction, and at days 7 and 21 beads were fixed for staining.
Histological preparation
At desired time points, alginate beads were recovered and fixed for 3 h at room temperature in 4% paraformaldehyde (Sigma-Aldrich) with 0.1 M sodium cacodylate and 10 mM calcium chloride. Samples were then placed in histological cassettes and dehydrated through a series of ethanol washes followed by two Citrisolv (Fisher Scientific) washes. The samples were embedded in paraffin (Paraplat X-tra; Fisher Scientific) and cut into 4 μm sections and mounted on a glass slide (Superfrost; Fisher Scientific).
Histochemical staining
Samples were dried at 64°C for 2 h, deparaffinized using Citrisolv, and rehydrated. The samples were then stained using Alcian blue and Sirius red staining solutions (Poly Scientific). All samples were viewed under an Axiovert 40CFL light optical microscope (Zeiss) and images were captured using SPOTSOFTWARE (Diagnostic Instruments, Inc.) imaging software.
Immunohistochemistry
Samples were dried at 64°C for 1 h, deparaffinized using Citrisolv and rehydrated. Samples were antigen retrieved using a Tris base and EDTA buffer (pH 8) containing TWEEN 20 steamed for 15 min in a vegetable steamer. Samples were incubated with PEROXIDAZED1 (Biocare), an endogenous peroxidase blocker, and Background SNIPER1 (Biocare), a blocking reagent. Samples were then stained with primary antibody to detect PRG4. The primary antibody used was anti-lubricin/PRG4 (rabbit polyclonal antibody that reacts with Human, Bovine, and African Green Monkey lubricin, ab28484; Abcam) diluted to a working concentration of 4 μg/mL. The HISTOSTAIN®-SP kit (Zymed) was used to visualize PRG4 presence by using the horseradish peroxidase-streptavidin-biotin system. The complex formation was then detected by a 3,3′-diaminobenzidine tetrahydrochloride chromogen. Samples were counterstained with hematoxylin, dehydrated, cleared in Citrisolv, and covered.
RNA isolation
Following isolation of MSCs from alginate beads, RNA was isolated using the RNeasy Mini Kit (Qiagen). Total RNA was eluted into 30 μL of RNase-free water and detected using a NanoDrop ND-1000 Spectophotometer (NanoDrop Technologies). RNA concentrations at 1, 7, and 21 days were diluted to ∼10 ng/μL.
Quantitative reverse transcriptase–polymerase chain reaction
Isolated RNA was reverse transcribed using a cDNA Archive Kit (Applied Biosystems), which can convert up to 10 μg of RNA to cDNA. cDNA was mixed with Universal Master Mix (Applied Biosystems) and oligonucleotide primers and TaqMan probes (Applied Biosystems) for the genes of interest and a control gene. Table 1 shows the sequences for all forward primers, reverse primers, and probes used. Glyceraldehyde 3 phosphate dehydrogenase (GAPDH) was the endogenous control gene. The reaction volume was 20 μL, and the final concentration of cDNA per reaction well was ∼0.5 ng/μL (10 ng per well). The reaction was conducted on a 7900HT Fast Real-Time PCR System Prism 7000 sequence detector (Applied Biosystems). The thermal profile followed was 2 min at 50°C, 10 min at 95°C, 40 cycles of 15 s at 95°C, and 1 min at 60°C. Gene expression profiles were analyzed using the comparative Ct method. The day 1 control alginate samples were used as calibrators in all analysis to allow for changes over time to be appreciated throughout the differentiation process. Fold changes in gene expression were calculated and are reported as the mean relative quantification (RQ) values with associated standard deviations (n=3), in accordance with methods previously described by our laboratory.28,29
GAPDH, glyceraldehyde 3 phosphate dehydrogenase; PRG4, proteoglycan 4; qRT-PCR, quantitative reverse transcriptase–polymerase chain reaction.
Statistical analysis
For gene expression data each time point was analyzed in technical triplicate (n=3), gene expression data was confirmed by consistent trends in biological replicates (data not shown). All data was analyzed using one-way analysis of variance and Tukey's multiple-comparison test to determine statistical differences. A confidence interval of 95% (α=0.05) was used for all analysis and means and standard deviations are shown on each figure.
