
Abstract
The mechanical and biochemical microenvironment influences the morphological characteristics of microvascular networks (MVNs) formed by endothelial cells (ECs) undergoing the process of vasculogenesis. The objective of this study was to quantify the role of individual factors in determining key network parameters in an effort to construct a set of design principles for engineering vascular networks with prescribed morphologies. To achieve this goal, we developed a multiculture microfluidic platform enabling precise control over paracrine signaling, cell-seeding densities, and hydrogel mechanical properties. Human umbilical vein endothelial cells (HUVECs) were seeded in fibrin gels and cultured alongside human lung fibroblasts (HLFs). The engineered vessels formed in our device contained patent, perfusable lumens. Communication between the two cell types was found to be critical in avoiding network regression and maintaining stable morphology beyond 4 days. The number of branches, average branch length, percent vascularized area, and average vessel diameter were found to depend uniquely on several input parameters. Importantly, multiple inputs were found to control any given output network parameter. For example, the vessel diameter can be decreased either by applying angiogenic growth factors—vascular endothelial growth factor (VEGF) and sphingosine-1-phsophate (S1P)—or by increasing the fibrinogen concentration in the hydrogel. These findings introduce control into the design of MVNs with specified morphological properties for tissue-specific engineering applications.
Introduction
E
Moreover, the dominating influence determining network morphology is generally assumed to consist of fluid flow dynamics and related shear forces acting on the endothelium. However, it has long been recognized that mechanical and biochemical properties of the microenvironment can also play important roles in determining MVN morphology. The goal of our research was, therefore, to probe the biochemical and mechanical environment of ECs undergoing the process of vasculogenesis in a three-dimensional hydrogel to determine the influence of these factors on the morphological characteristics of the resulting engineered MVNs. By uncovering these design principles, we are able to take advantage of the natural control mechanisms employed in vivo to guide MVN formation toward a desired morphology.
To achieve this goal, we used a multiculture perfusable microfluidic platform enabling real-time observation and independent control over paracrine signaling, cell-seeding densities, and hydrogel mechanical properties (Fig. 1A, B). Human umbilical vein endothelial cells (HUVECs) were seeded in fibrin gels and cultured alongside—but not in contact with—human lung fibroblasts (HLFs). HUVECs spontaneously formed networks within 24 h and the engineered vessels contained patent, perfusable lumens as demonstrated by the passage of fluorescent microspheres after 4 days (Supplementary Videos SV2 and SV3; Supplementary Data are available online at
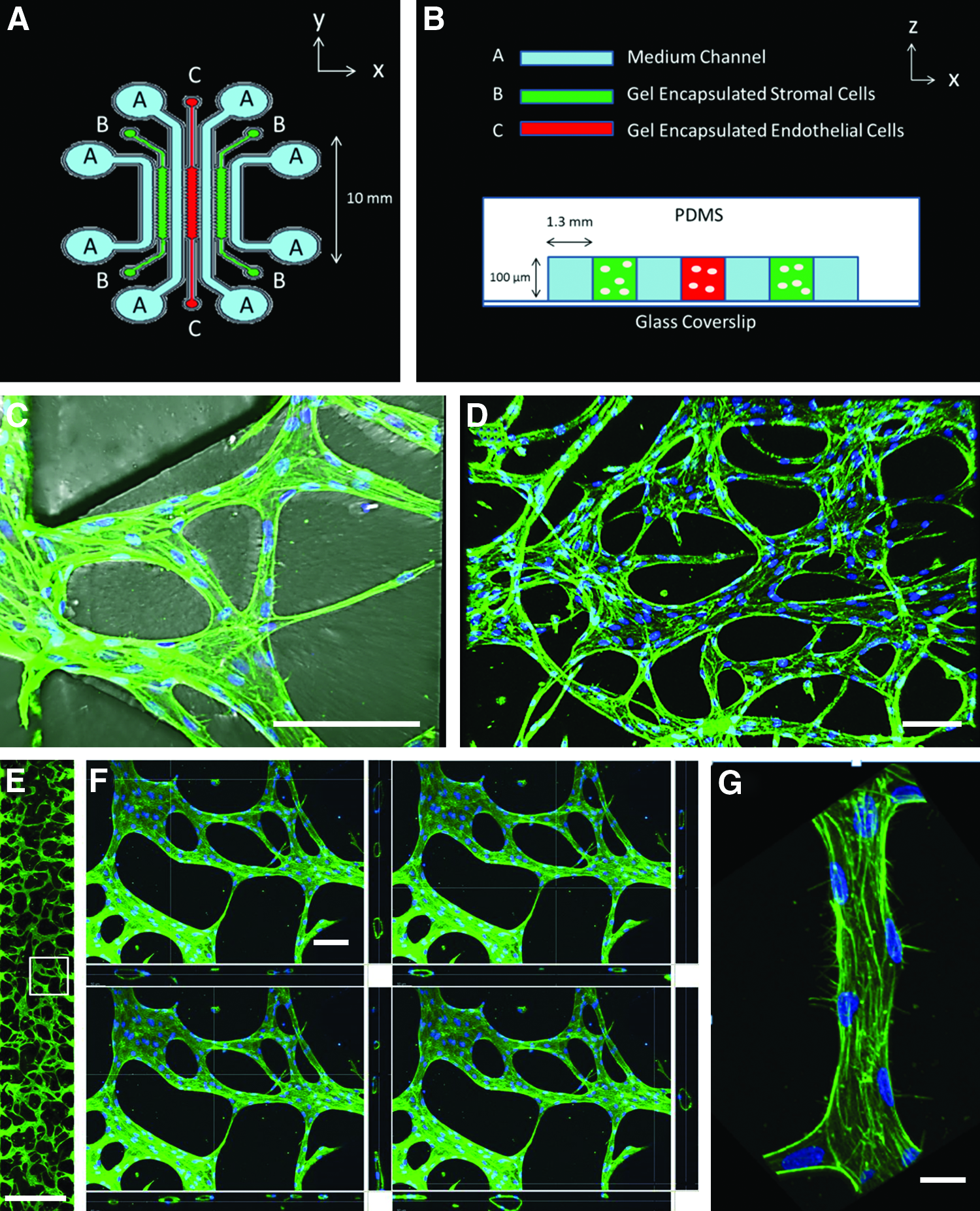
This study provides quantitative results for direct use in the design of engineered MVNs. It also demonstrates the powerful capabilities of miniaturized, perfusable, three-dimensional engineered MVNs to study the influence of a multitude of environmental cues affecting network morphology in a high-throughput and readily observable manner. It more generally demonstrates the ability to approach microvascular tissue engineering as a design problem using systematic, quantitative analysis.
