
Abstract
The incidences of skin cancers resulting from chronic ultraviolet radiation (UVR) exposure are on the incline in both Australia and globally. Hence, the cellular and molecular pathways that are associated with UVR-induced photocarcinogenesis need to be urgently elucidated, in order to develop more robust preventative and treatment strategies against skin cancers. In vitro investigations into the effects of UVR (in particular, the highly mutagenic UVB wavelength) have, to date, mainly involved the use of cell culture and animal models. However, these models possess biological disparities to native skin, which, to some extent, have limited their relevance to the in vivo situation. To address this, we characterized a three-dimensional, tissue-engineered human skin equivalent (HSE) model (consisting of primary human keratinocytes cultured on a dermal-derived scaffold) as a representation of a more physiologically relevant platform to study keratinocyte responses to UVB. Significantly, we demonstrate that this model retains several important epidermal properties of native skin. Moreover, UVB irradiation of the HSE constructs was shown to induce key markers of photodamage in the HSE keratinocytes, including the formation of cyclobutane pyrimidine dimers, the activation of apoptotic pathways, the accumulation of p53, and the secretion of inflammatory cytokines. Importantly, we also demonstrate that the UVB-exposed HSE constructs retain the capacity for epidermal repair and regeneration after photodamage. Together, our results demonstrate the potential of this skin equivalent model as a tool to study various aspects of the acute responses of human keratinocytes to UVB radiation damage.
Introduction
T
Chronic UVA and UVB exposure gives rise to cutaneous immunosuppression and DNA damage, which together form the intrinsic pathways to NMSC photocarcinogenesis. 7 The UVA wavelength penetrates into the dermis of the skin and is predominantly responsible for the photoaging effects of sun exposure. 8 Considered the major causative factor in epidermal malignancies, UVB is known to be highly destructive to keratinocytes via direct DNA damage. 9 UVB irradiation leads to the formation of DNA photolesions, predominantly cyclobutane pyrimidine dimers (CPDs) and (6–4) pyrimidine-pyrimidones, along with the generation of DNA-damaging reactive oxygen species. 10 Defence mechanisms against cutaneous photocarcinogenesis include the activation of cell cycle arrest and the subsequent initiation of the nucleotide excision repair complex. 11 Failure to correctly and completely repair UVR-induced DNA damage, however, can lead to the accumulation of signature mutations, such as C→T transitions, commonly found in UVR-induced skin tumors. 12 Higher doses of UVB radiation trigger apoptosis (or the formation of sunburn cells), yet another cellular safety net to inhibit the replication of genetically damaged proliferative keratinocytes. 13 The intricacies of keratinocyte responses to UVB, and in particular the processes surrounding NMSC formation, however, have yet to be fully mapped. Investigations into the biological reactions after UVB radiation exposure using human participants have several ethical constraints. These are attributed to the considerable risks involved with UVR exposure and the requirement of skin biopsy collection, which together have limited such studies. Hence, physiologically relevant in vitro experimental platforms that are capable of accurately modeling the human UVB epidermal response may represent a practical and accessible alternative to such in vivo studies.
A variety of animal, cell, and tissue culture-based models have been employed to investigate UVB-induced damage and repair in keratinocytes. 14 Drawbacks associated with these techniques, however, include strict ethical regulations, significant costs, and inconsistencies related to structural and biological aspects of native skin.15–17 The establishment of three-dimensional (3D), tissue-engineered, organotypic human skin equivalents (HSEs) has addressed some of these limitations.15,18,19 Essentially, the construction of HSEs involves the culture of epidermal skin cells on natural or synthetic dermal substrates. These HSEs may then be cultured at the air-liquid interface in order to promote keratinocyte differentiation and epidermogenesis.20–22 This results in the formation of an epidermal layer that is histologically similar to native human skin. 21 These tissue-engineered constructs serve as versatile and powerful research tools with numerous applications, such as the study of cancer, pigmentation, and toxicity. 15
In this study, we characterize the novel application of a human keratinocyte-based HSE using a de-epidermized dermal (DED) culture substrate as a tool to investigate keratinocyte responses to UVB radiation. Importantly, compared with conventional two-dimensional (2D) keratinocyte cultures, the HSE-DED more closely recapitulates the in vivo tissue structure and microenvironment.23,24 The use of synthetic dermal substrates such as collagen, 25 fibrin, 26 and chitosan-gelatin-hyaluronic acid 27 in HSE construction has shown mixed results in terms of keratinocyte attachment and differentiation.28,29 Moreover, countries such as Australia tightly regulate the import of commercially available HSE composites for research purposes. 24 Hence, the aim of this study was to validate the HSE-DED model as an accessible and biologically compatible platform to study UVB radiation-induced damage and postirradiation repair in keratinocytes.
This HSE-DED model has been established in our laboratory as a tool to study wound healing and re-epithelialization processes.20,21 Here, we describe an additional application of the HSE-DED to characterize the epidermal UVB photoresponse in vitro. Specifically, the HSE-DED was validated in terms of (1) the expression of key epidermal differentiation markers compared with native skin; (2) the generation of characteristic markers of UVB photodamage; and (3) the capacity for postirradiation repair and regeneration at cellular and epidermal tissue levels.
