
Abstract
Lung bioengineering, a novel approach to obtain organs potentially available for transplantation, is based on decellularizing donor lungs and seeding natural scaffolds with stem cells. Various physicochemical protocols have been used to decellularize lungs, and their performance has been evaluated in terms of efficient decellularization and matrix preservation. No data are available, however, on the effect of different decellularization procedures on the local stiffness of the acellular lung. This information is important since stem cells directly sense the rigidity of the local site they are engrafting to during recellularization, and it has been shown that substrate stiffness modulates cell fate into different phenotypes. The aim of this study was to assess the effects of the decellularization procedure on the inhomogeneous local stiffness of the acellular lung on five different sites: alveolar septa, alveolar junctions, pleura, and vessels' tunica intima and tunica adventitia. Local matrix stiffness was measured by computing Young's modulus with atomic force microscopy after decellularizing the lungs of 36 healthy rats (Sprague-Dawley, male, 250–300 g) with four different protocols with/without perfusion through the lung circulatory system and using two different detergents (sodium dodecyl sulfate [SDS] and 3-[(3-cholamidopropyl) dimethylammonio]-1-propanesulfonate [CHAPS]). The local stiffness of the acellular lung matrix significantly depended on the site within the matrix (p<0.001), ranging from ∼15 kPa at the alveolar septum to ∼60 kPa at the tunica intima. Acellular lung stiffness (p=0.003) depended significantly, albeit modestly, on the decellularization process. Whereas perfusion did not induce any significant differences in stiffness, the use of CHAPS resulted in a ∼35% reduction compared with SDS, the influence of the detergent being more important in the tunica intima. In conclusion, lung matrix stiffness is considerably inhomogeneous, and conventional decellularization procedures do not result in substantially different local stiffness in the acellular lung.
Introduction
S
Recently, several authors have used a variety of lung decellularization methods and assessed their effectiveness for eliminating donor cells while minimizing the alterations caused in the extracellular matrix (ECM).9,10 Striking this balance is not easy since a decellularization procedure should be aggressive enough to eliminate all the donor cell materials and at the same time allowing for the preservation of the composition and structural integrity of the ECM. Different decellularization methods have been used, based on the combination of physical steps for breaking down cell integrity (e.g., freezing/thawing and osmotic shock) and molecular agents for washing out the cell debris and obtaining an acellular lung (e.g., different types of detergents).11–17 The suitability of these decellularization procedures has been evaluated in terms of the composition and structural integrity of the ECM and mechanical properties of the whole acellular organ. 18
However, there are no data available on how different decellularization procedures may affect the stiffness of the lung ECM at the local level. This information is distinct from that provided by the pressure–volume relationship in the whole acellular lung. 19 Indeed, the bulk mechanical properties of the 3D mesh do not directly reflect the local stiffness of the fibers that constitute the matrix, 20 particularly in a heterogeneous scaffold comprising a variety of substructures, such as bronchi, alveoli, and blood vessels. It should be noted that local stiffness in acellular lungs may be relevant in lung bioengineering since each individual cell seeded during the recellularization process will sense the mechanical properties of the specific substrate where it is placed. And it has been shown that cell engraftment, proliferation, and, particularly, differentiation into phenotypes are modulated, among other factors, by substrate stiffness,21–27 particularly concerning epithelial28–30 and endothelial cells and vasculogenesis.31–33 The aim of this study, therefore, was to assess the effects of the decellularization procedure on the inhomogeneous local stiffness of the acellular lung. To this end, we used atomic force microscopy (AFM) to measure Young's modulus of acellular lungs on different sites in the alveoli, pleura, and vessels, using different decellularization protocols.
Methods
Animals and lung extraction
The study was carried out on lungs excised from 36 Sprague-Dawley male healthy rats (250–300 g), following experimental procedures approved by the Ethical Committee for Animal Research of the University of Barcelona. The rats were anesthetized with intraperitoneal urethane (1 g/kg), heparinized (250 U/kg), and sacrificed by exsanguination through the abdominal aorta. In half of the rats (N=18), the pulmonary artery was cannulated and 50 mL of phosphate-buffered saline (PBS) containing streptomycin (90 μg/mL), penicillin (50 U/mL), and amphotericin B (25 μg/mL) were perfused with a pressure of 20 mmHg. Finally, the lungs and heart were excised and, after removing external connective tissue, they were frozen by placing them in a −80°C freezer, and they were stored at this temperature until the decellularization process.
Lung decellularization
The first step in lung decellularization was a fourfold repetition cycle that involved thawing the lungs in a water bath at 37°C and freezing them again at −80°C (Fig. 1). After this first step, two different decellularization procedures were followed, depending on whether the lungs were previously perfused or not.