Materials and Methods
Device design and fabrication
A standard procedure for generating silicone molds was used. 13 Briefly, computer aided designs (CAD) were generated and used to print negative pattern transparency masks. A 100 μm layer of SU-8 photoresist was coated onto a silicon wafer, and the mask was used to photopolymerize the pattern on to the wafer.
The mask was repeatedly used to form microfluidic chips. Briefly, PDMS (Ellsworth Adhesives) and a curing agent were mixed at a 10:1 ratio and poured onto the wafer. After degassing, the PDMS was baked in an 80°C oven for 2 h. The individual devices were then cut out and a biopsy punch was used to create ports for gel filling and medium channels. Tape was used to remove dust from the surface, and the devices were placed in an autoclave for sterilization. Clean devices and coverslips were plasma treated (Harrick Plasma) and bonded together.
The design of this multiculture vasculogenesis device was based on earlier designs from our laboratory 14 with some important adjustments: (1) a third parallel gel region was included so that stromal cells could be cultured on either side of the vascularized gel region, (2) additional medium channels were included so that each gel region is flanked by two medium channels—one on each side—to provide adequate gas exchange and supply of nutrients, and (3) the length of the device was increased to provide a larger region for vascularization.
The width of each gel and medium channel was chosen to be 1.3 mm. This was to ensure that the vascularized region's maximum dimension was at least 1 mm, such that diffusion is insufficient for delivering nutrients and removing wastes from the inner core of a living tissue. This is the length scale for which prevascularization is necessary. 15 The width as well as the length of the gel region could be increased to provide a larger vascularized area without affecting the dynamics of the system. Since the ECs and HLFs communicate through diffusive transport of secreted factors, the width of the medium channel determines the time scale for this process to occur. For typical growth factors, the time constant for diffusion over a length of 1 mm is on the order of several hours. Since the process of network formation studied in our system occurs over a period of days, this width is sufficiently small to ensure adequate time for diffusion to occur. Because the necessary time for diffusion scales with the square of the distance between the gel regions, the width of the medium channel should not be increased. Finally, the dimensions of the fibroblast gel region, in combination with cell seeding density, determine the rate of secretion of growth factors and cytokines. We chose dimensions that enabled us to screen for the optimal seeding ratio between HUVECs and NHLFs within a range of 1:2–2:1 that is physiologically relevant and reported in other in vitro studies16,17 (Results discussed below).
To achieve optimal imaging conditions, we chose a height of 100 μm for the device and sealed it with a glass coverslip. This height enabled us to form generally planar perfusable networks, allowing us to capture the MVN morphological patterns from a single focal plane using epifluorescence. The height could be increased for clinical applications, in which it might be desirable to form vascularized tissue constructs containing several planes of vessels.