Materials and Methods
Isolation and culture of human keratinocytes
Human keratinocytes were isolated from skin tissue that was obtained from surgical discard as previously described. 30 Briefly, skin was washed in three serial washes with antibiotic-antimycotic solution (Invitrogen) that was supplemented with gentamicin sulfate. Skin tissue was cut into ∼3×3 mm pieces and incubated in 0.125% trypsin solution at 4°C overnight. After enzymatic digestion, the epidermis was detached from the dermis using forceps. Cells at the epidermal-dermal junction were scraped off gently and resuspended in Full Green's media (FGM) as previously described by Rheinwald and Green.24,30,31 Freshly isolated epidermal cells were then seeded into a T75 cell culture flask containing 2×106 irradiated 3T3 mouse fibroblasts at a ratio of 1:1. Keratinocyte cultures were maintained at 37°C with 5% CO2 until 80% confluency was reached. Cell culture media (FGM) was replaced every 2–3 days. Nonsun exposed skin was obtained from consenting donors undergoing elective cosmetic breast reduction or abdominoplasty procedures. Ethical approval for the collection of human tissue was obtained from ethics committees at participating hospitals and the Queensland University of Technology (St Andrew's Hospital; Approval #200 4/46, Brisbane Private Hospital and Princess Alexandra Hospital; Approval #QUT 3865H). All experimentation performed with donor tissue was in accordance with the Declaration of Helsinki, 2004.
Preparation and culture of HSE-DED
The HSE-DED composites were prepared using a previously reported method. 22 Briefly, the DED scaffold was prepared from 1.4 cm2 pieces of the dermis of ex vivo skin tissue according to previously described protocols22,23 by immersion in 1 M sodium chloride solution and incubated at 37°C overnight to decellularize the tissue. The epidermis was then separated from the dermal tissue and discarded. Next, ∼1.4 cm2 pieces of DED were prepared in 24-well cell culture plates (Thermo Fischer Scientific). The resulting DED was then washed with phosphate-buffered saline before being immersed in FGM overnight at 37°C before keratinocyte seeding. Cultured keratinocytes (passage 1, 2×104 cells) were then seeded (in sterile stainless-steel rings with silicone washer bases) (Aix Scientifics) onto the papillary surface of each prepared DED. The DED was then incubated with the rings in place at 37°C with 5% CO2 for 48 h before their removal. The HSE-DED composites were then cultured in FGM on stainless-steel grids at the air-liquid interface in six-well cell culture plates (Thermo Fischer Scientific) at 37°C with 5% CO2, for 9 days. 21
UVB-irradiation of skin tissues
Skin tissues (HSE-DEDs and ex vivo skin explants) were irradiated with a single dose of UVB radiation using a calibrated medical UVB bulb (TL-40; Phillips) that was calibrated to deliver 9.27 mJ/cm2/min UVB radiation, with a stabilized power supply under sterile conditions in the dark at room temperature. Cellulose acetate film (Kodacel TA-407; Eastman-Kodak) was used to block out contaminating radiation (wavelengths below 290 nm) from the UVB source. 32 The HSE-DED constructs were partially submerged in prewarmed PBS during irradiation. In the irradiated samples, the epidermal surface of HSE-DED constructs was placed 6 cm from the external surface of the UVB lamp and exposed to 50 mJ/cm2 UVB (two minimum erythemal dose) radiation. Irradiation was conducted inside a Class II Biological Safety Cabinet to maintain sterility. Mock irradiated samples were treated with similar conditions but in the absence of UVB radiation. The UVB irradiation dose was chosen based on dose-response studies in which 50 mJ/cm2 UVB generated classic markers of UVB radiation-induced damage without complete destruction of the HSE-DED epidermal layer (data not shown). The skin equivalent constructs were returned to the culture conditions as described in the previous section postirradiation. Ex vivo skin explant tissues that were used for a comparative analysis were UVB irradiated within 2 h of excision from the donor patient. The skin explant tissues were maintained in FGM on stainless-steel grids at the air-liquid interface in six-well cell culture plates (Thermo Fischer Scientific) at 37°C with 5% CO2 until analysis.
Histological analysis
At specific time points postirradiation, HSE-DED composite tissue was fixed in 10% neutral buffered formalin overnight at 4°C, before being dehydrated in a graded ethanol series and paraffin embedded. Sections of 5 μm thickness were cut (from the center of the HSE-DED samples) and mounted onto glass slides. The tissue sections were deparaffinized in xylene and rehydrated in a decreasing graded ethanol series before routine hematoxylin and eosin (H&E) staining.
Immunohistochemical analysis
The immunohistochemical analyses of the HSE-DED tissue sections were performed as per previously documented techniques. 22 Briefly, 3 μm-thick tissue sections were deparaffinized and rehydrated before heat-induced epitope revival treatment in either tris-ethylendiaminetetraacetic acid (EDTA) buffer at pH 9.0 or sodium citrate buffer at pH 6.0 for 4 min at 97°C, followed by 10 min at 70°C in a decloaking chamber. Endogenous peroxidases were blocked with hydrogen peroxide solution (Biocare Medial), and nonspecific primary antibody binding was blocked using a blocking reagent (Biocare Medical). The primary antibodies used in immunohistochemical analysis are detailed in Table 1. Primary antibodies were diluted in sample diluent solution (Biocare Medical) and incubated in a humidified chamber overnight at 4°C. A horseradish peroxidase-linked secondary antibody was then applied to the tissue samples and detected with a 3,3′diaminobenzidine (DAB) chromogen system (Biocare Medical). The tissue sections were counterstained using hematoxylin Gill 1 (G1) nuclear stain (HD Scientific).
Detection of apoptotic cells
Tissue sections of UVB-irradiated HSE-DED composites were analyzed for the presence of apoptotic cells using an in situ terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay (Roche Applied Science) as per the manufacturer's protocol. Cell nuclei were counterstained with DAPI solution (Invitrogen) and imaged using fluorescence microscopy.