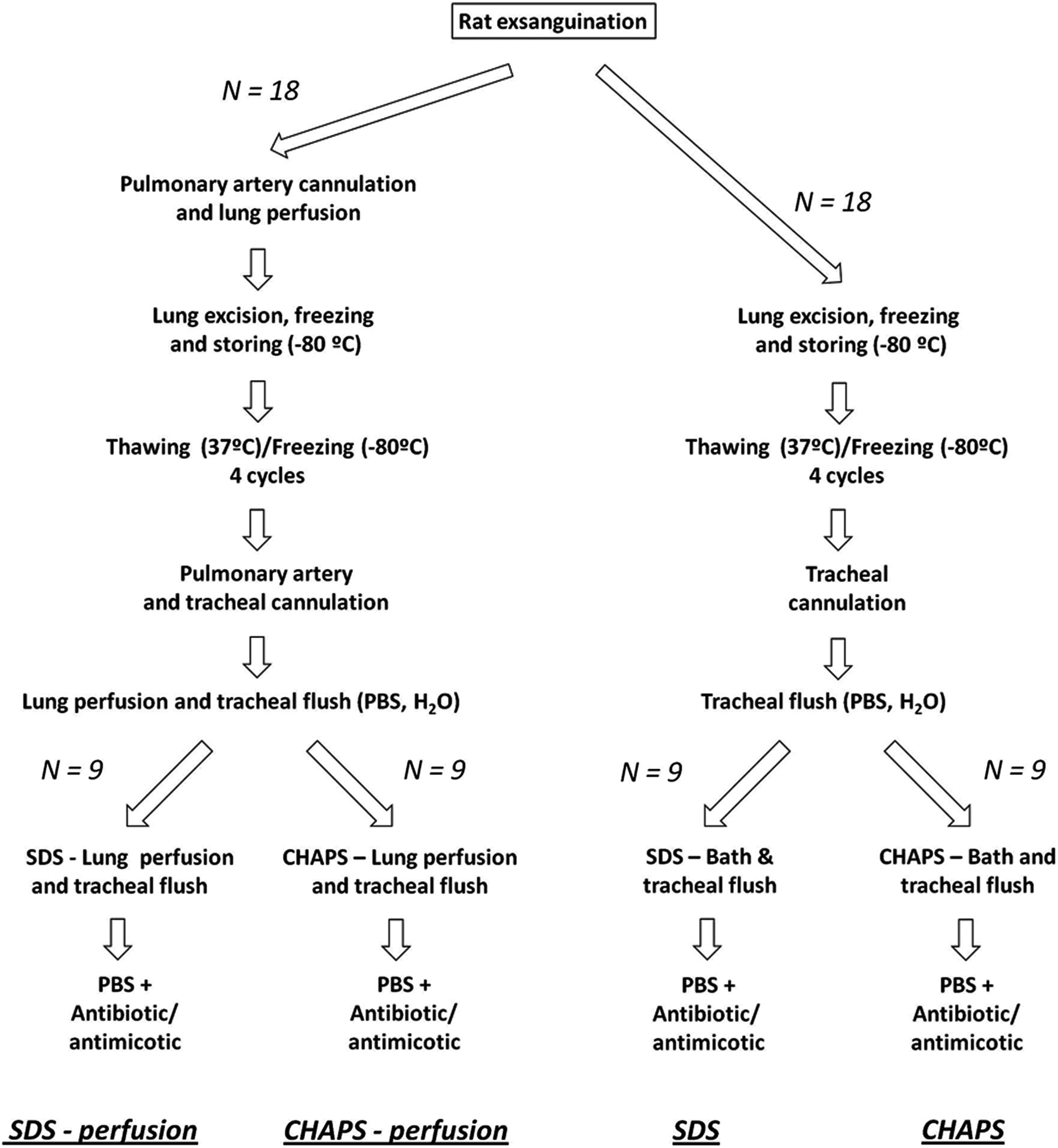
Lung decellularization procedures. Four decellularization procedures were performed by using two different detergent solutions (SDS and CHAPS) with and without perfusing these solutions through lung vessels, each in nine rat lungs. PBS, phosphate-buffered saline medium; SDS, solution containing sodium dodecyl sulfate; CHAPS, solution containing 3-[(3-cholamidopropyl) dimethylammonio]-1-propanesulfonate. See “Methods” for details.
The lungs that were excised without previous perfusion (Fig. 1, right side) were first rinsed and flushed through a tracheal cannula with PBS and deionized water several times until the liquid extracted from the lungs had a transparent appearance. Subsequently, the lungs were flushed with a decellularization solution—described below—until the liquid emanating from the lungs became transparent. The lungs were then kept under agitation inside this solution for 3 days and flushed with 25 mL of solution through the trachea twice a day. Finally, the lungs were flushed and maintained under agitation for another 2 days with a PBS solution containing the antibiotics and antimycotic at the concentration previously described (Fig. 1).
The lungs that were perfused before excision (Fig. 1, left side) followed a similar protocol to the one described above but with the addition of perfusion through the vascular compartment simultaneous with the flushing through the trachea. First, the lungs were rinsed and flushed with PBS through the tracheal cannula until the liquid was coming out transparent. The pulmonary artery was then cannulated and perfused with PBS for 30 min at a pressure of 20 mmHg, with deionized water for 15 min and with a 500 mL decellularization solution, as described below, for 2–3 h. During this process, a solution identical to the perfused solution was flushed through the trachea. Finally, the lungs were perfused with the previously described PBS solution for 30 min.
Two previously described decellularization solutions were used in both the perfused and nonperfused lungs.11,14 We compared the use of 1% sodium dodecyl sulfate (SDS) 11 and 8 mM of 3-[(3-cholamidopropyl) dimethylammonio]-1-propanesulfonate (CHAPS), 1 M NaCl, and 25 mM ethylenediaminetetraaceticacid in PBS. 14 Accordingly, four different decellularization processes were carried out (N=9 each): perfusion with SDS or CHAPS solutions (P-SDS and P-CHAPS, respectively) and nonperfusion with SDS or CHAPS solutions (NP-SDS and NP-CHAPS, respectively).