Cell culture
HUVECs (Lonza) were cultured on collagen I-coated flasks (50 μg/mL collagen solution in ascetic acid for 30 min; BD Biosciences) in EGM-2 MV (Lonza) growth medium and used in experiments between passages 6–8. Normal human lung fibroblasts (NHLF; Lonza) were cultured in FBM-2 (Lonza) growth medium and used in experiments between passages 6–10.
For exogenous growth factor experiments, EGM-2 MV growth medium was supplemented with 50 ng/mL VEGF (Peprotech) and 250 nM S1P (Sigma). The medium was replenished every 2 days.
Fibrin gel cell encapsulation
Fibrinogen (Sigma) was dissolved in PBS (Lonza) at twice the final concentration (2.5–20 mg/mL for fibrin concentration experiments). Thrombin (Sigma) was dissolved in PBS at 2 U/mL. These solutions were mixed, over ice, at a 1:1 ratio to produce a final fibrinogen solution with the desired concentration (1.25–10 mg/mL). The mixture was quickly pipetted into the device using the gel filling ports. The device was placed in a humidified enclosure and allowed to polymerize at room temperature for 10 min before fresh growth medium was introduced to the medium channels. The growth medium was replaced every 2 days.
For gels with encapsulated cells, a similar procedure was followed. Cells were spun down at 1200 rpm for 5 min and the cell pellet was resuspended in EGM-2 MV growth medium containing 2 U/mL of thrombin and mixed with the fibrinogen solution at a 1:1 ratio. The device was placed in a humidified enclosure and allowed to polymerize at room temperature for 10 min before fresh growth medium was introduced to the medium channels.
Staining/imaging
After 4 days, cells in devices were fixed with 4% paraformaldehyde (Electron Microscopy Sciences) for 15 min and rinsed 3×with PBS. Triton X (0.1%; Sigma) was then introduced to permeabilize the cells and rinsed 3×with PBS after 15 min. The devices were then treated with DAPI and phalloidin (Life Technologies) for 2 h to stain the nuclei and actin, respectively. Phalloidin staining was used to image vascular network morphology with an epifluorescent microscope (Nikon Eclipse Ti-S; Nikon Instruments, Inc.). Confocal images were taken with an Olympus IX81 microscope (Olympus America, Inc.).
Analysis/quantification
The procedure used to process and analyze fluorescent images is described in detail elsewhere.
18
Briefly, raw images were prepared by enhancing contrast and removing noise. Automatic thresholding was used to binarize the images. Unconnected segments, which were not part of the perfusable networks, were automatically removed from the images. Finally, networks were skeletonized and analyzed
19
using ImageJ software (
Effective diameter calculations were performed by dividing the thresholded vascularized area by the entire length of the skeletonized network.
For perfusability experiments with fibroblasts, the regions between PDMS posts containing vessels opening to the adjacent medium channels were counted and expressed as a percentage of the total number of regions.
Bead flow experiments
For bead flow experiments, vascular networks were fixed and stained as described above. Ten micrometer fluorescent microspheres (Life Technologies) were perfused through the networks by imposing a hydrostatic pressure drop across the EC gel region. Videos were recorded using NIS-Elements software (Nikon Instruments, Inc.) on a Nikon Eclipse Ti-S microscope (Nikon Instruments, Inc.) at a frame rate of 3.37 frames per second.
Statistical analysis
All data shown represent experiments with at least n=3 individual microfluidic devices. Reported values correspond to averages over these devices for any given condition. Error bars were calculated using standard error. Significance was calculated using a two-tailed Student's t test and p-values are supplied in the Supplementary Tables S1–S6.