Quantification of secreted cytokines
To quantify levels of secreted cytokines in UVB-irradiated HSE-DED cultures, enzyme-linked immunoassay kits (Quantikine®; R&D Systems) for the detection of human IL-6 and IL-8 were utilized as per the manufacturer's instructions. Before irradiation, FGM was replaced with serum-free media (FGM without fetal calf serum) in the HSE-DED cultures. Cell culture supernatant samples were collected before and after UVB irradiation and were centrifuged to remove cellular debris. Samples were stored at −80°C until the assay was performed.
Image analysis
Digital image analyses of antibody-labeled tissue sections were performed using ImageJ software (public domain software downloaded from
Statistical analysis
One-way ANOVA with Tukey's post hoc tests (all group comparisons) was applied to analyze the data. Statistical significance was accepted when p<0.05.
Results
Epidermal characterization of the HSE-DED model compared with native skin
The human epidermis has a characteristic polarized pattern of keratinocyte growth, with actively proliferating cells situated at the dermal-epidermal junction. A comparative immunohistochemical study was conducted on unirradiated HSE-DED and native skin tissue to identify parallels in the expression of key epidermal markers (Fig. 1). Keratins 1, 10, and 11 were detected in terminally differentiated keratinocytes within the stratum spinosum, stratum granulosum, and stratum lucidum in both the HSE-DED and ex vivo skin sections. Upper spinous and granular keratinocytes, known to express cytokeratin 2e, were also detected in the HSE-DED composites.34,35 Similarly, the presence of keratin 14-expressing keratinocytes was detected in both native skin and HSE-DED sections, with higher immunoreactivity levels observed among the basal and suprabasal cell populations. Positive immunoreactivity of filaggrin was detected in the cornified envelope of keratinocytes within the stratum corneum in both the HSE-DED and skin tissue samples alike. The presence of type IV collagen, a major constituent of the BM zone in the skin, was detected at the dermal-epidermal junction in both the HSE-DED and skin tissue samples. Besides separating the epidermis and dermis, collagen IV is also known to line sweat glands, hair follicles, and blood vessels, which accounts for its positive immunoreactivity in these structures within the dermis and DED. 36
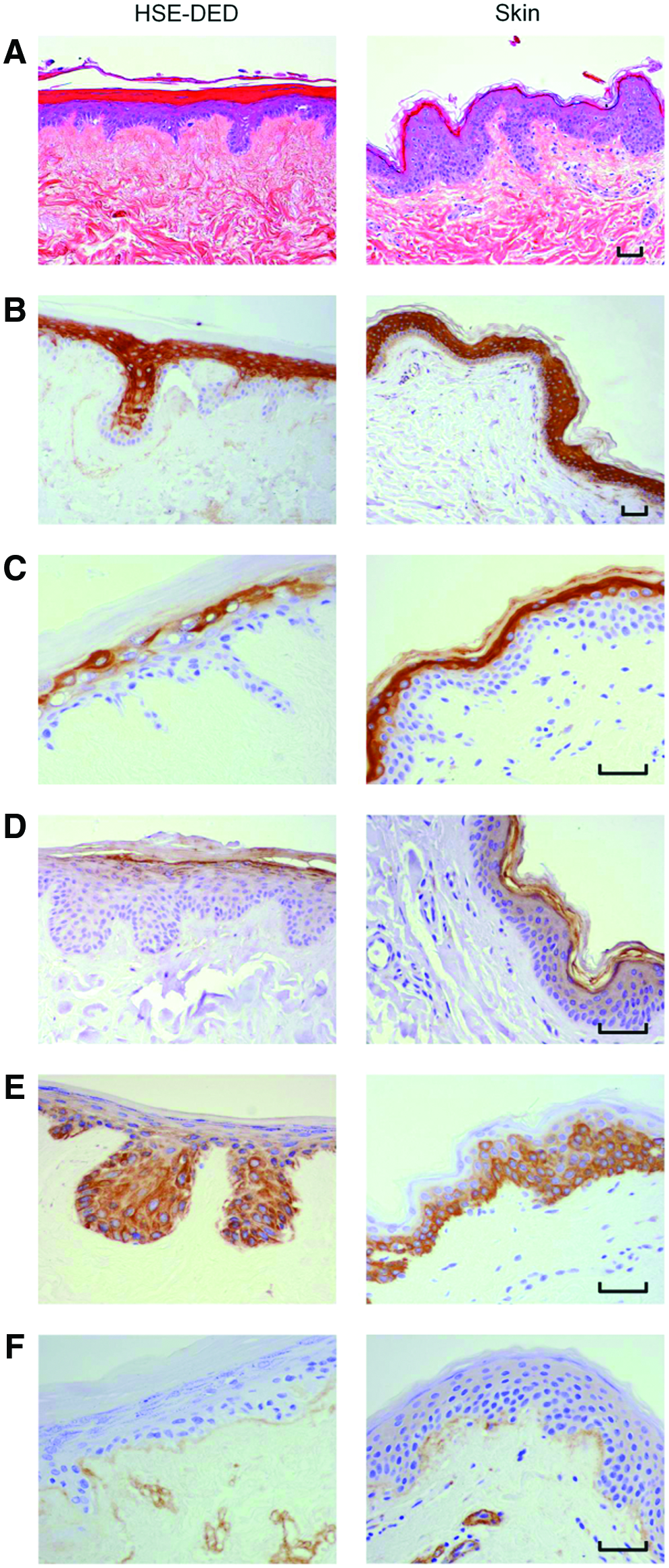
Characterization of unirradiated human skin equivalent-de-epidermized dermal (HSE-DED) constructs compared with native skin. Hematoxylin and eosin staining of HSE-DED and native skin sections
UVB irradiation induces markers of photodamage in the HSE-DED
A histological analysis of UVB-irradiated HSE-DED and ex vivo skin explant tissue revealed the formation of apoptotic keratinocytes in the epidermal layer, characterized by the presence of pyknotic nuclei and cytoplasmic shrinkage (Fig. 2A). The formation of CPD molecular lesions in the nuclei of UVB-irradiated keratinocytes is a classic biomarker of DNA damage and an important factor in NMSC development. 10 Immunohistochemical staining using an antibody that was specific for CPDs was performed on irradiated and nonirradiated HSE-DED tissue (Fig. 2B). CPD-positive keratinocytes were observed only in UVB-treated HSE-DED constructs, showing marked positive nuclear immunoreactivity in basal and suprabasal cells.