Preparation of lung matrix slices
Slices of the decellularized lungs were prepared for imaging and for assessing the local stiffness of the matrix using AFM. The acellular lungs were embedded in a solution containing Optimal Cutting Temperature Compound (OCT; Sakura) and PBS in a 4:1 ratio. This solution was also infused (3 mL) through the trachea to fill the lungs until a physiological end-inspiratory inflation level and the lobes were excised and snap-frozen in an isopentane bath (precooled in liquid nitrogen) for cryosectioning. The slices for AFM measurements were cut at a nominal thickness of 7 μm and adhered on top of positively charged glass slides (Superfrost; Thermo Fischer Scientific). The lung matrix slices to be used for immunostaining and imaging with conventional confocal microscopy or with multiphoton microscopy were cut at the thickness of 40 μm and placed on top of cover slips pretreated with silane. This pretreatment involved incubating the cover slips for 5 min with a silane solution prepared in acetone (1:50), then rinsing them with acetone and, finally, with deionized water. The lung slices were stained with Hoechst 33342 (Molecular Probes) to allow us to detect nuclei within the wavelengths used in our two-photon microscopy setting, specifically in the same channel as elastin. 34
Additional fresh rat lungs were prepared to provide a control in the immunostaining and multiphoton microscopy imaging. Briefly, the lungs were excised, embedded in OCT (as previously described), and snap-frozen in an isopentane bath (precooled in liquid nitrogen). These fresh lungs were not perfused to keep them as intact as possible. Fresh rat lungs were also sliced at a thickness of 40 μm and placed on top of silane-treated cover slips. The slices were stored at −20°C until they were stained and imaged.
Imaging decelullarized lung slices
To perform conventional fluorescent immunostaining, decellularized 40-μm-thick lung slices were allowed to thaw and the OCT was removed by several PBS rinses. The samples were then blocked with a solution containing TBS, 0.5% triton, 6% fetal bovine serum (FBS), and 1% bovine serum albumin (BSA) for 45 min. Primary antibody against laminin (Sigma; 1:20) was incubated in a solution of TBS, 0.1% triton, 6% FBS serum, and 0.1% BSA overnight at 4°C. Slices were allowed to warm up at room temperature for 15 min and rinsed three times with the same type of solution. Secondary donkey anti-rabbit Cy3 (Jackson) antibody was incubated at a 1:200 dilution in 1× TBS+0.1% triton+6% FBS serum+0.1% BSA for 2 h. Three 15-min rinses with PBS were applied to eliminate the unbound secondary antibodies. Fresh-frozen lung slices (40 μm thick) were treated with the same protocol, but a fixation step with 4% paraformaldehyde for 10 min, followed by several PBS rinses, was introduced before the blocking. Images were obtained by a Leica SP-5 fluorescence microscope. Glycosaminoglycans (GAGs) were conventionally stained with Alcian blue and observed by optical microscopy (TE2000; Nikon).
Collagen and elastin were imaged using a nonlinear technique of second-harmonic generation (SHG) and two-photon excited fluorescence (TPEF) in a simultaneous mode.35,36 Collagen has a highly crystalline triple-helix structure that is not centrosymmetric. This makes it very efficient in generating the second harmonic of incident light. Elastin, which is normally found together with collagen, is a significant source of ECM autofluorescence and can be imaged with TPEF. The experimental arrangement was based on a Leica inverted confocal laser scanning microscope SP-5 with META scanning module equipped with a mode-locked Ti:sapphire femtosecond laser. Laser emission was 810 nm, and two channels were used to simultaneously detect the SHG and TPEF signals. The detection spectral range was a narrow band (387–409 nm) for the SHG signal of the collagen and a broadband (450–600 nm) for the TPEF signal that belonged to the elastic fibers. The images of the 40 μm slices were collected using a Leica 25×0.95-NA water objective. The step-size applied to the z-stacks acquired was 2.1 μm. For the multiphoton techniques, the average laser powers were 4 mW in the TPEF detection channel and 7 mW in the SHG detection channel, respectively.
Samples for scanning electron microscopy (40 μm thick) were fixed using 2% glutaraldehyde and 2.5% paraformaldehyde in 0.1 M cacodylate buffer (EMD Biosciences) for 2 h at room temperature, then rinsed in cacodylate buffer, and dehydrated through an ethanol gradient. Samples were further dehydrated in hexamethyldisilizane for 10 min and dried overnight, then sputter-coated with gold, and analyzed using a JEOL JXA-8600.
Measurement of local stiffness in decellularirzed lung slices
Before the AFM measurements, the acellular lung slices (7 μm thick) were allowed to thaw and the OCT was removed by several PBS rinses. A rubber chamber was placed on top of the slide and filled with PBS, which was replaced regularly during the measurements. A custom-built AFM 37 attached to an inverted optical microscope (TE2000; Nikon) was used to measure the local Young's modulus (E) of the decellularized lung matrix. Briefly, a 3D piezoelectric translator was used to position the tip on the region of interest and drive and record the vertical displacement of the cantilever (z). The cantilever deflection (d) was measured using the optical lever method. The matrix was probed by Si3N4 V-shape cantilevers with nominal spring constant of 0.1 and 0.03 N/m and with a pyramidal tip (MLCT; Bruker). The slope of a deflection–displacement (d–z) curve obtained on a bare region of the cover slip was used to calibrate the relation between the photodiode signal and the cantilever deflection. A fully linear calibration curve with a sharp contact point was taken as indicative of a clean and undamaged tip.
For each lung matrix slice, local stiffness was measured on five different sites: alveolar septum, septum junction, visceral pleura, and vessels' tunica adventitia and tunica intima (Fig. 2). For each site, stiffness was measured at five 3 μm-separated points, and each measurement consisted of five d–z curves (triangular ramp, oscillation frequency of 1 Hz and maximum indentation of ∼1 μm). The d–z curves were converted to force–indentation (F–δ) curves by assuming linear elasticity for the cantilever [F=k·(d−d0)] and defining indentation as δ=(z−z0)−(d−d0). The contact point (z0, d0) and the Young's modulus (E) were calculated for each force–indendation curve, using custom built software (MATLAB; The MathWorks), by the least-squares fitting of the Hertz pyramidal model to the F–δ curves. 38 The fitting was performed with a maximum indentation of 0.5 μm to avoid the effect of the underlying glass slide. Moreover, the effect of the Young's modulus on indentation was assessed by progressively increasing the indentation range of the fitting. For a given measurement point, E was computed as the average of the Young's modulus corresponding to each of the five force–indentation curves. E for each site in a given sample was computed as the average of the E values obtained for the five different points measured on that site.

