
Abstract
Soft alginate hydrogels support robust neurite outgrowth, but their rapid disintegration in solutions of high ionic strength restricts them from long-term in vivo applications. Aiming to enhance the mechanical stability of soft alginate hydrogels, we investigated how changes in pH and ionic strength during gelation influence the swelling, stiffness, and disintegration of a three-dimensional (3D) alginate matrix and its ability to support neurite outgrowth. Hydrogels were generated from dry alginate layers through ionic crosslinks with Ca2+ (≤10 mM) in solutions of low or high ionic strength and at pH 5.5 or 7.4. High- and low-viscosity alginates with different molecular compositions demonstrated pH and ionic strength-independent increases in hydrogel volume with decreases in Ca2+ concentrations from 10 to 2 mM. Only soft hydrogels that were synthesized in the presence of 150 mM of NaCl (Ca-alginateNaCl) displayed long-term volume stability in buffered physiological saline, whereas analogous hydrogels generated in NaCl-free conditions (Ca-alginate) collapsed. The stiffnesses of Ca-alginateNaCl hydrogels elevated from 0.01 to 19 kPa as the Ca2+-concentration was raised from 2 to 10 mM; however, only Ca-alginateNaCl hydrogels with an elastic modulus ≤1.5 kPa that were generated with ≤4 mM of Ca2+ supported robust neurite outgrowth in primary neuronal cultures. In conclusion, soft Ca-alginateNaCl hydrogels combine mechanical stability in solutions of high ionic strength with the ability to support neural growth and could be useful as 3D implants for neural regeneration in vivo.
Introduction
B
Alginate is a biocompatible, natural, linear, binary copolymer composed of D-mannuronic acid (M) and its C5 epimer L-guluronic acid (G) monosaccharide units, which are covalently linked by β-1-4 glycosidic bonds. M and G are distributed within the polymer chain in varying proportions to produce heterogeneous alternating (MG) and homogeneous (MM or GG) sequences in the primary structure.5–9 In aqueous solutions (sols), alginate exists as a negatively charged polyanion that forms a hydrogel by the physical association of polysaccharide chains through ionic crosslinking of G residues by multivalent cations (e.g., Ca2+, Sr2+, Ba2+, Zn2+, Cu2+, Ni2+, or Fe3+). This process has been described by the egg-box model in which divalent ions interact with two adjacent G residues and with two G residues of an opposing chain.10,11 The density of the alginate fiber network within a hydrogel and its gel strength are regulated by the number of connected gelling sites; as a result, these properties are elevated with an increasing level of crosslinking ion saturation in the alginate and reduced with an increasing number of free G blocks.6,12–14
The behavior of charged polyelectrolytes, which includes alginates, is strongly regulated by the pH and ionic strength of the surrounding solution; pH controls the degree of dissociation of the guluronic and mannuronic acid groups, while ionic strength triggers the interaction of the resulting negative charges along the polymer chains.15,16 Most polyelectrolyte-based gels disintegrate upon immersion in solutions of high ionic strength. Ionically crosslinked alginate hydrogels undergo disintegration in the presence of calcium chelators (e.g., phosphates), monovalent ions (e.g., Na+), and non-cross-linking divalent ions (e.g., Mg2+), which are present in the tissue culture medium and tissue fluids.17–19 Disintegration of polyelectrolyte-based hydrogels strongly limit their usefulness for in vivo applications, which require long-term 3D stability of the implant. Traditionally, alginate gels were fabricated in NaCl-free solutions; recent studies on gelation speed, swelling, and degradation and on the critical steps in Ca2+ binding to alginates were also performed on hydrogels fabricated in NaCl-free solutions.20–23 Conventional methods to stabilize alginate hydrogels include covalent crosslinking and application of high concentrations of crosslinking cations to ensure tight connections between the polymer chains.24–26
Physical cues such as substrate stiffness are important factors for cell differentiation and behavior.27,28 The successful use of hydrogels greatly relies on the refined control of the mechanical properties, including stiffness. Neurons can sense the stiffness of a substrate and will behave accordingly. For example, soft but not stiff alginate hydrogels support robust neurite outgrowth in vitro. 29 Dorsal root ganglion neurons display maximal growth on soft substrates with stiffnesses <1 kPa.30,31 Neurons plated on softer substrates showed decreased branching relative to neurons grown on stiffer substrates.32,33 Mesenchymal stem cells and primary neural stem cells differentiate into neurons on soft hydrogels with stiffnesses <1 kPa.34–37 Stiff alginate hydrogels were demonstrated to be nonadhesive for neurons; to provide an adhesive surface, these hydrogels required functionalizations with signaling molecules such as laminin and fibronectin38–40 and integration of polyglycolic acid, heparin, or basic fibroblast growth factor (bFGF).41–44 Anisotropic, stiff alginate scaffolds, which were covered with collagen or polylysine or incorporated gelatin, were shown to support neurite elongation along capillary channels, but prohibited neural ingrowth into the hydrogel core.45,46
Soft alginate hydrogels support neural growth, but nothing is yet known about how to enhance their mechanical stability in solutions of high ionic strength without elevation of their stiffness and affecting their interaction with neural cells. This study emphasizes that gelation of alginate layers with substoichiometric concentrations of calcium ions in the presence of NaCl generates soft Ca-alginateNaCl hydrogels, which combine mechanical stability with high adhesion to neural cells. This translational research opens a new perspective on the design of stable 3D polyelectrolyte-based hydrogels for long-term in vivo applications.
Materials and Methods
Materials and reagents
Ultrapure sodium alginates with trade name PRONOVA were purchased from Novamatrix, now part of FMC BioPolymer, as guluronic acid rich alginate (LVG) and mannuronic acid rich alginate (LVM), and endogen-free S-alginate were purchased from Sigma (71238; Taufkirchen, Germany). The molecular compositions of LVG and LVM were provided by the supplier, but the composition of S-alginate was quantified for this study by a proton 1 H-NMR spectroscopy method. 47 LVM, LVG, and S-alginates contained 43%, 68%, and 68% guluronic acid residues, respectively. Reagents, including the neurobasal medium, DMEM/F12 (1:1), B-27 supplement, Dulbecco's Phosphate-Buffered Saline without Ca2+ and Mg2+, and trypsin/EDTA solution were all purchased from Invitrogen (Darmstadt, Germany). CaCl2 dihydrate, HEPES, TRIS, NaCl, and DNase I originated from Sigma.