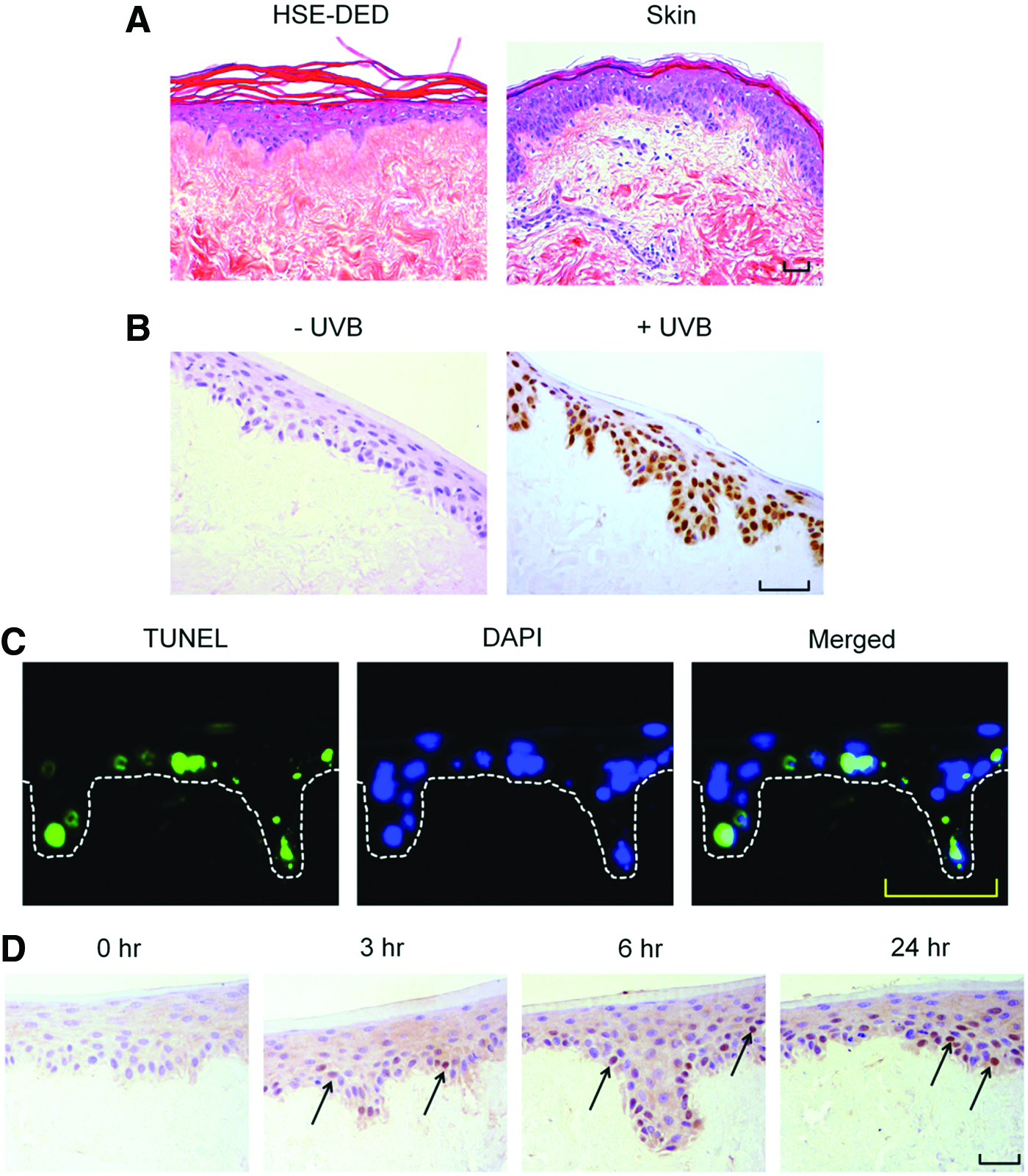
Analysis of UVB photodamage markers in irradiated HSE-DED constructs. Images represent the histology
Keratinocytes that have sustained irreparable DNA damage resulting from UVB damage undergo apoptosis to prevent the propagation of cells harboring photolesions. Using the TUNEL assay, DNA fragmentation (as a marker of apoptosis) was detected in UVB-exposed HSE-DED constructs (Fig. 2C). At 24 h after a single two MED UVB dose, cells undergoing apoptosis were found to be present along the stratum basale and among suprabasal keratinocyte populations. On average, 19.9%±4.1% of the total number of epidermal nuclei were found to be TUNEL positive at 24 h postirradiation.
The activation and downstream processes mediated by p53 are known to be critical for suppressing oncogenesis in UVB-irradiated keratinocytes. The presence of p53 in UVB-exposed HSE-DED keratinocytes was detected using immunohistochemical analysis (Fig. 2D). Sections from these HSE-DED constructs sacrificed at 0, 3, 6, and 24 h postirradiation reveal the intranuclear accumulation of p53.