Results
Results show coculture of cartilage explants and alginate-encapsulated MSCs can induce chondrogenic differentiation in MSCs comparable to that of standard chondrogenic media delivery. This effect is primarily observed in the MSCs cultured in proximity to superficial zone cartilage explants. Specifically, by day 21, all measured mRNA markers of the chondrogenic lineage, including the lubricating protein PRG4, are upregulated in the superficial zone coculture group compared with standard chondrogenic control media.
Figure 2 shows mRNA expression for chondrogenic markers during coculture throughout the culture period by all groups, as seen in Figure 2A. At day 1, expression is similar among all groups, and at day 7 all groups are upregulated over day 1. At day 7, only the M− group shows increased SOX9 gene expression as compared to the control, however, by day 21 a shift in pattern in observed. The control group remains elevated, however, all experimental groups other than superficial zone−TGF-β3 (S−) are significantly downregulated compared with the control. The S− group has the highest SOX9 mRNA expression, upregulated at 3.4-fold over the control.
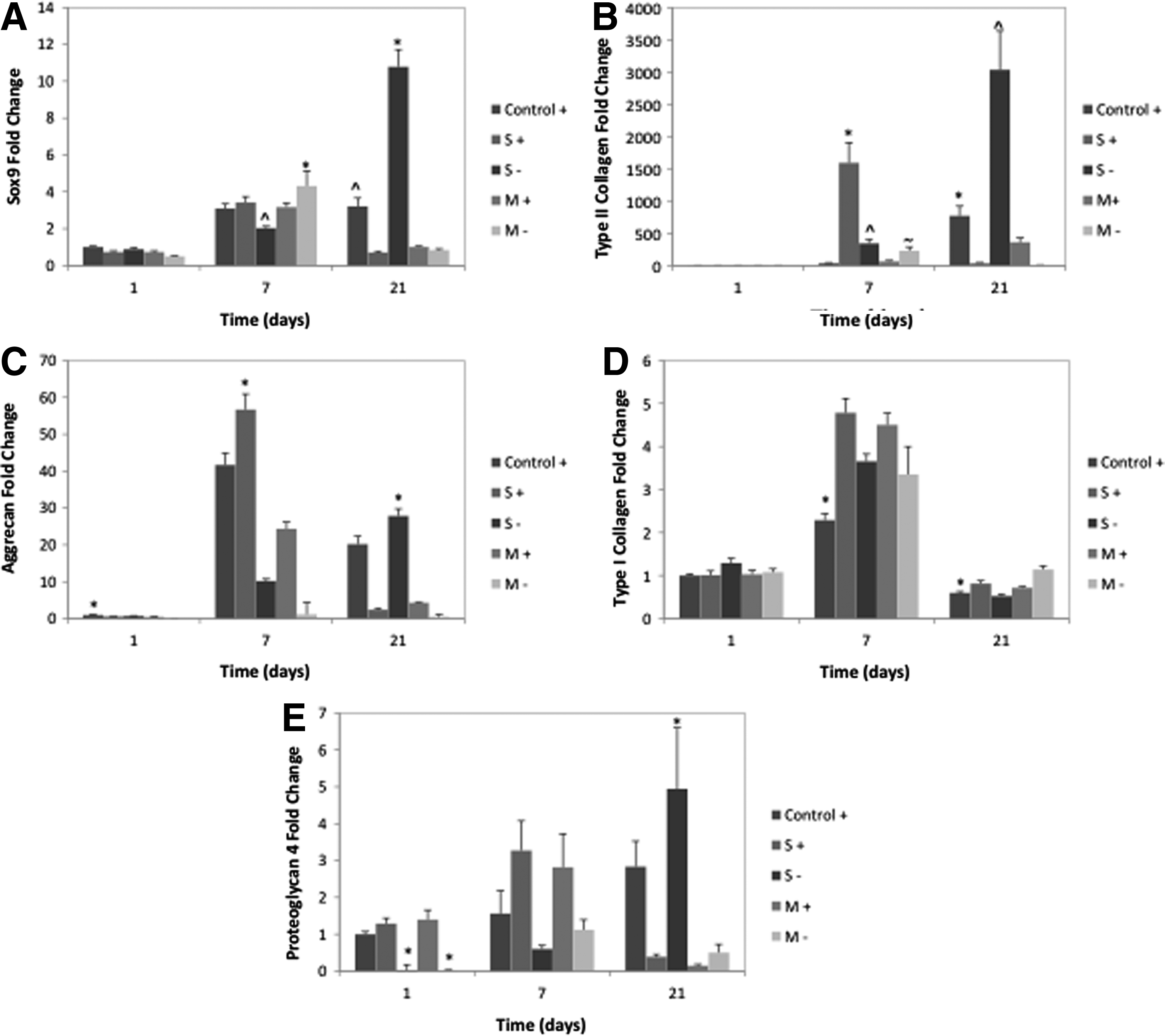
Chondrogenic differentiation markers by mRNA expression of MSCs cocultured with zonal cartilage explant chips. An “S” indicates coculture with superficial zone sections, an “M” with middle/deep zone sections, and a “+” indicates TGF-β3 delivery. Control groups are standard chondrogenic differentiation media with TGF-β3.
Corresponding to this SOX9 upregulation in the S− group at day 21 is significant upregulation in chondrocyte phenotype markers. Type II collagen and aggrecan are major matrix components of articular cartilage and thus mark the chondrogenic lineage. PRG4, unique to the superficial zone, marks the phenotype of these cells. In the S− group on day 21 type II collagen is upregulated 11.4-fold over the control (Fig. 2B), aggrecan 5.6-fold over the control (Fig. 2C), and PRG4 1.75-fold over the control (Fig. 2E), thus indicating that interactions between MSCs and superficial zone explants have a favorable effect on chondrogenic differentiation.