Top view of decellularized lung slices during the measurement of local stiffness with AFM measurements. In the top image, the AFM tip was on the pleura (P). A representative alveolar septum (S) and junction (J) are also marked. The bottom image shows a vessel with the AFM tip on top of the tunica intima (TI). The tunica adventitia (TA) can also be clearly distinguished. Scale bars correspond to 50 μm. AFM, atomic force microscopy.
Statistical analysis
For each acellular lung site (alveolar septum, septum junction, visceral pleura, and vessels' tunica adventitia and tunica intima) and decellularization method (SDS with/without perfusion and CHAPS with/without perfusion), stiffness E was computed from averaging the results obtained from the nine independent experiments carried out (N=9): 36 rat lungs, each independently subjected to one of the four decellularization processes. Stiffness data are presented as mean±standard error (SE). A two-way analysis of variance was used to assess differences in E attributable to two factors: acellular lung site and decellularization procedure. Post hoc analyses were carried out with the Student–Newman–Keuls method. Statistical significance was considered at p<0.05.
Results
Figure 3 shows images of acellular and normal lung slices stained for the presence of elastin (and cell nuclei), collagen, laminin, and GAGs. These examples qualitatively illustrate that, as expected, these representative components of the ECM remained after the application of the different decellularization procedures. All the acellular lungs exhibited very few nuclei, compared with normal lung. Figure 4 illustrates the different relative abundance of collagen and elastin in the substructures of the lung matrix. Figure 5 shows examples of lung vessels when using SDS and CHAPS solutions for decellularization.
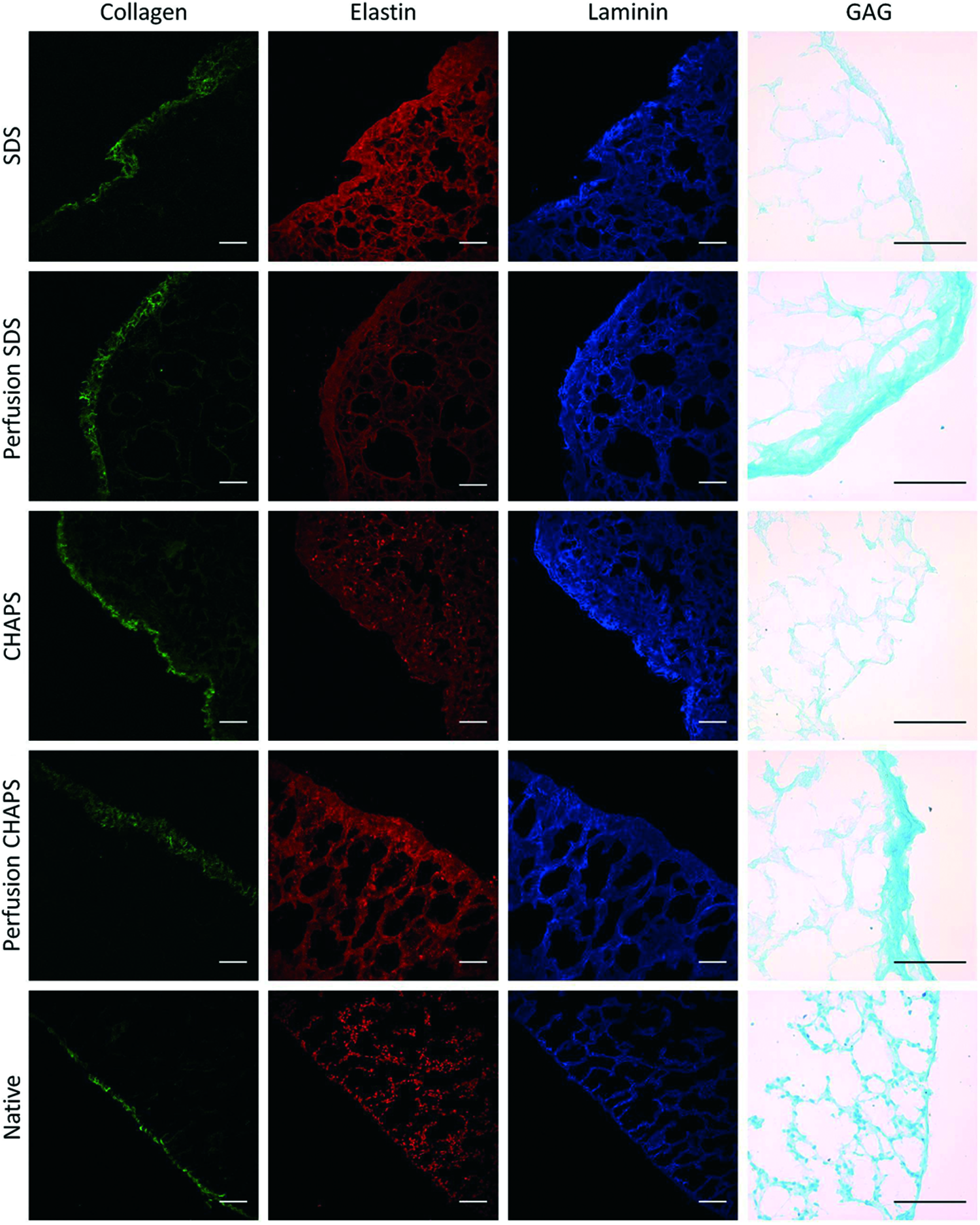
Representative images of decellularized and fresh lung slices for elastin (including nuclei stained with Hoechst), collagen, laminin, and GAGs. Images of elastine, collagen, and nuclei were obtained by two-photon microscopy, laminin by conventional immunofluorescence, and GAGs by Alcian blue staining. Scale bars correspond to 75 μm. GAGs, glycosaminoglycans. Color images available online at

Examples of collagen and elastin content at the substructures of the lung matrix. Two-photon and second harmonic generation images
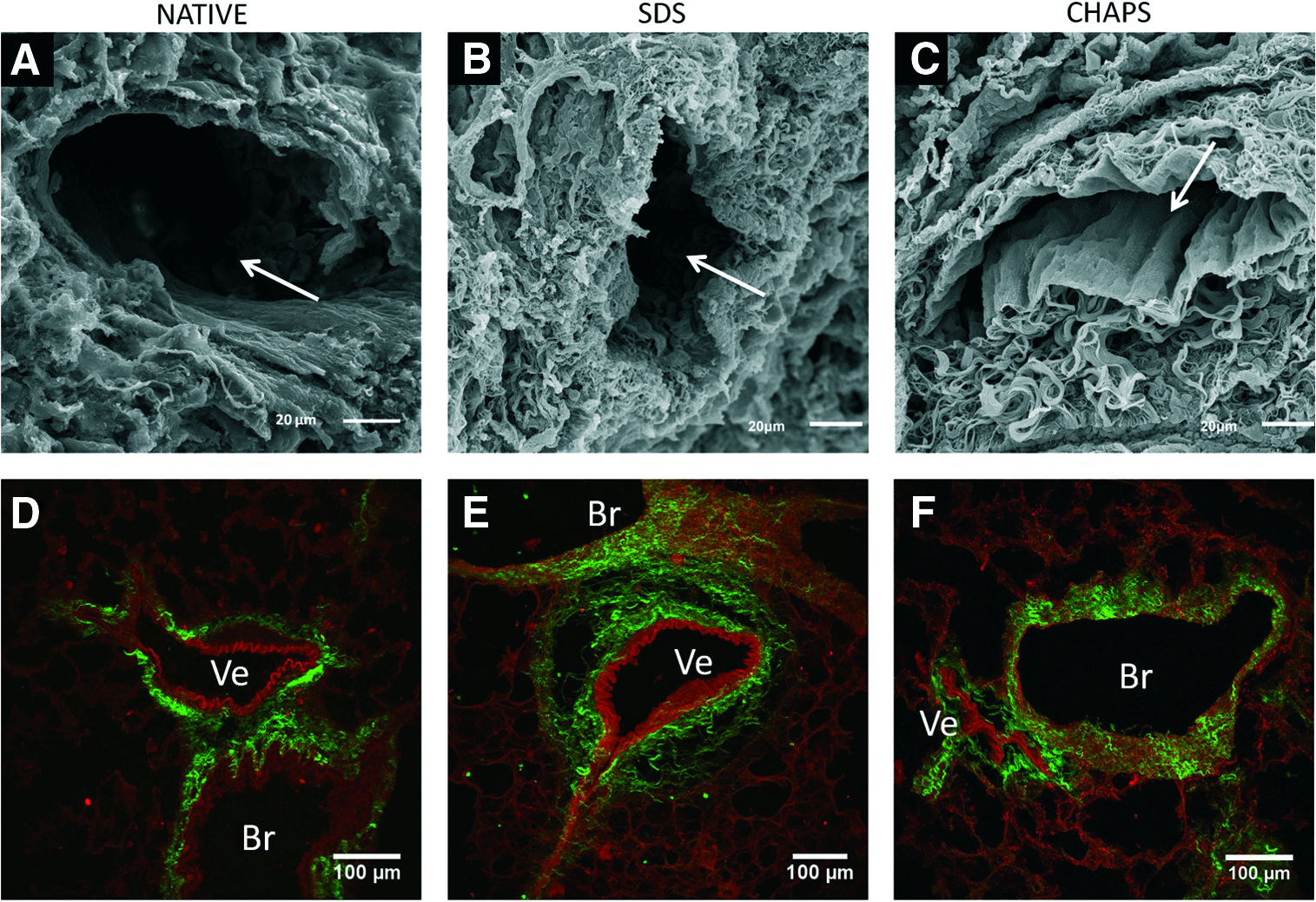
Examples of lung vessels areas in a native lung and in lungs decellularized with two detergents (SDS and CHAPS). Lung parenchyma vessels when decellularized with CHAPS
The dependence of the Young's modulus on indentation depth is shown in Figure 6. Data are the average of the fitting performed in the alveolar septa of each rat taking a force curve recorded with indentation of >0.8 μm. After marked changes at shallow indentations (<300 nm), the Young's modulus reached a plateau extended beyond the maximum indentation of 500 nm used in the fitting algorithm.