The up-regulation of IL-6 and IL-8 production by UVB-irradiated keratinocytes enable them to be considered central catalysts for the activation of local and systemic immune responses.37,38 The relative levels of secreted IL-6 and IL-8 were quantified in the media of HSE-DED cultures before and after UVB exposure with an ELISA (Fig. 3). Increases in the production of both cytokines were observed in irradiated HSE-DED at the 24 h postirradiation time point compared with samples obtained at t=0 h. This includes a significant increase from 1.00±0.3 to 90.3±16.8 pg/mL in detected IL-6. Basal levels of IL-8 that were detected were relatively higher (24.0±1.0 pg/mL) due to this cytokine being constitutively expressed by keratinocytes. UVB exposure resulted in a slight increase in IL-8 in HSE-DED media, with an average of 66.1±5.7 pg/mL.

Quantification of cytokine secretion in HSE-DED culture supernatants. Cell culture medium samples were obtained from HSE-DED cultures before and 24 h after irradiation with two MED UVB radiation. Supernatants were analyzed for the presence of interleukin-6
HSE-DED composites retain the capacity for repair and regeneration after UVB irradiation
Due to the frequency of their exposure to damaging UVR, keratinocytes possess highly sophisticated repair mechanisms to cope with the formation of UVB-induced DNA mutations. 39 The efficient removal of CPDs, for instance, has been strongly associated with the prevention of oncogenic transformation in irradiated keratinocytes. 40 In view of this, an immunohistochemical analysis was performed to study the removal of UVB-induced CPDs in irradiated HSE-DED composites (Fig. 4A). After a single dose of UVB radiation, a gradual decrease in the intensity of CPD-positive immunoreactivity was observed in the epidermis of the HSE-DED over 5 days under culture conditions. At 2 to 5 days postirradiation, a significant decrease in the number of CPD-bearing keratinocytes was observed in these constructs as compared with levels immediately after UVB exposure (Fig. 4B).
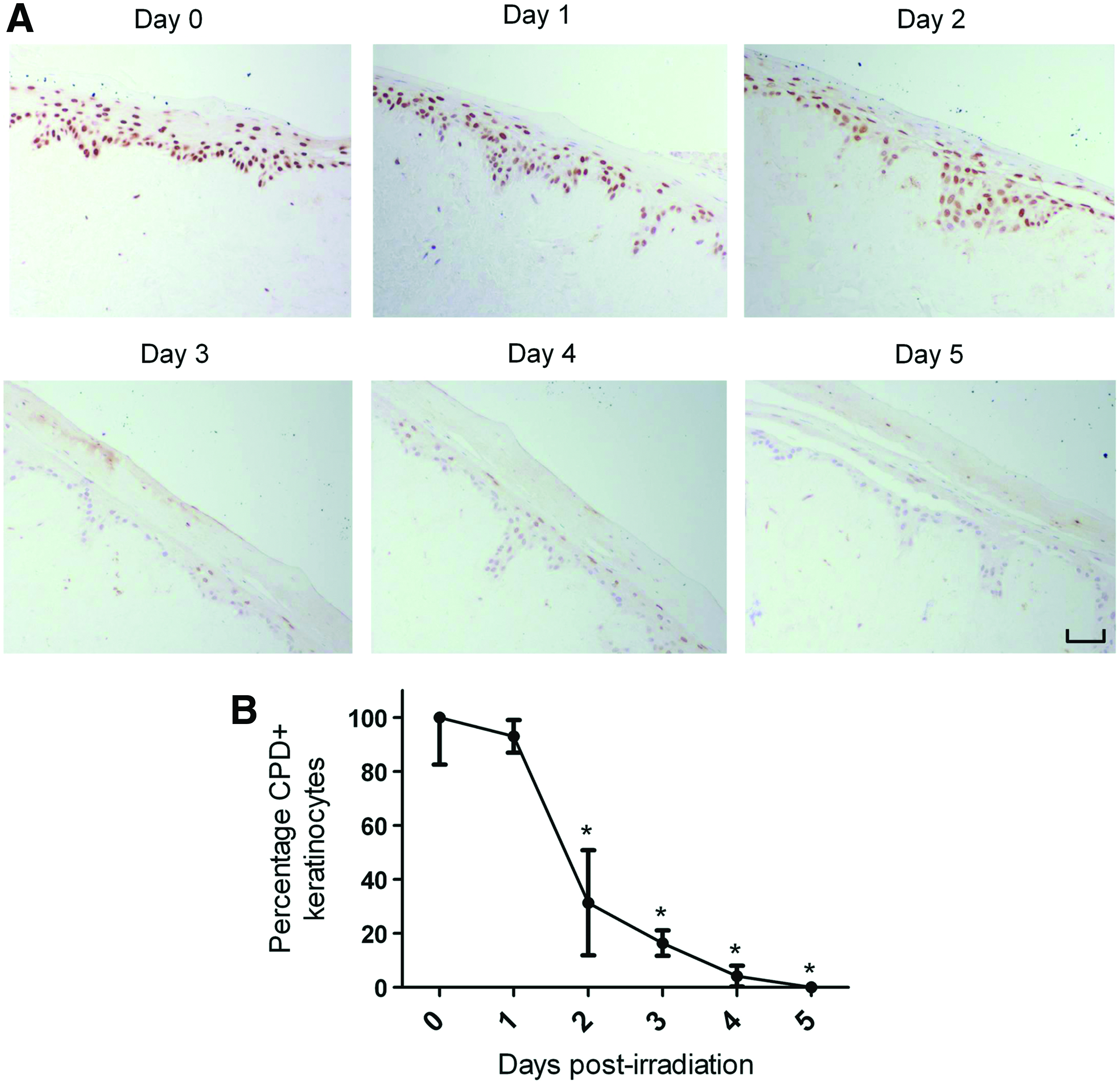
Postirradiation removal of UVB-induced CPDs in HSE-DED constructs. CPDs in the nuclei of HSE-DED keratinocytes irradiated with two MED UVB over 5 days were detected using immunohistochemical analysis
Epidermal regeneration after UVB-induced damage is facilitated by the stimulation of proliferation by various growth factors and cytokines. 41 The nuclear expression of the Ki-67 antigen occurs during all active phases of cellular proliferation. 42 Keratinocyte proliferation during the HSE-DED epidermal regenerative phase was investigated using immunohistochemical detection of Ki-67 in tissue sampled over 5 days postirradiation (Fig. 5). In UVB-exposed HSE-DED constructs, significantly lower numbers of Ki-67-expressing keratinocytes were observed at 1 and 2 days after irradiation (relative to unexposed HSE-DEDs). Subsequently, at 3 and 4 days post-treatment, there was a significant elevation in the number of proliferative keratinocytes in the UVB-treated constructs as compared with the untreated controls. Similar levels of Ki-67-positive keratinocytes were observed at Day 0 and 5 in both irradiated and nonirradiated samples. This indicates that UVB exposure triggers an initial inhibition of proliferation in the HSE-DED keratinocytes, potentially coinciding with the repair of DNA damage. This is followed by a proliferative burst that may function to replace UVB-damaged cells in which differentiation or apoptotic pathways have been activated.