Preparation of alginate hydrogels at ambient conditions
Alginates were dissolved in deionized water under permanent stirring at room temperature (RT) for 12 h. The resulting aqueous 0.5%, 1.0%, or 1.5% sols were sterilized by pressing through a 0.45-μm sterile filter. Dried alginate layers were formed on glass coverslips (24×24 mm) when each sol was evenly distributed onto a glass surface and dried at RT for 24 h. Dry alginate layers were stored under humidity-free conditions and used within 10 days after preparation. To form a hydrogel, a glass coverslip with a dry alginate layer was placed into a Petri dish (Ø 5 cm) and overlayed with a 15 mL crosslinking solution; the gelation reaction was carried out at RT for 24 h, and a single interconnected gel slice was formed on the glass surface. The excess crosslinking solution was removed before gravimetric and rheological analyses. Representative images of hydrogels were taken with a digital camera. For the cell culture experiments, hydrogels were prepared under sterile conditions in a laminar flow hood as described previously. 29 For rheological characterization, hydrogels were prepared in plastic Petri dishes (Ø 5 cm) from dry alginate layers derived from 1% alginate sol, and the gelation reaction was carried out at RT for 24 h.
Composition of crosslinking solutions
Alginate hydrogels were prepared through ionic crosslinking, and an aqueous calcium chloride solution was used as a source of Ca2+ cations. The concentration of CaCl2 in the crosslinking solutions ranged from 0 to 10 mM. The pH values of the crosslinking solutions were adjusted to either 5.5 or 7.4 with the HEPES buffer solution. Some of the crosslinking solutions contained NaCl (150 mM, final). Our abbreviation scheme describes the characteristics of the hydrogel's ionic composition in the crosslinking solution; for example, Ca-alginate and Ca-alginateNaCl hydrogels were generated with CaCl2 dissolved either in water or in 150 mM NaCl, respectively.
Measurements of alginate hydrogel swelling in gravimetric assay
The weight of each hydrogel (g) was measured on an analytical balance (Sartorius, Göttingen, Germany; 0.1 mg precision). To preserve gel integrity, the cumulative mass of a glass–gel sandwich was measured and the average weight of the glass coverslips (0.22 g) was subtracted to obtain the hydrogel's mass. At least three independent samples were prepared for every experimental condition. The Student's t-test for unpaired observations was applied to make a statistical analysis of the data. The rate of swelling was estimated after comparison of the hydrogel mass with the mass of the 1% alginate sol (taken as 100%).
Rheological characterization of alginate sols and hydrogels
Rheological measurements were carried out using an Anton-Paar Physica MCR301 (Graz, Austria) instrument. The viscosity values of aqueous alginate solutions were recorded under continuous logarithmically increasing shear in a double slit chamber DG26.7 at 20°C. A stress sweep at a constant frequency of 1 Hz was performed to obtain values for the elastic modulus (G′) of alginate hydrogels. Viscosity values were recorded with the help of a measurement cell with parallel plate geometry (PP25) under logarithmically increasing amplitude (0.001–100 Pa) at 37°C. Each hydrogel was transferred onto the measuring device and cut into the shape of a disk (Ø 25 mm). Samples were equilibrated at 37°C for 5 min before measurements.
Culturing of primary neuronal cells
Neuronal cell cultures were prepared from E20 Wistar rat fetuses. Cells were isolated from cortices as described. 48 As we reported previously, cleavage of the cell surface molecules during acute brain tissue dissociation using serine protease trypsin alters initiation of neural cell adhesion and growth on soft alginate hydrogels. 29 Therefore, in the present study, we used primary cell spheroids, which were completely recovered from treatment with trypsin. Primary cells were seeded at an initial cell density of 1×106 cells/well in uncovered six-well plates with 2 mL of expansion medium consisting of DMEM/F-12 (1:1) supplemented with 2 mM L-glutamine, 15 mM HEPES, 20 ng/mL of rhEGF (Sigma), and 10 ng/mL of rh-bFGF (Sigma). Subsequently, multiple spheroids formed after reaggregation or/and proliferation of cells on day 3 in vitro. Spheroids were collected, replated onto hydrogel samples, and cultured in a differentiating medium to potentiate differentiation toward neural cells. The differentiating medium consisted of 2% B-27 supplemented neurobasal medium, 0.5 mM L-glutamine, and 1% ampicillin and 1% streptomycin solution. Cell incubation was carried out at 37°C in a humidified 95% air and 5% CO2 atmosphere.
Immunocytochemistry
The primary monoclonal mouse antibodies were anti-β-tubulin III, anti-GFAP, and anti-Tau-1 (from Chemicon, Schwalbach, Germany) and anti-nestin (R&D Systems, Wiesbaden-Nordenstadt Germany), and secondary antibodies were Alexa Fluor-488 chicken anti-mouse (Molecular Probes/Invitrogen). Labeling with antibodies was performed according to a standard procedure; Hoechst 33342 (10 μg/mL in TBS) was used to label the nuclei. Samples were examined under a fluorescence microscope (Axiovert 40; Zeiss, Jena, Germany) and confocal microscope (LSM-710; Zeiss).