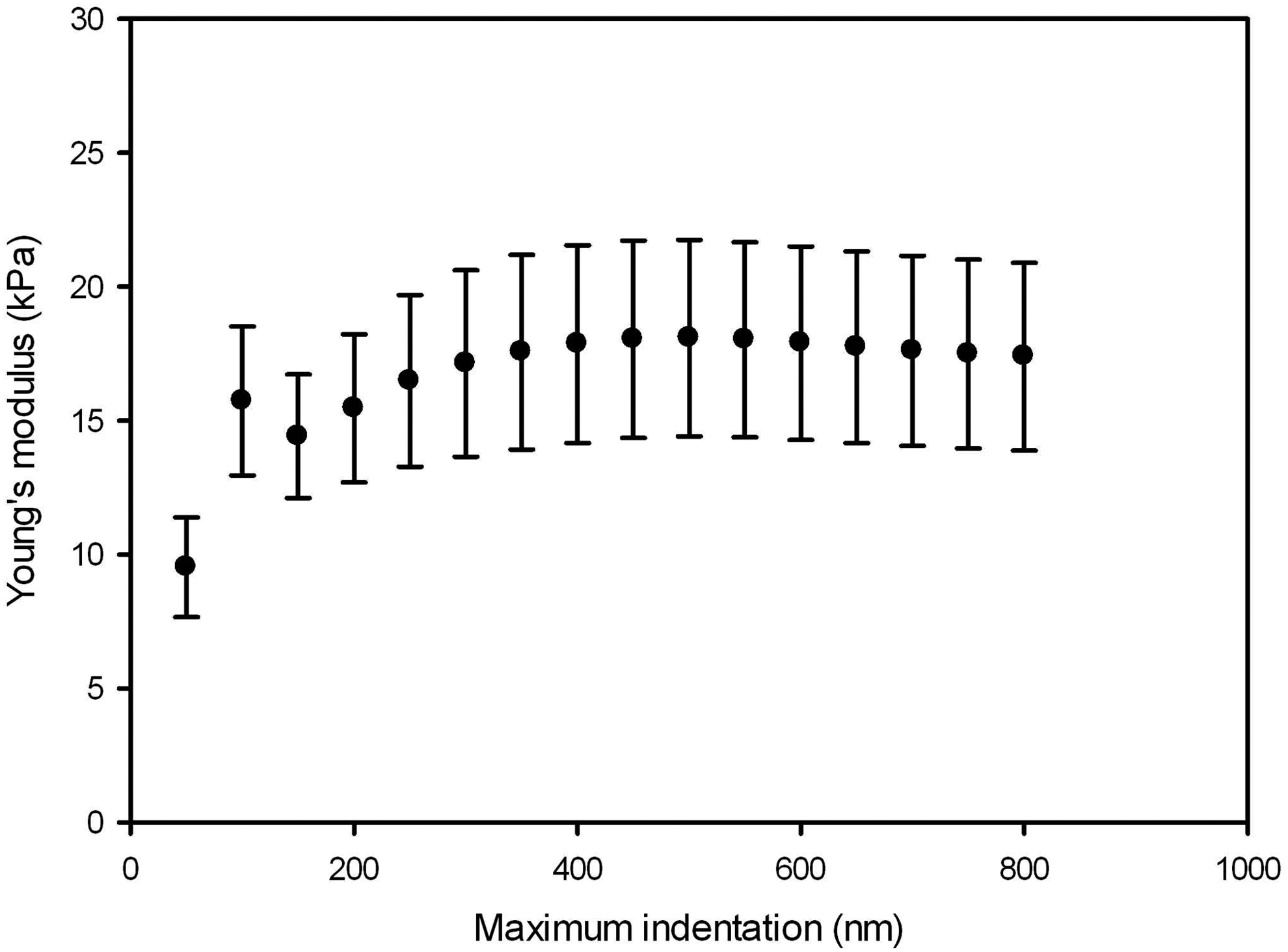
Dependence of the Young's modulus on maximum indentation. Data are mean±SE of fitting performed in each rat (n=9) from a force curve recorded in alveolar septa. SE, standard error.
Figure 7 shows the stiffness (mean±SE) of the different lung regions after the four decellularization processes. The coefficient of variation (CoV=standard deviation/mean) ranged from 21% in alveolar septum decellularized using SDS without perfusion to 74% in tunica adventitia subjected to SDS with perfusion. This relatively low variability compared with those found in bulk parenchymal strips15,39,40 could be partly attributable to our precise selection of the measured region (Fig. 2) and also to the fact that we characterized the stiffness of each region in a given rat as the average of measurements taken at five points on that region.

Local stiffness of decellularized rat lungs. Young's modulus (E) was measured by AFM at different sites of the acellular lung for the four decellularization procedures used combining two detergents (SDS and CHAPS) without or with vascular perfusion. Values correspond to the mean and standard error computed from nine rats. Each individual data for a given rat and decellularized lung site was computed as the average from five measurements, as detailed in “Methods”. The local stiffness of the acellular lung matrix significantly depended on both the site within the matrix and the decellularization process. Specifically, differences between SDS and CHAPS were statistically significant only for the tunica intima (*p<0.05). Data are mean±SE (n=9).
The local stiffness of the acellular lung matrix significantly depended both on the site within the matrix (p<0.001) and on the decellularization process used (p=0.003), with significant interaction between the two factors (p<0.001).
Pooling the data of the four decellularization methods, the lowest values of Young's modulus were found in the alveolar septum and junction, showing similar values (16.8±2.4 and 15.6±2.5 kPa, respectively). E for the pleura and the tunica adventitia were similar (34.6±2.4 and 30.9±2.5 kPa, respectively) and significantly greater than septa E values. The Young's modulus of the tunica intima was 59.6±2.5 kPa, significantly higher than on the other sites of the ECM (Fig. 7).
Applying perfusion during lung decellularization did not result in significant differences in the Young's modulus (Fig. 7). In contrast, the type of detergent used did result in significant changes. The Young's modulus for SDS and CHAPS, computed as the average of the stiffness measured for all sites, was 39.8±3.9 and 22.9±3.5 kPa, respectively, when no perfusion was applied, and 37.8±3.6 and 25.5±3.5 kPa, respectively, when decellularization was performed with perfusion. More specifically, the differences between SDS and CHAPS were statistically significant only for the tunica intima, the Young's modulus being almost half the value for CHAPS that it was for SDS (Fig. 7).