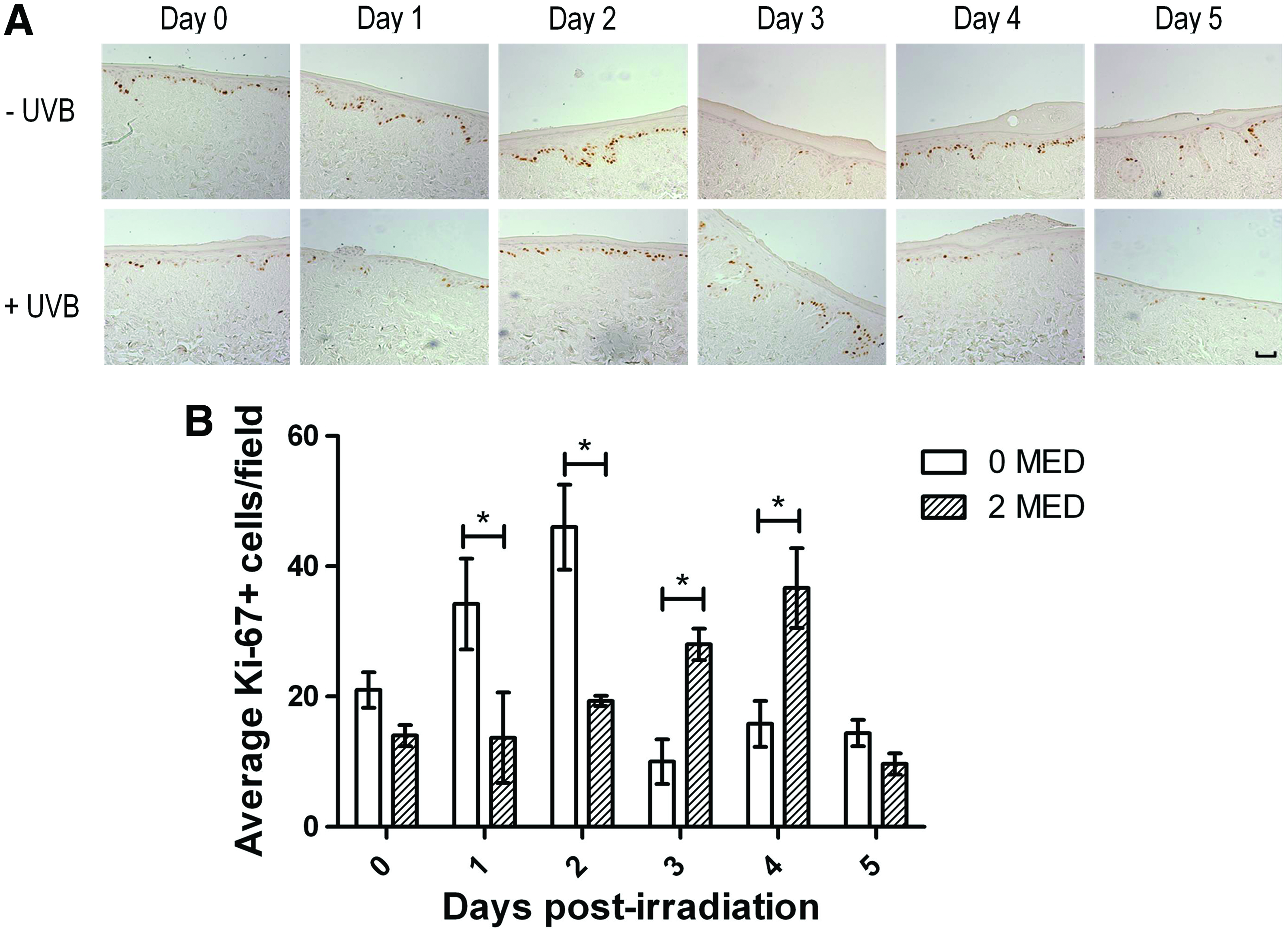
Analysis of keratinocyte proliferation in HSE-DED constructs. The proliferation marker, Ki-67, was detected using immunohistochemistry in 2 MED UVB-irradiated and non-irradiated control HSE-DED constructs
Another physiological epidermal response to UVR exposure is the thickening of the stratum corneum, which, along with the tanning, is considered a protective mechanism. 43 This increase in thickness is associated with elevated levels of keratinocyte differentiation into corneocytes. 44 The layer of differentiated keratinocytes (expressing keratins 1/10/11) in the HSE-DED epidermis was investigated subsequent to UVB exposure via immunohistochemistry to identify whether similar processes occur in these constructs (Fig. 6). These areas were then quantified using image analysis software. In the first 3 days after UVB irradiation, a decline in the average immunoreactive areas to keratins 1/10/11 was observed. This was localized to the spinous and granular layers of the epidermis. This was accompanied by a thickening of the stratum corneum layer in these constructs as observed in the tissue sections. Similar results of UVB-induced epidermal thickening have also previously been reported from in vivo studies involving irradiated test subjects.45,46
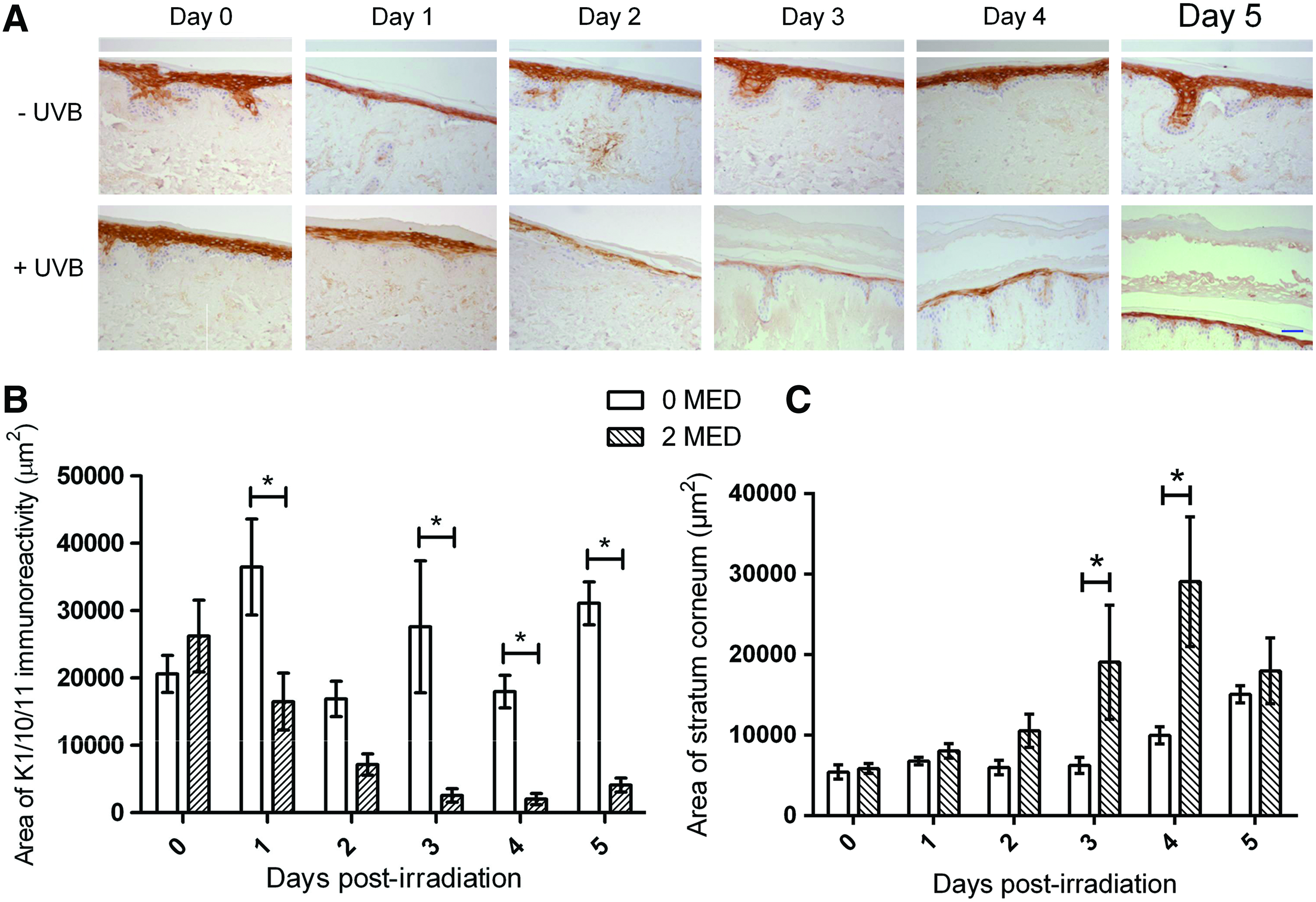