Type I collagen and PRG4 are markers for the chondrocyte and superficial zone chondrocyte phenotype respectively. At earlier time points (days 1 and 7) groups with TGF-β3 delivery show upregulated PRG4 expression indicating that growth factor played a role in its expression. However, by day 21 the only group with elevated expression over the control is the S− group, indicating that exogenous TGF-β3 delivery is no longer as effective, or another influence of the coculture system has come into play. Type I collagen, a marker for fibrocartilage not articular cartilage, is significantly elevated in all experimental groups on days 7 and 21 other than the S− group on day 21 for which it is significantly downregulated (1.14-fold compared to the control group), again indicating a favorable effect of the S− coculture group on chondrogenesis.
Figure 3 shows immunostaining for type II collagen and PRG4. Control and experimental groups show antibody specific staining for type II collagen, confirming production of matrix products. In Figure 3A, PRG4 specific staining, the protein is clearly detected, however, no significant differences are observed between time points or groups, as reported in other hydrogel systems. 30 Figure 4 shows histology staining for proteoglycans by Alcian Blue (Fig. 4A) and collagens by Sirius Red (Fig. 4B), both confirm matrix product production with no major differences observed between groups.


Figure 5 shows chondrogenic mRNA markers over the course of the conditioned media study. All experimental groups are downregulated compared to a control delivered chondrogenic media supplemented with TGF-β3. There is a decrease in SOX9 mRNA expression in all experimental groups at all time points compared to the control. Following the same trend, the expression of type II collagen is significantly reduced in all experiment groups compared with the control at days 7 and 21 indicating a lack of the chondrocyte phenotype. At both later time points there are no significant differences between type II collagen expression in experimental groups and the control is on average 236-fold and 47-fold greater than the experimental groups on days 7 and 21 respectively. Likewise, matrix and protein production is limited in all experimental groups as seen in Figures 6 and 7. Protein and gene expression data confirm poor induction of chondrogenic markers in all conditioned media experimental groups.