Discussion
This study was aimed to assess whether the application of different detergent-based decellularization procedures affects local stiffness on different sites on the acellular lung matrix. Specifically, the effects of perfusing or not perfusing decellularization agents through the lung circulatory system and the use of SDS and CHAPS as detergents were investigated. One initial result—which was novel, as it was not previously investigated—was that there was a considerable inhomogeneity in the local stiffness of the acellular lung, despite the decellularization method used, and, interestingly, this inhomogeneous pattern was the same regardless of the decellularization process. Specifically, the vessel tunica intima matrix was four times stiffer than the alveolar septum. The main result indicated that, on the whole, acellular lung stiffness depends significantly, albeit modestly, on the decellularization process. Whereas perfusion was a factor that did not induce significant differences, using CHAPS as detergent resulted in a whole reduction stiffness by ∼35% compared with SDS (average across all measured sites), the influence of the detergent being more pronounced in the tunica intima.
The measurement of local stiffness was carried out with AFM since this technique allows for the measurement of the Young's modulus of the sample at the cell scale, and more specifically, the micrometric dimensions of focal adhesions. 41 To this end, instead of using an AFM spherical tip with a relatively large contact area that would sense different acellular lung structures,15,39 we used an AFM sensor with a pyramidal tip. 42 To avoid the effect of the rigid underlying substrate, we fitted the pyramidal Hertz model in an indentation range with maximum indentation (500 nm) higher than ∼300 nm and lower than 10% of the thickness of the sample (7 μm).37,42 Moreover, we computed the dependence of E on the indentation range (Fig. 6). The Young's modulus exhibits substantial variability at low indentation, reaching a reasonable plateau for indentations deeper than ∼300 nm. Therefore, the maximum indentation of 500 nm used in our fitting algorithm avoids the uncertainties in the contact point and the effects of the rigid substrate. Furthermore, the observed plateau reveals a nearly linear behavior for the low local strains applied by the AFM tip. Taking into account the measurement indentation (0.5 μm), the area of contact between the tip and the sample was ∼0.05 μm2 (lateral length 0.2 μm). Therefore, the tip–matrix contact lateral dimension was more than one order of magnitude lower than the thickness of the thinner sites measured (septum wall and tunica intima) (Fig. 2). Interestingly, this lateral resolution permitted the detailed measurement of local heterogeneity in the acellular lung, including several independent measurements within the same site. Moreover, the indentation was in the range of local deformations exerted on the microenvironment by active contraction of the cell cytoskeleton. 43 Thus, the local stiffness of the different sites in the acellular lung was measured on a scale that would be sensed by the cells seeded during recellularization.
The considerable differences observed in local stiffness between acellular lung sites can be partly attributed to the ECM components in each region. Indeed, whereas collagen was mostly present in the tunica adventitia and the pleura, almost no collagen was found in the tunica intima of the vessels, as it is composed of a dense laminar layer of elastin (Fig. 4). Although qualitative, these images may help to interpret the Young's modulus data we obtained (Fig. 7). The considerable high value of stiffness of the tunica intima (∼60 kPa; four times that of alveolar septa) could be attributed to the high concentration of elastin. Moreover, the absence of elastin and presence of collagen in the pleura and tunica adventitia could explain why these sites exhibited a stiffness (∼32 kPa) that fell between those of the alveolar septa and the tunica intima. This interpretation of the local stiffness of elastin- versus collagen-enriched sites within the acellular lung is consistent with results reported in the porcine arterial wall. 44 These authors showed that the Young's modulus of the elastin-rich region of the arteria ranged from 50 to 70 kPa (remarkably close to the 60 kPa reported here for the lung vessel tunica intima) and that the stiffness of collagen-rich regions was significantly lower. 44
From a lung bioengineering viewpoint, the local stiffness inhomogeneity observed is of particular interest when considering the sites where stem cells seeded during recellularization will engraft. The two collagen-richest sites—pleura and tunica adventitia—share a similar local stiffness (Fig. 7) and population by stromal cells (submesothelial cells and pericytes, respectively).45,46 Moreover, the alveolar wall and the tunica intima—with a fourfold difference in stiffness—would provide different mechanical cues to seeded cells. Taking into account the modulatory effect of substrate stiffness on stem cell fate,21–27 it could be expected that the differentiation of stem cells homing into the alveoli and the lung vessel walls into phenotypes typical of these areas (i.e., epithelial vs. endothelial/smooth muscle) could partly be driven by the local stiffness of the acellular lung.11,47 However, there are no quantitative data available on the specific case of differentiation toward the different lung cell phenotypes of interest in lung bioengineering. When SDS detergent was used in the decellularization process, the tunica intima, the substrate for endothelial cells, was approximately four times stiffer than the alveolar septa, the substrate for epithelial cells (Fig. 7). According to indirect data available,21–27 it could be possible that such a difference in matrix stiffness plays a role in the potential differentiation of stem cells toward epithelial or endothelial phenotypes. In this respect, it is interesting to note that when CHAPS was used as a decellularizing detergent, the stiffness of the tunica intima was almost half the value than when SDS was used (Fig. 7), suggesting that specific details in the decellularization process of the lung could have an impact when the acellular lung matrix is recellularized. However, these specific questions require further investigation.