Results
Fibroblast stabilization
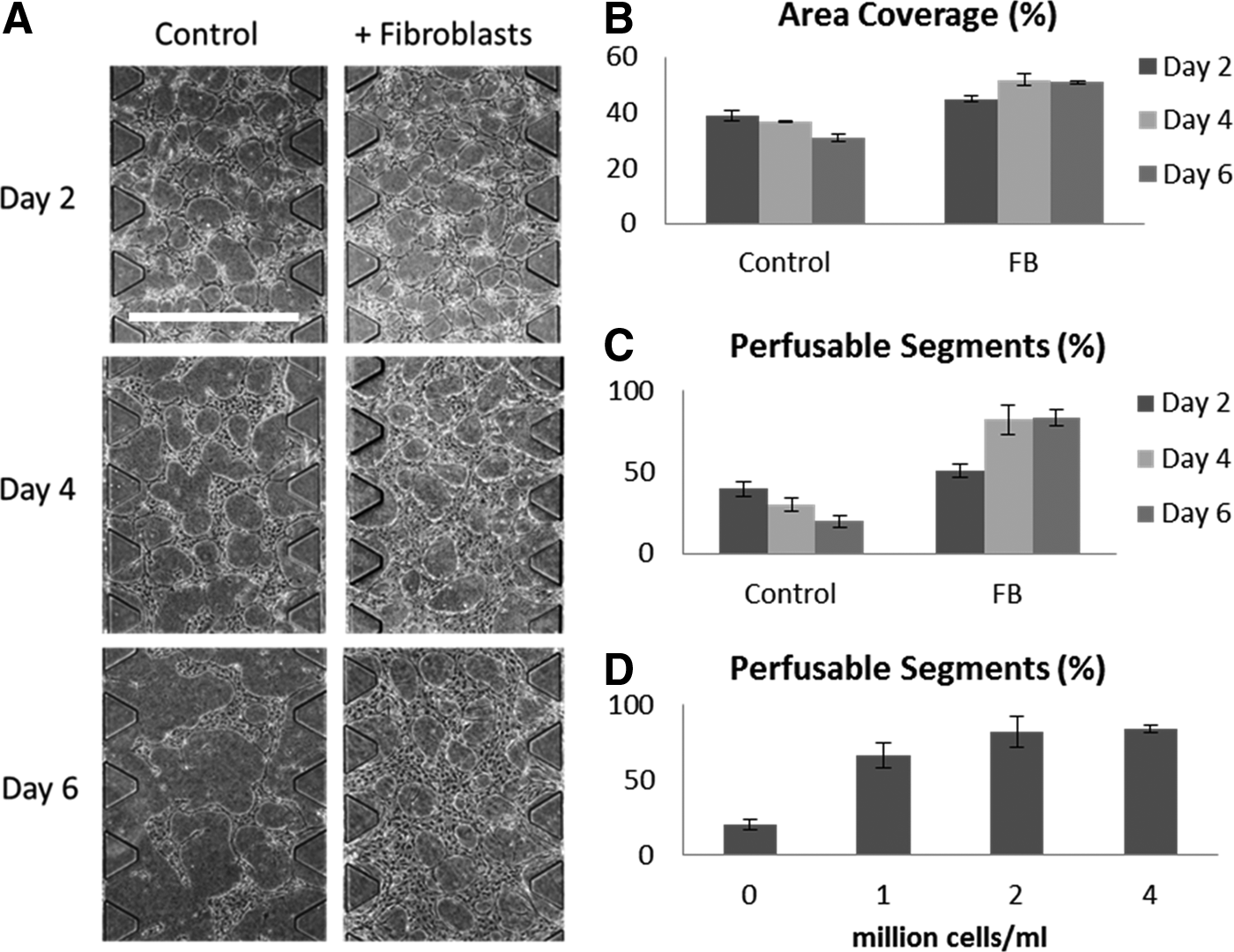
In initial experiments, we used a microfluidic device with a single gel region seeded with ECs alone (data not shown). ECs alone did form vascular networks after 2 days of culture. However, these networks subsequently regressed with time. It has been shown that coculture with fibroblasts enhances microvascular network formation in sprouting angiogenesis,20,21 vasculogenesis, 10 and in mixed coculture experiments. 11 We suspected that fibroblasts in noncontact coculture might also be important in stabilizing the nascent vascular networks before they regress. Using the revised multiculture version of our microfluidic device to encapsulate fibroblasts in the adjacent gel regions, we indeed found that the presence of fibroblasts effectively stabilized the MVNs as measured by the total vascularized area and network perfusability (Fig. 2). In the case of monoculture, these properties gradually decreased after initial network formation, consistent with our preliminary experiments. However, under coculture conditions, they reached a stable plateau after 4 days.