Chondrogenic differentiation markers by mRNA expression of MSCs cultured in zonal cartilage explant conditioned media. An “S” indicates delivery of superficial zone conditioned media, an “M” with middle/deep zone conditioned media, and a “+” indicates TGF-β3 delivery. Control groups are standard chondrogenic differentiation media with TGF-β3.

Discussion
Coculture of superficial zone explants and alginate-encapsulated MSCs drives differentiation of MSCs to chondrocytes of the superficial zone phenotype. At day 21 of culture, the superficial zone coculture group without TGF-β3 supplementation (S−) has significantly greater mRNA expression of chonrogenic markers than all other groups: SOX9 expression is 3.4-fold greater than the control chondrogenic media group, type II collagen is 11.4-fold great than the control, aggrecan is 5.6-fold greater than the control, and PRG4 is 1.75-fold greater than the control. Soluble signals derived from superficial zone cartilage explants drive chondrogenesis and differentiation of PRG4 mRNA-expressing MSCs. At day 1, few differences are observed between the groups. At the intermediate time point of day 7, the group with highest gene expression for chondrogenic markers is the superficial zone coculuture groups with TGF-β3 supplementation (S+). Surprisingly, at the study endpoint (day 21) presence of TGF-β3 in the culture media in the superficial zone groups inhibits the chondrogenesis, and the highest gene expression for chondrogenic markers is seen in the superficial zone coculture group without TGF-β3 (S−) group. While the mechanism for this observed difference is unclear the addition of large amounts of soluble protein may disrupt the concentrations of available proteins and change the profile of proteins available to the cell for binding and initiation of cell signaling. Although middle/deep zone coculture groups express similar levels of markers on days 1 and 7, by day 21 this expression is considerably decreased in all groups. It is important to note that our control in all experiments is a positive control for chondrogeneic induction that contains TGF-β3. Thus, mRNA upregulation, which is comparable or above the control, indicates the coculture strategy is similar in differentiation efficiency to that of standard media delivery protocols.
Cells of the superficial zone are metabolically distinct from those of the middle and deep zones,31–33 so variation in their influence over a progenitor population is not surprising. The most established and understood difference between the superficial zone and deeper tissue zones is the production and presence of the lubricating protein PRG4, which is critical for boundary mode lubrication at the articulating surface.2,3,34,35 Superficial zones cells are also physically different than middle and deep zone cells with a flattened morphology and smaller size. 8 Recent evidence also supports another major difference between the top layers of articular cartilage tissue and the lower layers: the presence of a progenitor cell population.36–40 While the presence of a progenitor population within articular cartilage has now been independently reported by several groups,36,37,39 the relative percentage of the progenitor population and the precise distribution has not been determined. Reports of the relative percentage of cells with stem cell markers range from 0.14% to 45% of the tissue's total cell population, depending on detection method and definition.37,39,40 At present it is unclear what the relative percentage of stem/progenitor cells is and exactly where that population is zonally distributed. However, results support that the superficial zone contains the highest relative percentage of a progenitor population compared with other zones. 38 The absence of a progenitor population in the middle/deep zone may result in the tissue from this zone lacking the soluble signaling factors necessary to guide progenitor cell fate. Thus, in a region of the tissue with no progenitor population signals are not present to drive differentiation. As seen in Figure 2, middle/deep coculture groups do not provide soluble signal to drive MSC chondrogensis.