Epidermal blistering and regeneration after UVB irradiation in HSE-DED constructs. Immunohistochemical staining of keratin 1/10/11 in irradiated and mock-irradiated HSE-DED was conducted to identify changes to the thickness of the suprabasal layer containing differentiated keratinocytes
Discussion
The gold standard of in vitro photobiology models should display biological parallels with native skin, as well as classical photodamage markers on UV irradiation. Conventional cell-based techniques, such as 2D cultures, do not adequately represent the complex morphology and organizational aspects of the human epidermis. Moreover, limitations associated with animal models include ethical restrictions, high costs, and differences to human skin anatomy and physiology. Accordingly, the HSE-DED reconstituted skin model constructed here was validated as an experimental model to investigate keratinocyte responses to UVB radiation. Composed of primary human keratinocytes cultured on a DED dermal substrate, the HSE-DED constructs maintain histological and physiological aspects of native skin, many of which are critical to the UVB photoresponse. Previously, we have reported that the 3D HSE-DED organotypic model can be applied to the investigation of a diverse range of epithelial processes, including woundbhealing.21, 22 In this current study, UVB irradiation of the HSE-DED was found to induce the generation of characteristic markers of photodamage in human keratinocytes. Significantly, our data indicate that the HSE-DED keratinocytes also retained their capacity to repair during epidermal regeneration postirradiation.