Paracrine signaling
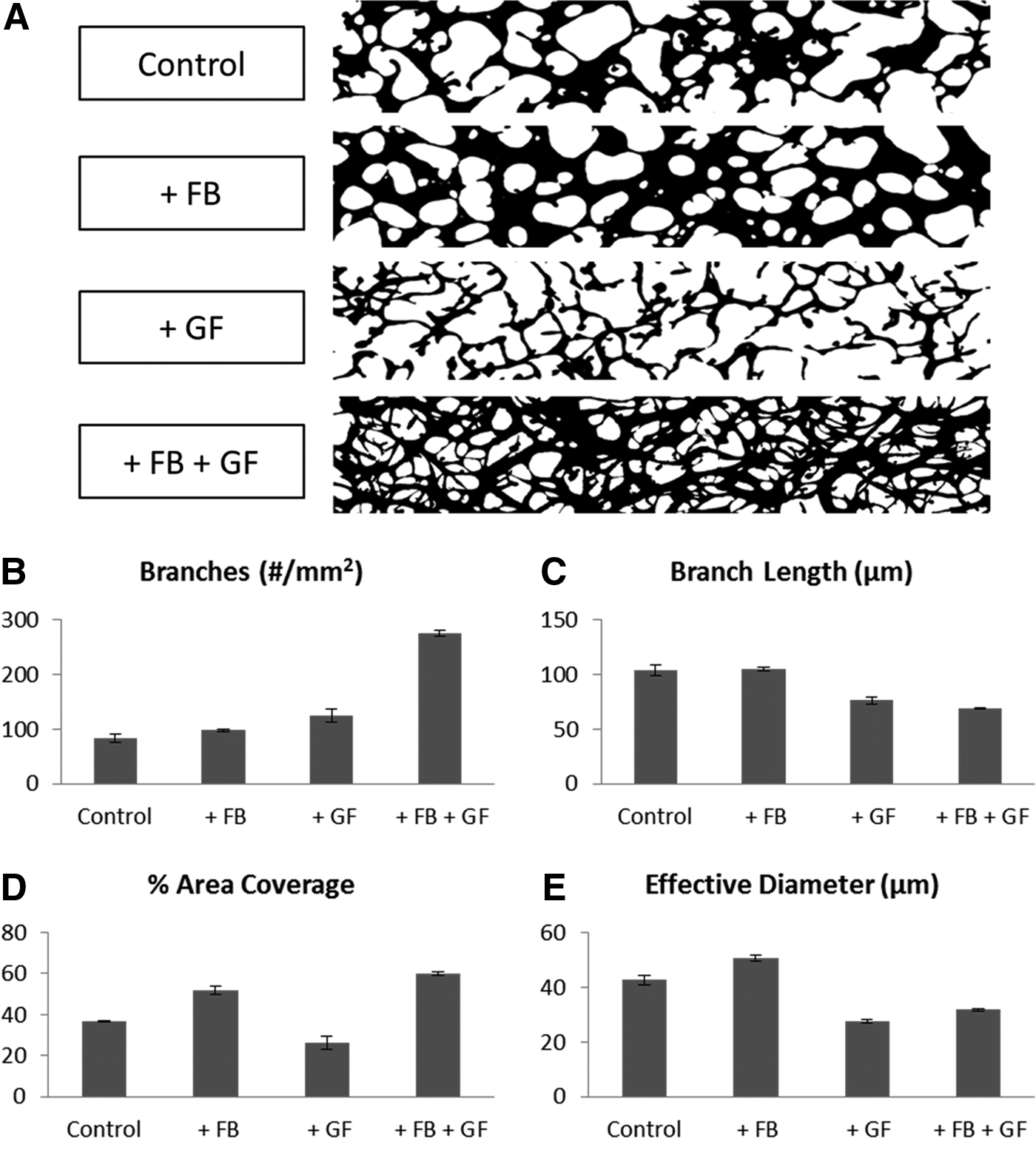
In an attempt to recapitulate the stabilizing effects of the fibroblasts, proangiogenic growth factors—VEGF (50 ng/mL) and S1P (250 nM)—were added to the growth medium. We chose these growth factors and specific concentrations based on previous work in our laboratory in which they enhanced vessel sprouting and lumen formation in an angiogenesis model. 18 These growth factors did not significantly improve the long-term stability of the networks. However, they did give rise to vascular networks with significantly different morphological characteristics (Fig. 3). The resulting networks contained more branches with shorter average length and smaller diameters than those cultured with fibroblasts or alone in EGM-2 MV growth medium. The thinner structures in these networks covered less of the total area. Interestingly, by combining fibroblast coculture with the supplemented growth factors, we were able to achieve the larger coverage area and network stability provided by fibroblast coculture, while maintaining the reduced diameters and average length induced by the angiogenic growth factors.