Neurite-bearing spheroids, neurite length, and statistical analysis
Spheroids that were formed from 1×106 primary cortical cells were collected into a sterile tube containing a fresh portion (6 mL) of a differentiating medium, then mixed, and evenly distributed on top of three identical Ca-alginateNaCl hydrogels (e.g., Ca-alginateNaCl hydrogels derived from S-alginate crosslinked with a solution containing 2 mM of CaCl2, 150 mM of NaCl, and at pH 7.4) and cultured subsequently for 48 h. The diameter of spheroids ranged from 50 to 250 μm by day 5 in vitro. Spheroids that extended 10 or more neurite from at least a quarter of its surface contacting the hydrogel were defined as neurite-bearing spheroids; of note, the vast majority of neurite-bearing spheroids extend 50 or more neurite. Single cells as well as aggregates of cells (2–10 cells) were not considered during spheroid counting. The proportion of neurite-bearing spheroids was quantified after the examination of 50 random live spheroids cultured on top of three identical Ca-alginateNaCl hydrogels. Different types of hydrogels were derived either from S-alginate, LVG, or LVM alginates crosslinked at pH 5.5 or 7.4 with a solution containing different concentrations of aqueous CaCl2 (i.e., 2, 4, 6, 8, or 10 mM) and 150 mM of NaCl as described above. In each independent cell culture experiment, 30 different hydrogel types were generated and tested for their ability to support neurite outgrowth. Every hydrogel type was generated in triplicate; subsequently, in each independent cell culture experiment, spheroids were examined on 90 individual hydrogels. The final proportion of neurite-bearing spheroids on every hydrogel type was estimated from two independent cell culture experiments after the examination of 100 (in total) individual spheroids on six identical hydrogels. The distance of neurite extension from spheroids was quantified in two independent cell culture experiments 48 h after plating of spheroids on top of different types of soft Ca-alginateNaCl hydrogels generated using S-alginate, LVG, and LVM with 2 mM of aqueous CaCl2 at pH 7.4 as described above. Subsequently, 20 random spheroids were examined per one type of a hydrogel and the length of 20–50 random neurite per spheroid was quantified on live phase-contrast images (Zeiss Ph2 Plan-NEOFLUAR 20X/0.5 objective, Axiovert 40; Zeiss) using NeuronJ (
Results
Swelling of Ca-alginate hydrogels in aqueous CaCl2 solution
Swelling is a result of liquid absorption by the polymer, and could be estimated by measuring the hydrogel mass. Dry alginate polysaccharide chains swell in water to produce a sol, but they can swell in the presence of crosslinking di- or multivalent cations, which lead to a hydrogel. Crosslinking of surface-attached dry alginate layers by a 2 mM aqueous CaCl2 solution resulted in the interconnection of polysaccharide molecules and the formation of a single gel slice (soft hydrogel), which support robust neurite outgrowth in vitro; in contrast, gelation of the alginate sol in an aqueous solution under gentle stirring generated multiple gel fragments of irregular size. The appearance of the soft hydrogels (e.g., fragments or slices) did not alter their high adhesiveness to neurite. These results matched our previous data 29 and demonstrated that dry alginate layers could efficiently form soft hydrogels with the ability to support robust neural growth.