From the results of our coculture study, it is clear that the superficial zone is able to provide distinct signals to differentiating MSCs, which results in expression of chondrogenic markers, and even the superficial zone chondrocyte marker, PRG4. The gene expression data for chondrogenic markers and histological and immunochemical staining demonstrate that this differentiation is comparable to that induced by the positive chondrogenic media control. Reports from the literature support that mesechymal progenitor cells express PRG4 during TGF-β-induced chondrogensis, however, the mechanisms and pathways involved in this expression are not clear.12,26,30 As evidence now supports a population of progenitor cells within the superficial zone, stem cell differentiation may be a normal biological process within this zone, and thus signals derived from it provide favorable signals for chondrogenesis. What is not clear from our data is the source, or identity, of the factors that induce chondrogenesis in the superficial zone coculture groups. The explanted tissue would contain both a superficial zone chondrocyte population and the reported progenitor population; therefore, either or both populations may have influenced MSC differentiation. In terms of responsible soluble factors, previous studies in our laboratory show superficial zone cells in alginate culture express significantly higher levels of insulin-like growth factor 1 (IGF1) mRNA compared with middle and deep zone cells, and reduced levels of IGF-binding protein mRNA—indicating higher levels of protein available for cell binding. 25 In addition, TGF-β1,2,3 are reported at elevated levels in superficial zone tissue. 41 In native cartilage, TGF-βs are synthesized in a latent complex, which is unable to bind to cell membrane receptors. 42 It was recently demonstrated that the primary mechanism of TGF-β activation is via mechanical shearing in the synovial fluid, rather than chemically induced factors. 43 Further, while large amounts of latent TGF-βs are synthesized and secreted from both the synovial fluid and all three cartilage zones (superficial, middle, and deep), upon shear-induced activation TGF-βs bind and accumulate preferentially in the superficial zone. This zone-specific accumulation leaves the superficial zone with up to 35 times the concentration of activated TGF-βs than the middle and deep zones. 44 It has also been demonstrated that the presence of TGF-β is necessary for PRG4 production by superficial zone chondrocytes,45,46 indicating that the shear-induced activation may be required for active TGF-βs to accumulate in the superficial zone, and for the cells of the superficial zone to utilize these signals in maintenance of their zonal phenotype. These studies indicate that the significantly higher levels of activated TGF-βs present in superficial zone cartilage may influence the differentiation and PRG4 expression of proximal MSC populations—both in vivo and in our coculture model.
Our results also show that the chondrogenesis induced by the coculture groups is dependent on communication between the explanted tissue cell populations and the MSCs, as evidenced by the control conditioned media study. Control groups within the conditioned media study received standard chondrogenic media, and display normal differentiation profiles in terms of gene and protein expression. However, chondrogenesis is not induced by media conditioned by explants. This provides the hypothesis that the differentiation observed in the superficial zone coculture groups is a result of critical communication between cell populations. Without sensing the presence of an undifferentiated progenitor population explanted cells are not triggered to secrete signaling proteins to guide stem cell fate. However, in the presence of a progenitor population, superficial zone chondrocytes or MSCs in the explanted tissue secreted signals to guide MSCs differentiation. It also possible that signals secreted by the MSC population influenced phenotype retention of the explanted chondrocytes, which further increases the ability of the chondrocytes to secrete signaling molecules and direct stem cell fate. This phenomenon has previously been reported,22,23 however, the mechanism remains unknown, and is likely to be a factor in the coculture driven differentiation observed in our studies.
It is important to note that both chondrocyte and MSC populations are from healthy juvenile bovine populations, and that differences in the activity of the isolated cell populations and that of adult or diseased human cell populations likely exist. Adult/diseased populations are likely to be less metabolically active and possibly involved in production of signaling molecules that cause further tissue damage. Upon application of tissue engineering strategies the age and disease state of the utilized cell populations must be taken into careful consideration.
Conclusion
In the presented study we demonstrate that cartilage explants from the superficial zone of articular cartilage are able to induce MSC chondrogenesis through soluble signaling factors, and that this chondrogenesis is comparable to that induced by positive chondrogenic control media. The highest expression of chondrogenic markers is observed in the superficial zone coculture group without exogenous TGF-β3 delivery on day 21 of culture. Superficial zone explants are also able to provide signals that upregulate the superficial zone phenotype marker PRG4 in the encapsulated MSC population. Further, we show that the coculture-induced chondrogenesis is dependent on communication between the cell populations. Conditioned media from the same zonal explants was unable to induce chondrogenesis, even when supplemented with exogenous TGF-β3. We provide further evidence of important differences between the zones of articular cartilage and show that signals derived from the superficial zone have a distinct role in guiding progenitor cell fate.
Footnotes
Acknowledgments
This work was supported by the National Science Foundation (CBET 1264517) and the National Institutes of Health (R01 AR061460).
Disclosure Statement
No competing financial interests exist.