Fibrin concentration
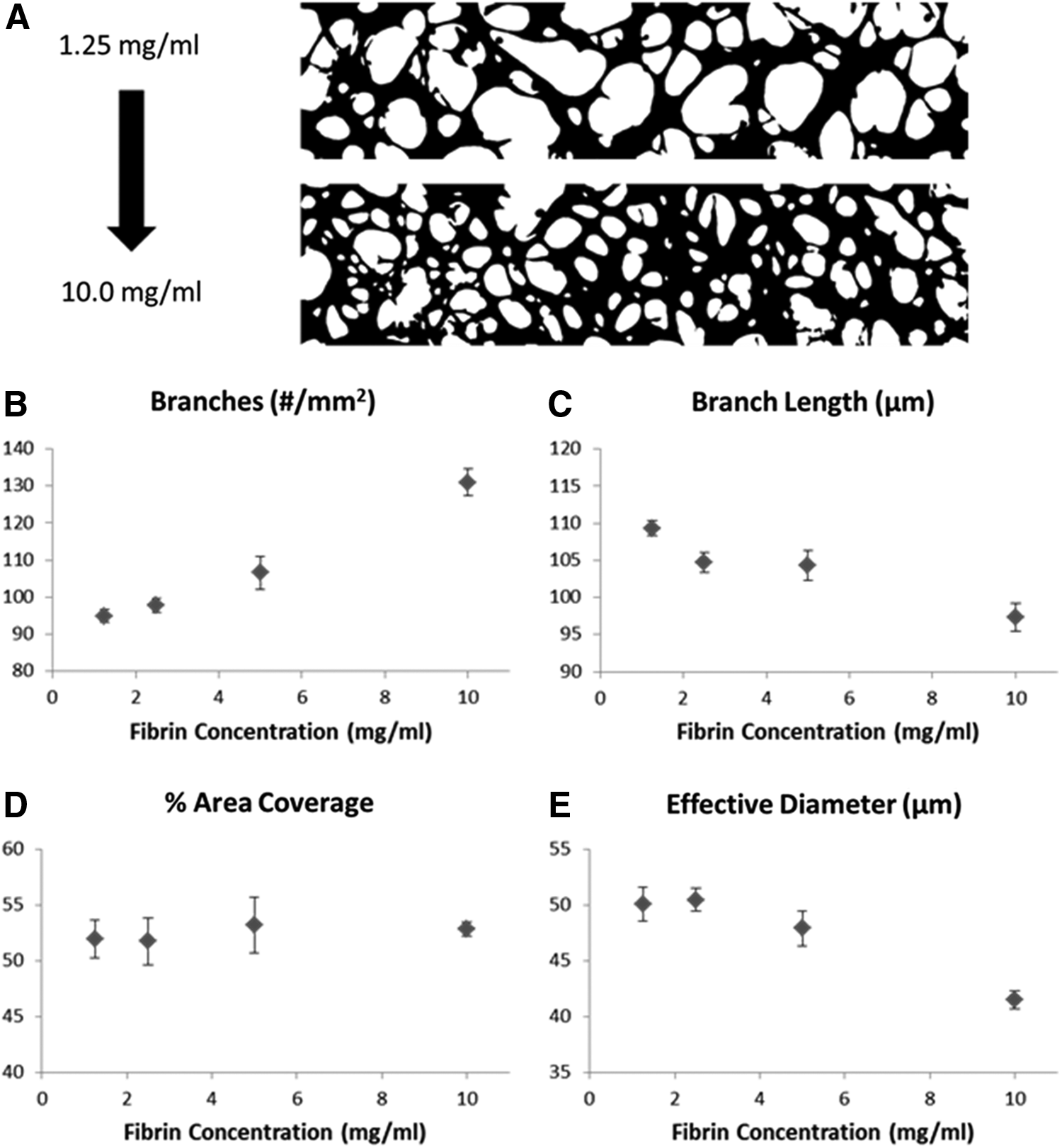
Fibrin gel was chosen for our experiments due to its biological relevance during tissue regrowth at wound healing sites and its unique mechanical properties. Specifically, fibrin gel demonstrates strain stiffening, which allows it to be easily deformed by network forming ECs at small strains, while maintaining its overall structural stability at the network level. 22 It is capable of undergoing large, recoverable deformations and can withstand considerably large strains before rupture.23–25 We chose to alter the mechanical properties of our fibrin gel by varying the fibrinogen concentration from 1.5 to 10 mg/mL. The fibrinogen concentration is a key determining factor of the mechanical structure and strength of fibrin gels and has been shown to influence MVN formation.26,27 We found that the number of branches in a network was highly sensitive to the fibrinogen concentration, while the coverage area remained little changed over the entire range of concentrations studied. Both the average branch length and effective diameter decreased with increasing fibrinogen concentrations (Fig. 4). The change in effective diameter between the two extreme conditions was 17.1%±4.6%.
HUVEC seeding density
We found that HUVEC seeding density plays an important role in MVN formation and the resulting network morphology. At a HUVEC seeding density of one million cells/mL, we found that sparse networks formed with comparatively short branches and small diameters. This represents the lowest seeding density achievable, beyond which perfusable MVNs will not form. With increasing seeding density, we found that the branch length, diameter, and area fraction of the vascularized region all increased (Fig. 5).