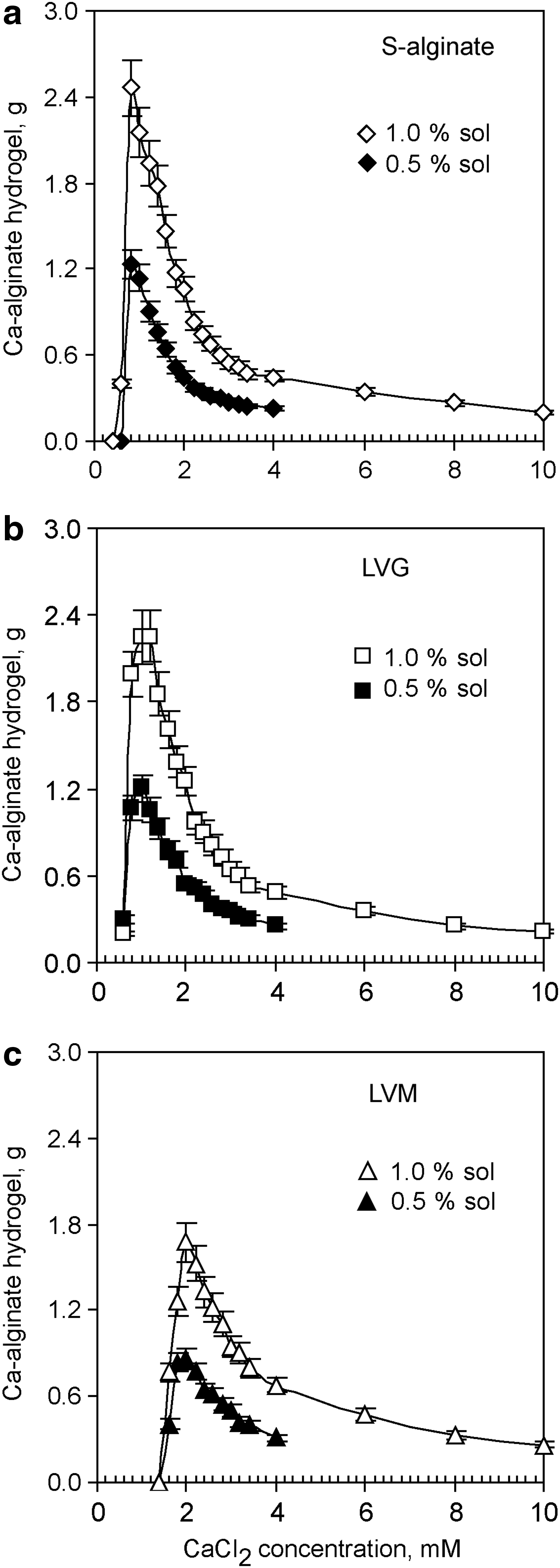
While cultivating neurons on top of soft hydrogels, we often observed variability in the long-term hydrogel stability: some hydrogels disintegrated in the presence of a cell culture medium during the first 3 days, but others persisted for up to 10 weeks and longer. In this study, we generated hydrogels from surface-attached dry layers of different alginates and tested how changes of the ionic strength and pH during the crosslinking influenced mechanical properties of hydrogels. At a constant concentration of calcium (2 mM of CaCl2), pH 5.5 and either in the absence or presence of NaCl (150 mM) during the crosslinking, the gel mass was proportional to the sol volume (Fig. 1a) and increased linearly with increased alginate concentrations in the sol (Fig. 1b). In the concentration range from 0–10 mM of CaCl2 in the crosslinking solution, the amount of Ca2+ had a nonlinear effect on the growth of a gel mass. A slow linear increase of the hydrogel's mass was correlated with a subsequent decrease in CaCl2 concentration from 10 to 4 mM, but a much more rapid increase of hydrogel mass was detected after gelation with ≤4 mM of CaCl2. The formation of a single interconnected hydrogel slice was abolished after Ca2+ reached its minimal gel-forming concentration. This swelling profile was observed for S-alginate, LVG, and LVM (Fig. 2). Moreover, the minimal gel-forming concentration of CaCl2 decreased with an increase of G-content in the alginate, and the level was found to be 0.8±0.2, 1.0±0.2, and 2.0±0.2 mM for S-alginate, LVG, and LVM, respectively (Fig. 2). In the presence of substoichiometric concentrations of calcium ions, larger networks formed when using highly viscous alginates such as LVG and S-alginate (145 and 132 mPa) than when using less viscous LVM (21 mPa) (Fig. 2). The differences in the magnitude of swelling between G-enriched (LVG) and M-enriched (LVM) alginates could be explained by the number of gelling points and their distribution in the primary alginate sequence and by the viscosity of alginates. Indeed, the affinity to bind Ca2+ increases with a greater number of G-blocks present in the polymer. 6 On the other hand, alginate viscosity is proportional to polysaccharide chain length, and high-viscosity alginates can form larger hydrogels because of the higher probability of crosslinking of the polysaccharide chains and physical entanglement. 7
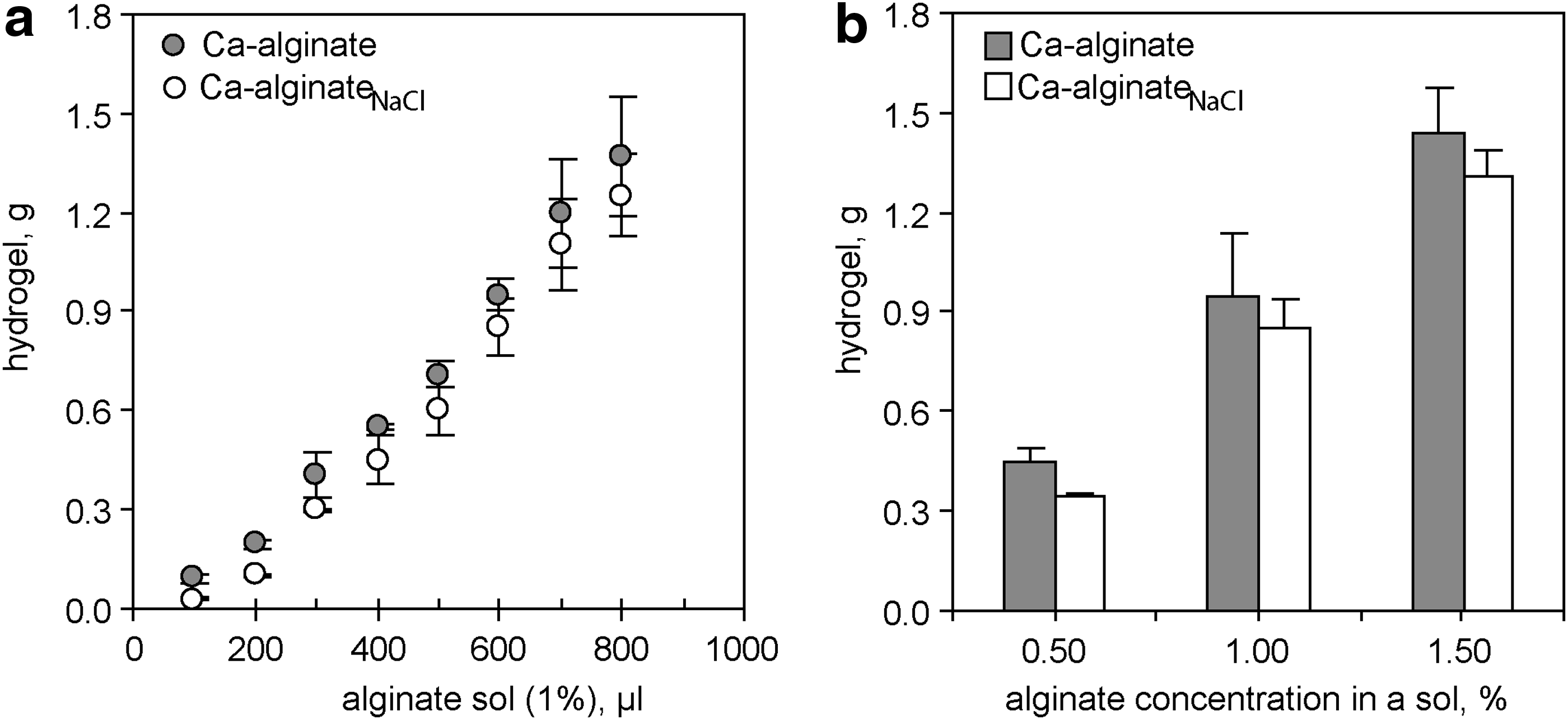
Hydrogel volume increases with an increase in
Swelling of the Ca-alginate hydrogel increases with decreased CaCl2 concentrations in the crosslinking solution. Hydrogels were generated at pH 5.5 by crosslinking dry alginate layers that originated from 600 μL of a 1% sol or 0.5% sol of
The dissociation constants for mannuronic and guluronic acid monomers are 3.38 and 3.65, respectively. In aqueous solutions with pH 5–9, alginate behaves as a polyanion because carboxylic groups in sodium alginate are fully dissociated. 7 In agreement with this fact, we found that the Ca-alginate hydrogel's swelling profile and the minimum gel-forming concentration of calcium ions were not influenced by the pH value (5.5 or 7.4) during the crosslinking (Figs. 2 and 3).