Histological analysis of the HSE-DED revealed that several morphological features of the native epidermis, such as the presence of rete ridges and the epidermal structural organization, were retained. This was in accordance with previously reported studies on the histology of similar tissue-engineered skin constructed in vitro.47–49 The use of similar HSE models has also been reported in studies conducted on UVR-induced alterations to epidermal viability 50 and protein expression. 51 In these models, different dermal substrates such as collagen matrices, as well as various combinations of UVR wavelengths and doses were utilized. Importantly, to date, a fully characterized model using reconstituted human tissue is still lacking. By incorporating a single cell type into the HSE-DED model described here, we aimed at obtaining baseline data on the effects of UVB radiation in a physiologically relevant setting. This can then be used to compare, for example, the protective effects of other cutaneous cell populations, such as melanocytes and fibroblasts, on their incorporation into the HSE model.
The presence of a multilayered epithelium with a polarized differentiation gradient was further confirmed in the HSE-DED via the immunohistochemical detection of keratins 14, 1/10/11, 2e and filaggrin. Importantly, the HSE-DED enables the investigation into specific responses of basal keratinocytes to UVB radiation, in the context of a differentiated epidermis. Indeed, damage to DNA in basal keratinocytes as a result of UVB exposure is considered a major initiating factor in the development of squamous cell carcinoma and basal cell carcinomas. 3 The protective function of the stratum corneum and filaggrin expression in the HSE-DED epidermis is also significant in the modeling of UVB-induced epithelial damage.52–54 Monolayer and tissue-engineered cultures lacking filaggrin, for example, have elevated levels of DNA damage and apoptosis observed in the stratum basale. 54 In addition, the positive identification of the basement membrane component, collagen IV, at the epidermal-dermal junction is notable as a physiologically relevant photobiology model. Lacking in 2D cell cultures, such BM components have been found to profoundly influence keratinocyte responses to many external stimuli, including UVB radiation.55,56
The classical markers of acute cutaneous photodamage have been well documented, and they include the formation of DNA photolesions, apoptosis, inflammation, and the tanning response. 57 Notably, in the studies reported here, key UVB-induced keratinocyte damage markers were shown to be inducible in the HSE-DED in an in vitro setting. The formation of CPDs and other photoproducts are a fundamental mutagenic event in the transformation of basal keratinocytes. 3 In the HSE-DED, irradiation was found to result in the formation of these thymine dimers in keratinocyte nuclei, in line with investigations on UVB-exposed native skin biopsies.58,59 Interestingly, the rate of CPD removal in the HSE-DED was comparable not only to studies using other organotypic skin constructs, but also to the in vivo experiments mentioned earlier.49,60 This indicates that the cells in the HSE-DED retain the ability to initiate DNA damage response and repair pathways, which potentially enables the screening of novel mechanisms to enhance these processes.
The activation of both DNA damage-induced apoptosis and repair mechanisms are highly dependent on the function of p53.61–63 The UVB-induced accumulation of p53 in HSE-DED basal and suprabasal keratinocytes correlates with previously documented reports.63,64 Importantly, major differences in p53 activation have been identified between irradiated keratinocyte monolayers and in vivo skin. 65 This has been attributed to factors such as keratinocyte differentiation status and cellular adhesion. 66 The UVB-triggered secretion of inflammatory cytokines, including IL-1, -6, -8, -10, -12, and tumor necrosis factor-α, stimulates erythemal and immunomodulatory responses postirradiation.67–70 In line with earlier reports, our data demonstrate a significant augmentation of IL-6 and IL-8 levels in HSE-DED culture media after UVB irradiation.37,38 Potentially, the irradiated HSE-DED model could, thus, be applied to the analysis of pathways involved in inflammation as a result of UV exposure or cutaneous diseases such as psoriasis.71,72
Several previous studies on keratinocyte responses to irradiation involve the use of nonphysiological doses of UVB to induce photodamage.73,74 In part, this is because monolayer keratinocyte cultures are selective for cells with a more proliferative phenotype and an associated higher tolerance to UVB-induced damage. 75 Moreover, the use of pigmented skin equivalents has been shown to withstand higher UVB doses; this is due to the shielding effect of melanin.76–78 Similarly, other in vivo studies in pigmented skin show relatively lower levels of apoptosis induction after irradiation compared with those observed in the HSE-DED. 79 The studies reported here characterizing the HSE-DED model were conducted using physiological doses of UVB radiation. Indeed, the doses utilized have previously been reported to be experienced within the Australian population.80,81 The HSE-DED, therefore, may be applied to the testing of potential therapeutics or preventative strategies against keratinocyte damage in a biologically appropriate setting.
In summary, we demonstrate here that the HSE-DED model holds potential as a versatile and powerful photobiology tool due to the retention of several physiological cutaneous properties. Moreover, the HSE-DED represents a basic epidermal model into which additional cell types can be incorporated, resulting in the creation of an improved biologically relevant skin model. It is important to bear in mind that despite the HSE-DED model being flexible in terms of the origin and type of primary cells which may be incorporated, this may also contribute to inter-donor variability, a phenomenon that has been reported in a variety of skin studies.82–84 Indeed, studies have shown variability to exist, particularly in keratinocyte UVB responses between individuals. 85 In the future, the development of increasingly elegant synthetic dermal substitutes may provide improved dermal scaffolds that can reduce variability while preserving the physiological properties of the dermis more faithfully.19,25–27
Footnotes
Disclosure Statement
The authors declare that no competing financial interests or conflicts of interest are associated with the work presented here.