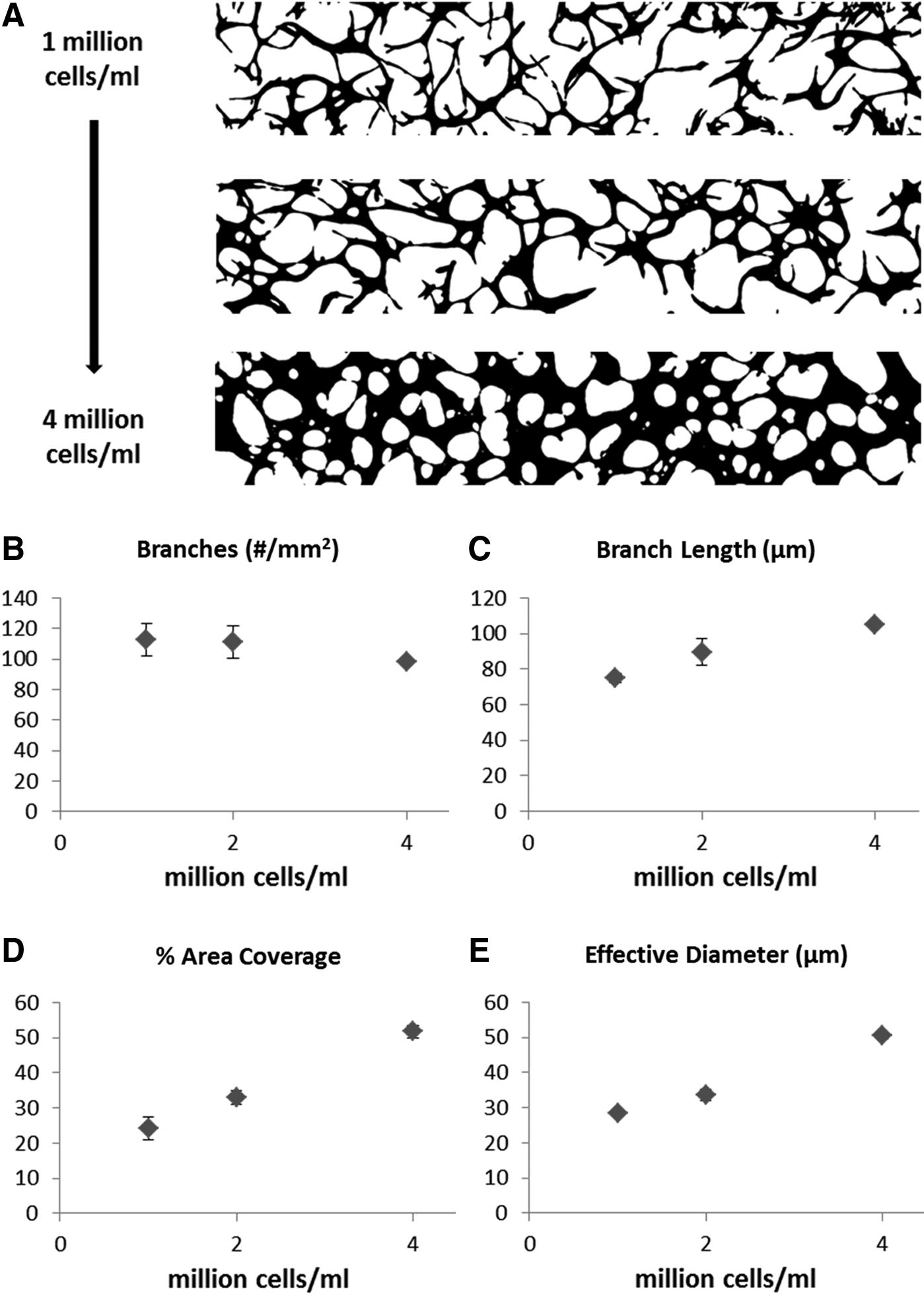
Fibroblast seeding density
The fibroblasts in our system are physically separated from the HUVECs and their effect on MVN formation is through paracrine signaling. We tested to see whether the amount of fibroblasts cultured in the stromal cell gel region had a significant impact on the engineered MVN morphology. Holding the HUVEC seeding density constant at 4 million cells/mL, we varied the amount of fibroblasts seeded from one half to twice the amount of HUVECs seeded. We found no significant differences in any of the measured MVN parameters (data not shown). Interestingly, we found that the stabilizing effect—as measured by the percentage of perfusable segments at day 6—was dose dependent on the amount of fibroblasts present and reached a steady maximum value at a HUVEC:HLF ratio of 1:1 (Fig. 2C).