NaCl restricts the swelling of hydrogels. Hydrogels were generated at pH 7.4 by crosslinking dry alginate layers with aqueous CaCl2 solutions in the absence (Ca-alginate) or in the presence of 150 mM of NaCl (Ca-alginateNaCl); alginate layers originated from 600 μL of 1% sol of
Swelling of Ca-alginateNaCl hydrogels in aqueous CaCl2 solution
Like Ca-alginate hydrogels, hydrogels generated in the presence of NaCl (150 mM, Ca-alginateNaCl) displayed nonlinear swelling profiles in solutions with pH values of 5.5 (not shown) and 7.4, and a decrease in the CaCl2 concentration to below 4 mM was accompanied by a rapid increase in hydrogel mass (Fig. 3). At a constant concentration of CaCl2, the Ca-alginateNaCl hydrogel's mass was proportional to the sol volume and concentration of alginate in the sol (Fig. 1a, b). Na+ ions do not have any specific binding sites within the alginate, but interact electrostatically with the negatively charged carboxylic groups of the polysaccharide. 19 We observed that at a low degree of crosslinking (≤4 mM of CaCl2), exogenous Na+ ensures the generation of less swollen hydrogels (Fig. 3) and shifts the minimum gel-forming concentration of Ca2+ ions toward higher Ca2+ concentrations for G-enriched alginates (1.4 and 1.6 mM of CaCl2 in S-alginate and LVG, respectively). Thus, when a high Na+/Ca2+ ratio exists, Na+ could inhibit some G-enriched sequences in the primary alginate structure from being crosslinked by Ca2+, but does not alter the fundamental principal of hydrogel formation with Ca2+ (CaCl2≤10 mM). Moreover, as we demonstrated below, only strongly swollen Ca-alginateNaCl hydrogels with the degree of swelling above 66% supported robust neurite outgrowth in vitro.
Mechanical stability of Ca-alginateNaCl and instability of Ca-alginate hydrogels in buffered physiological saline
Sodium is a constituting ion of every extracellular medium in the body, and it contributes to the high ionic strength of extracellular fluids, including blood plasma and cerebrospinal fluid. Moreover, the pH value in normal blood and cerebrospinal fluid is tightly regulated between 7.35 and 7.45.49,50 To predict how long different in situ prepared hydrogels can maintain their dimensions in the presence of these physiological fluids in vivo (e.g., as implants into injured spinal cord) and subsequently support neural growth and regeneration, we quantified hydrogel disintegration in a buffered physiological saline. Ca-alginate and Ca-alginateNaCl hydrogels generated in the presence of 2 mM of CaCl2 were immersed in buffered physiological saline (150 mM of NaCl, pH 7.4) supplemented with 2 mM of CaCl2 and the hydrogel mass was regularly quantified during the subsequent 30 days. Ca-alginate hydrogels derived from LVM completely collapsed, while those derived from LVG and S-alginate lost ≥50% of their mass within the first 30 days (Fig. 4a). Surprisingly, analogous Ca-alginateNaCl hydrogels demonstrated long-term mass stability during the entire observation period (Fig. 4b).
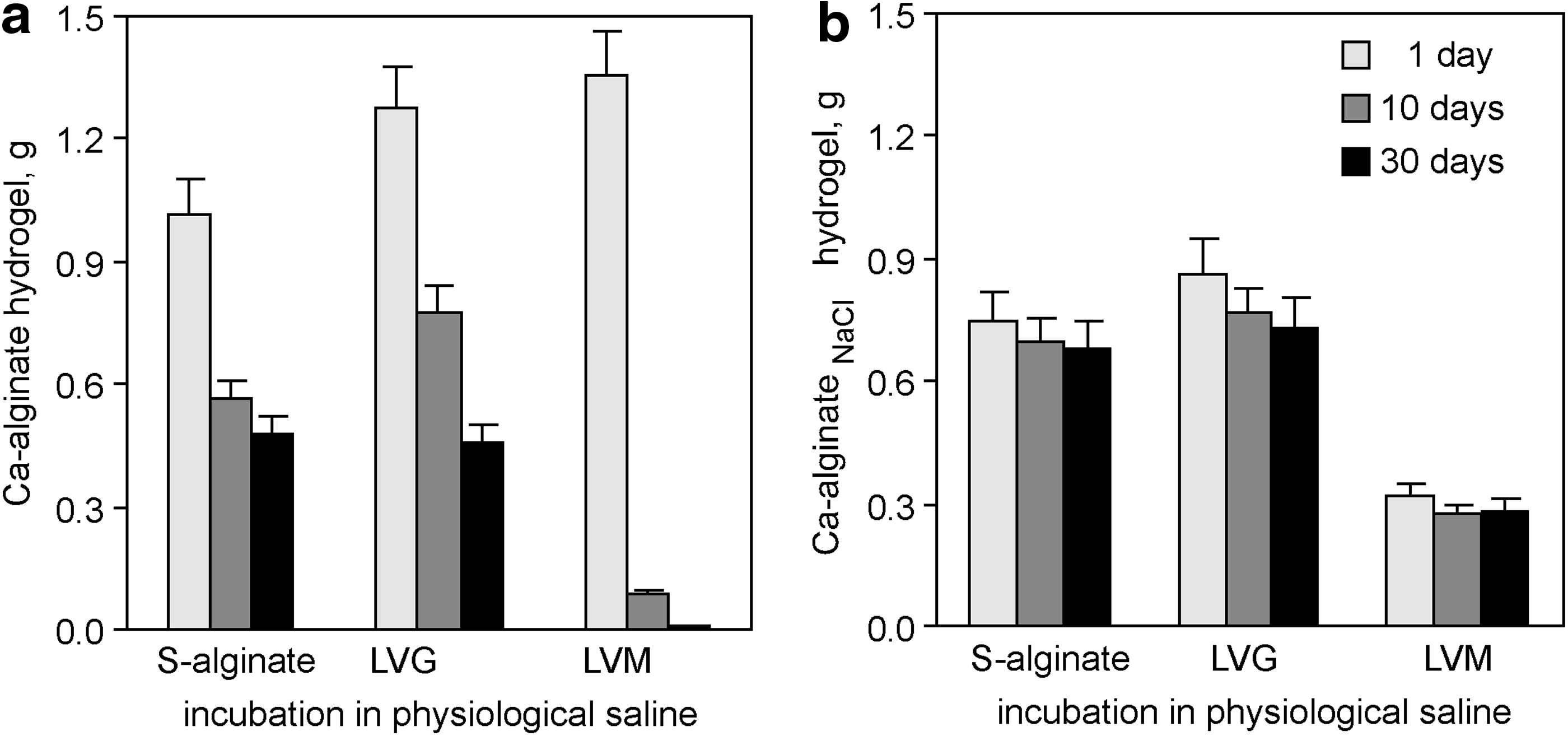
Ca-alginateNaCl hydrogels demonstrate prolonged volume stability in buffered physiological saline. Ca-alginate and Ca-alginateNaCl hydrogels were generated at pH 5.5 from dry alginate layers in the presence of 2 mM of CaCl2
Macroscopic morphology of hydrogels
Rapid binding of calcium ions to alginate produces macroscopically inhomogeneous structures; conversely, slow gelation kinetics, which commonly occur in the presence of components that compete with alginate for Ca2+ complexation such as oligoguluronates and EDTA, lead to more homogeneous gels.12,51 We noticed remarkable differences in the surface profiles between Ca-alginateNaCl and analogous Ca-alginate hydrogels. Many of the Ca-alginateNaCl hydrogels had a smooth and uniform surface, whereas Ca-alginate gels had ruffles and creases of irregular shape and pattern (Fig. 5). The differences in gel appearance were independent of alginate type and became more apparent in gels generated with solutions containing ≤3 mM of CaCl2 (Fig. 5f, g). Thus, at a high Na+/Ca2+ ratio, Na+ could delay Ca2+-mediated chain association, which optimizes the spatial organization of the alginate network.