The Young's modulus of alveolar and pleural regions is in agreement with the elastic shear approximately threefold larger than the storage modulus (G′) reported by us in a recent rheological study applying low-amplitude oscillation, where the oscillatory response of the acellular lung revealed a solid-like behavior. 37 This E/G′ ratio is consistent with the solid-like behavior exhibited by the alveolar and pleural matrix. Given that in a solid-like material E≈G′·2·(1+ν), being ν the Poison ratio, and that ν=0.5 for an incompressible material, E/G′ is expected to be≈3. 38 Consistently with this ratio, the values of E for the alveolar and pleural regions in this work were approximately threefold larger than G′ reported in our previous study. 37 On the other hand, Booth et al. 15 measured two samples of native and decellularized human lung parenchymal strips (1 mm thick) with AFM. The decellularized strips exhibited a mean E of 1.6 kPa similar to that of the native samples. Comparable stiffness was reported by Liu et al. 39 and Brown et al. 40 in native parenchymal strips of mouse lungs measured with AFM. Taken together, these data indicate that lung ECM is one order of magnitude stiffer than parenchymal strips. Tissue strips exhibit a complex porous 3D structure embedding alveoli, small airways, and blood vessels. AFM measurements performed in thick samples reflect the intrinsic mechanical properties of the different parenchymal components, as well as additional factors including alveolar wall thickness, 3D network geometry, and gas trapped within the alveoli. Moreover, strip stiffness measurements could be affected by residues of the low-melting agarose used to prepare the tissue strips. The main objective of our work was to assess the differential effect of four methods of lung decellularization on the intrinsic mechanical properties of different regions of the lung. To specifically measure the intrinsic mechanical properties of the different cell niches of the lung, excluding network effects, we probed thin slices (7 μm) of decellularized lungs. The AFM tip applied local deformations in the micrometer scale. As reflects the plateau in Figure 6, the deformation field spreads less than the thickness of the sample. Under these conditions, we measured the intrinsic mechanical properties of the ECM, which depend on the matrix composition but not on the thickness of the sample. In contrast, the mechanical behavior of lung parenchymal strips in tensile assays should depend on alveolar wall thickness and composition. Tanaka et al. 48 compared changes in mechanical properties with changes in tissue composition of native parenchymal strips of baby, young, and adult rats. Interestingly, they found an increase in strip stiffness during maturation attributable to alveolar wall thickening rather than changes in the amount of collagen and elastin fibers in the tissue.
The extensive literature on methods of organ decellularizacion describes different approaches to ideally eliminating the donor cell content, thereby leaving an intact extracellular organ scaffold.9,10 It has been reported that each physical, chemical, or enzymatic step has both advantages and drawbacks, and the limited experience available suggests that the optimal decellularization process would be organ-dependent. 8 Lung decellularization is a relatively new field of research, compared with that of other organs, and the experience accumulated to date is limited. However, the few groups working on lung decellularizacion have used several detergents (mainly, but not exclusively SDS and CHAPS), and in some cases, the decellularizing agents and media were perfused through the lung vasculature, intratracheally infused or with a combination of both methods.11–17,47 Although there is no systematic study assessing an extensive sample of lung decellularization methodologies in different species (mouse, rat, porcine, and human are the most common ones), it seems that, in terms of matrix preservation, several approaches provide quite similar results, with the time required to complete the process a significant parameter distinguishing the methods. 49
In this study, four common lung decellularization approaches were assessed. As expected from previous reports—and illustrated by Figure 3—elastin, collagen, laminin, and GAGs, which can be considered a representative sample of ECM components, were similarly preserved by the four decellularization procedures. Figure 3 also illustrates that GAGs tended to be partially increased in the pleural zone when the decellularization process was carried out with perfusion, for both SDS and CHAPS. This finding did not translate into changes in local stiffness, although it has been previously reported that GAGs substantially contribute to the elastic properties of the lung tissue. 50 It was also observed that when using CHAPS, vessel walls were more prone to partial collapse than when SDS was used (Fig. 5). This tendency to collapse could be partly caused by the reduced local stiffness observed in both the tunica intima and the adventitia when using CHAPS, compared with SDS (Fig. 7). However, in this study, imaging was carried out in a qualitative way to illustrate that, as expected from other published reports using similar lung decellularization procedures, the different main components of the ECM were still present in the acellular lungs. A precise determination of the effects of decellularization processes on the content of each cellular matrix component, which was not the aim of this study, would require quantitative measurements by conventional Western blots, image data processing, or proteomic analysis.15,51–53
It should be mentioned, however, that no overall conclusions were reached as regards the preferability of lung decellularization with or without perfusion, or of SDS versus CHAPS. Indeed, other factors in the process may play a significant role in the effectiveness of lung decellularization, and specifically in preserving local composition and stiffness. Factors such as the concentration of detergent in the media, the extension of infusion/perfusion (in terms of the number/duration of maneuvers or perfused volume), or the duration of the decellularization process may be more decisive than the specific type of detergent used. This may explain why, for instance, one report suggests that CHAPS could be more suitable than SDS for lung decellularization, both as regards ECM preservation and whole organ pressure/volume relationship. 18 Whereas these authors consistently reported that their specific CHAPS treatment preserved matrix composition and bulk mechanics much better than their SDS treatment, the data presented here suggest that the specific protocols used produced similar results in terms of both the preservation of the main matrix components and of local stiffness (Figs. 3 and 7). Therefore, given the limited experience available in the literature, and the considerable number of variables needed to fully define a lung decellularization process, each specific procedure should be assessed in detail.
In conclusion, lung matrix stiffness is considerably inhomogeneous and conventional decellularization procedures do not result in substantially different local stiffness in the acellular lung, with vessel walls being the site potentially most sensitive to the decellularization method. For lung bioengineering, the measurement of local stiffness on relevant sites of the acellular lung scaffold, as described in this work, is an interesting tool for additionally characterizing the micromechanics of acellular lungs after a given decellularization process.
Footnotes
Acknowledgments
The authors wish to thank Rocío Nieto and Miguel A. Rodríguez for their excellent technical assistance. This work was supported in part by the Spanish Ministry of Economy and Competitiveness (SAF2011-22576, FIS-PI11/00089).
Disclosure Statement
No competing financial interests exist.