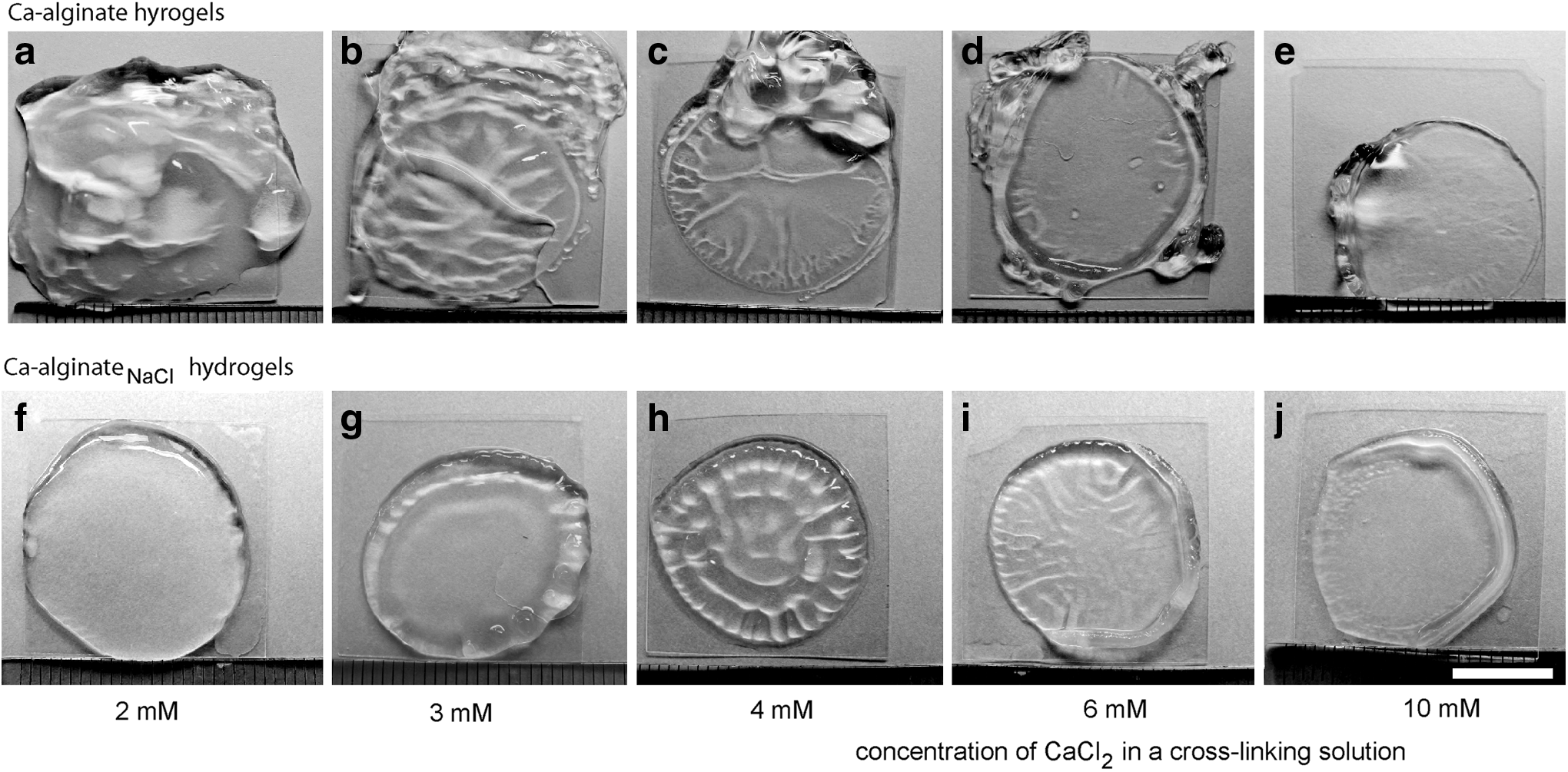
Phase-contrast images of Ca-alginate and Ca-alginateNaCl hydrogels. Hydrogels were generated at pH 7.4 by crosslinking dry alginate layers of S-alginate with 2–10 mM of aqueous CaCl2 solutions
Rheological characterization of Ca-alginateNaCl hydrogels
We carried out oscillatory shear measurements in a rheometer and quantified the stiffness of the Ca-alginateNaCl hydrogels. The stiffness of all tested hydrogels increased with higher levels of Ca2+ in the crosslinking solution. In addition, LVM-derived gels were softer than analogous S-alginate- and LVG-derived gels (Fig. 6). For example, the average stiffness of S-alginate- or LVM-derived Ca-alginateNaCl hydrogels formed in the presence of 2, 4, 8, and 10 mM of CaCl2 were 0.011, 1.470, 3.665, and 19.230 kPa (S-alginate) and 0.025, 0.622, 3.085, and 7.356 kPa (LVM), respectively.
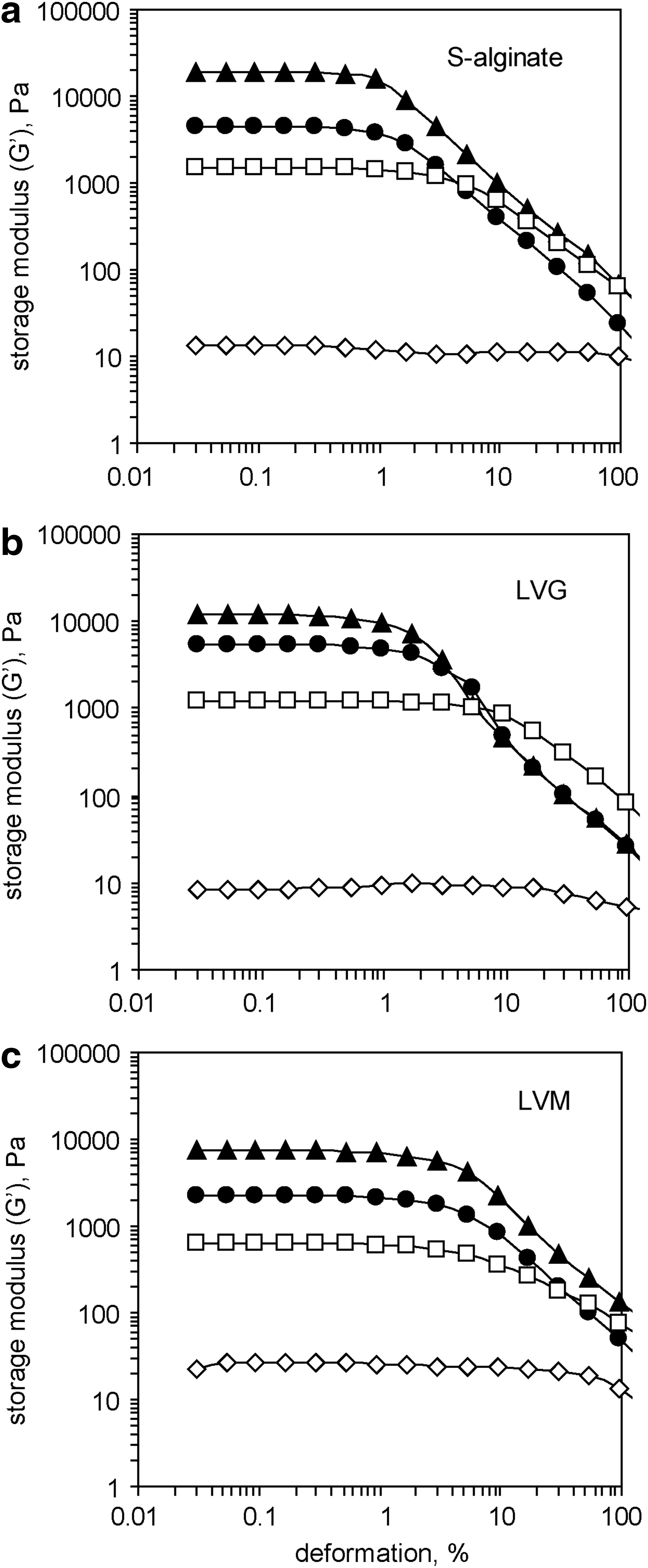
Increase in the storage modulus (G′) of a Ca-alginateNaCl hydrogel correlates with an increase in the concentration of CaCl2. Hydrogels were generated at pH 5.5 by crosslinking dry alginate layers of
Neurons rapidly extend neurite on soft Ca-alginateNaCl hydrogels
Neurons have an intrinsic capacity to extend neurite on surfaces they can adhere to. Previously, we described that neurons extend neurite on soft alginate hydrogels prepared with 2 mM of Ca2+, but gelation with 10 mM of Ca2+ resulted in a stiff and nonadhesive hydrogel; this change was attributed to increased tightness of the alginate network. 29 We tested whether Ca-alginateNaCl hydrogels are still capable of promoting neural adhesion and growth under growth factor-free conditions and without serum. An array of hydrogels were generated from LVM, LVG, and S-alginate; crosslinking was performed in the presence of a constant concentration of NaCl (150 mM) and different concentrations of CaCl2 (i.e., 2, 4, 6, 8, or 10 mM) at pH 5.5 or 7.4.
As we reported previously, the ability of a soft alginate hydrogel to promote neurite extension did not depend on the cell culture system because two-dimensional (2D) adherent monolayers of neurons and 3D neural spheroids produced identical results.
29
Thus, 3D spheroids were chosen for the cell culture experiments in this study. Spheroids consisted of living cells, which neither accumulated trypan blue dye nor contained fragmented nuclei (data not shown). Three hours after being plated on soft Ca-alginateNaCl hydrogels (S-alginate, LVG, and LVM gelled with 2 mM of CaCl2), multiple neurite extended in all directions and away from the center of the spheroids. The spheroid-derived neurite expressed the neural markers MAP2 and β-tubulin III (Fig. 7a, e, i and b, f, j) and developed clearly defined growth cones (Supplementary Fig. S1; Supplementary Data are available online at

Soft Ca-alginateNaCl hydrogels support neurite outgrowth from primary cortical neurons. Fluorescence images show
In agreement with data in the literature, we observed that multiple nestin- and GFAP-labeled cells migrated from the spheroids on poly-l-lysine-coated surfaces to form an adherent 2D cell monolayer (not shown). In contrast, spheroids preserved their 3D shape on alginate hydrogels, and only neurons extended neurite on the top of the soft hydrogel (Fig. 7a, b, e, f, i, j). Unlike neurons, GFAP-positive cells did not extend their processes outside spheroids neither on soft nor on stiff Ca-alginateNaCl hydrogels.
Since soft Ca-alginateNaCl hydrogels can be shaped to take any form or size and do not collapse during gentle mechanical treatment, they can be implanted in vivo by direct placing of a piece of hydrogel into a tissue lesion (e.g., lesion formed after traumatic spinal cord or brain injury). Biocompatible, soft Ca-alginateNaCl hydrogels promote neural growth and could be tested in the future as neural bridges in vivo.
Discussion
This study describes soft Ca-alginateNaCl hydrogels as stable 3D matrices with high adhesiveness to neural cells. Within a pH range that support dissociation of sodium alginate in aqueous solutions, fluctuations in the pH or ionic strength of the solution and drying of the alginates did not affect neural growth on/within the hydrogel, but gelation in the presence of NaCl enabled the generation of mechanically stable and soft 3D matrices with mechanical compliance close to brain tissue (≤1.5 kPa) that could be tested in the future as neural bridges in vivo.52–54
Mechanical compliance of the substrate determines many aspects of cellular behavior, cell morphology, and gene expression.27,55 We previously demonstrated that soft, but not stiff alginate hydrogels were able to create an adhesive matrix for primary neurons. Now, we have shown that gelation in the presence of NaCl enhanced the mechanical stability of soft hydrogels, but did not alter the capacity of the resulting hydrogels to support neural growth. As described for pure Ca-alginate hydrogels, neurons prefer soft over stiff substrates; for example, the soft Ca-alginateNaCl hydrogels with elastic moduli between 0.01–1.5 kPa supported neurite extensions, while neurons were unable to adhere or spread neurites onto stiff Ca-alginateNaCl hydrogels, which have elastic moduli above 3 kPa.
There is strong auto-cooperative binding of calcium and other di- or multivalent cations by the alginate polysaccharide chains in the gel state. 6 Subsequently, our in vitro neurite outgrowth assay revealed a sharp and noncontinuous transition from adhesive to nonadhesive states for Ca-alginateNaCl hydrogels that was observed between 4 and 6 mM of CaCl2; these qualitative changes were pH independent and were detected in hydrogels derived from both G-enriched and M-enriched alginates with viscosities ranging from 21 to 145 mPa.
Polymers behave differently on surfaces than in solutions.15,16 Nevertheless, our results obtained for hydrogels derived from surface-attached alginate layers are largely in agreement with the egg-box model, which was developed for alginate hydrogels in general. We observed that (1) the swelling profiles and stiffnesses of hydrogels fabricated in the presence or absence of NaCl were controlled by the concentration of Ca2+ and (2) high-viscosity G-enriched alginates required less Ca2+ to form a gel and eventually created larger networks than low-viscosity M-enriched alginates. Alginate concentrations directly influenced the hydrogel mass, but had no noticeable effects on the hydrogel swelling profile; therefore, manipulation of the amount of alginate in a layer could be used to fine tune the hydrogel's dimensions (i.e., to generate a thin matrix to culture neurons or a thick scaffold for implantation in vivo).
The acidic form of alginate is not water soluble, but its sodium salt is. In water, sodium alginate dissociates into negatively charged polymer chains, and mobile sodium ions that form a loosely bound counterion cloud around negative charges assure the polymer's water solubility and cause a decrease in intrachain repulsion. Depending on the charge distribution along the chain, polymers could exist either in globular or persistent configurations. Subsequently, hydrogels formed by the association of globular polymer chains occupy more space, while hydrogels formed by persistent polymers are more compact. Additionally, slow gelation kinetics result in macroscopically homogeneous alginate gels.12,15 Thus, the macroscopic homogeneity and compactness of Ca-alginateNaCl hydrogels suggest that exogenous Na+ could enable slow gelation kinetics and optimize the spatial organization of an alginate network.
One Ca2+ ion binds up to four guluronic acid residues. 21 In firmly connected, soft Ca-alginate hydrogels (e.g., in hydrogels derived from 1% LVG sol and 2 mM of CaCl2, where ∼12% of the G residues have the potential to be crosslinked), the number of endogenous Na+ counterions are not sufficient to neutralize the negative charges on the polysaccharide chains, and repulsion between non-cross-linked sequences results in the formation of extremely swollen hydrogels. After immersion of such soft Ca-alginate hydrogels into physiological saline, exogenous Na+ ions penetrate into the hydrogel and, presumably, reduce intrachain repulsion to trigger global 3D changes in the entire network and leakage of Ca2+ ions from gelling sites, which lead to the eventual collapse of the hydrogel. In contrast, soft neurite-supportive Ca-alginateNaCl hydrogels did not collapse after infiltration of exogenous Na+ and maintained their mass and volume after immersion in buffered physiological saline, which makes them interesting for in vivo applications.
Polysaccharides (i.e., sugars) decorate proteins within the ECM and have been found to influence cell migration, axonal guidance, synapse development, and functioning.56,57 Most recently, wobble oligosaccharide motifs of chondroitin sulfate/dermatan sulfate proteoglycans had been demonstrated to induce signaling pathways that are essential for the proliferation, self-renewal, and cell lineage commitment of neural stem cells. 58 Although stiff alginate hydrogels were described as an inert matrix, alginate-derived oligosaccharides have been reported to costimulate growth factor-mediated proliferation, migration, and adhesion of endothelial cells, keratinocytes, and skin fibroblasts in vitro;59–61 cytokine production by RAW264.7 cells; 62 and phagocytic activity of peritoneal monocytes. 63 The density of substrate-immobilized signaling molecules is crucial to initiate extracellular signaling events that lead to cell differentiation, adhesion, growth, and migration, and this must be considered during scaffold designing.64,65 In light of these reports, we hypothesize that firmly crosslinked alginate networks (and, therefore, soft hydrogels) could create a matrix with a high density of immobilized, adhesion-promoting carbohydrate sequences.
Identification of the molecular mechanisms involved in neural growth on alginate matrices could provide valuable information for regenerative medicine and open new perspectives on using alginate polysaccharides for the functionalization of nonadhesive matrices.
Footnotes
Acknowledgments
This study was funded by the German Research Foundation (DFG).
The authors wish to acknowledge Prof. Helmut Kettenmann from Max-Delbrück-Center for Molecular Medicine Berlin-Buch, Berlin, Germany, for fruitful discussions, Fabian Luthardt from the Fraunhofer Institute for Ceramic Technologies and Systems (IKTS), Dresden, Germany, for assistance with the rheological measurements, and Dr. Komber from Leibniz Institute of Polymer Research, Dresden, Germany, for 1 H-NMR spectroscopy.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.