Discussion
The ultimate goal of tissue engineering is to replace ailing tissues and failing organs in vivo. It has been increasingly realized that forming prevascularized tissue in vitro may be a necessary first step.15,28,29 Additionally, in vitro platforms enabling the growth and maintenance of a functional microvasculature provide significant benefits in their own right. Perfusable, in vitro models can be used as physiological drug-screening platforms for developing and identifying metastasis preventing cancer drugs.30,31 As demonstrated here, the system can be used to isolate the environmental factors influencing MVN formation and to study their effects. This is an important step in continuing to develop tissue engineering as a quantitative science and to enhance control in the design of tissue-engineered constructs. Whereas the present experiments have been performed in an in vitro system, not intended for in vivo use, we anticipate that the design principles we discovered would apply for other, similar tissue-engineered constructs. Alternatively, by nonpermanently sealing the device with a second layer of PDMS—rather than a glass coverslip—we can successfully remove the vascularized hydrogel after it forms. The removed constructs could potentially be implanted directly in vivo.
We successfully formed MVNs using our system containing vessels with diameters ranging between 1 and 100 μm. These vessels form complex, interconnected networks and are perfusable. It is known that the flow characteristics of blood perfusing through the vasculature is important in determining and maintaining network morphology. In one in vitro study, it was found that interstitial flow during vasculogenesis was necessary to form perfusable MVNs. 11 This was not the case for our system in which perfusable networks formed in the absence of flow. However, it is plausible that interstitial flow might affect the dynamics of network formation and the resulting morphology. Interstitial flow has been shown to affect cancer cell migration 32 and angiogenic sprouting through an integrin-mediated sensing mechanism, 33 which may also be important during vasculogenic network formation. In this study, we focused on preflow factors and showed that they direct the early stages of MVN formation in vasculogenesis. In future studies, it will be interesting to study the relative importance of flow through the networks after initial formation as it is also likely to play an important role. Flow-induced shear stress sensed by the endothelium acts to regulate the diameter of blood vessels in vivo. 34 It also leads to the pruning of extraneous branches eventually directing the vascular network morphology toward an energetically optimal pattern.35,36 We would expect that similar phenomena would occur in our system with the addition of flow. This could be confirmed by introducing tracer particles into the networks to visualize the flow field and correlating regions of pruned vessels to regions of limited flow. The presence of chronic shear flow 36 and the introduction of stabilizing pericytes37,38 would also provide a more authentically native microvascular environment and would likely lead to network morphologies that more closely mimic those found in vivo.
In designing vascularized tissue, it is important to consider that, in vivo, ECs form vascular networks within a complex environment involving dynamic heterotypical interactions. In addition to mechanical and signaling cues provided by the surrounding extracellular matrix, communication with supporting cell types is critical.39,40 Our microfluidic system allows for spatial segregation of various cell types in a compact environment enabling communication through the diffusion of soluble factors. By culturing the two cell types together in a single gel region, we could also study the potential stabilizing effects due to cell–cell contact and juxtracrine signaling between the two cell types and fibroblast remodeling of the ECM. It has been shown that the stabilizing effects of MSCs on MVN formation in vitro depend on the time at which MSCs are introduced into the mixed coculture system. Interestingly, MSCs only promoted network stability when introduced at least 24 h after EC seeding. 41 Seeding two cell types simultaneously in the same gel would presumably alter the chemical gradient profiles, lead to a competition for local nutrients and cytokines, and affect the structural properties of the ECM in which ECs are forming a vascular network.
The stabilizing effect of fibroblasts in our noncontact coculture system was striking. Without HLFs, the nascent EC networks quickly regressed after initial formation. However, through communication with HLFs, these nascent networks were able to maintain a stable morphology and level of perfusability. These results are in agreement with those found by Kim et al., 10 in which a lumenized vasculature was lacking after 4 days of EC culture without fibroblasts present. Our results further elucidate that vascular networks do initially form, but subsequently regress in the absence of a second cell type. It has been shown that fibroblasts enhance MVN formation through direct paracrine signaling 20 and also by adapting the ECM through secreted molecules. 21 It will be interesting in future studies to observe the ECM in real time during MVN formation to characterize this process. It will also be useful to test the effects of various other stromal cells and determine their efficacy in MVN formation for use in regenerative cell therapies.
An important finding from our study is that factors secreted from fibroblasts affect the processes of (a) network formation and (b) stabilization independently. When HUVECs were cultured with HLFs alone, the networks formed with a larger diameter than the control condition and maintained perfusability through day 6. HUVECs cultured with VEGF and S1P formed networks with a smaller diameter and regressed with time. Combining HLF coculture with VEGF and S1P produced networks with smaller diameters that remained stable. This suggests that different growth factors might independently control the individual characteristics of network morphology and that this process is also independent of network stabilization. Future work will explore the individual effects of specific growth factors known to be involved in vasculogenesis.
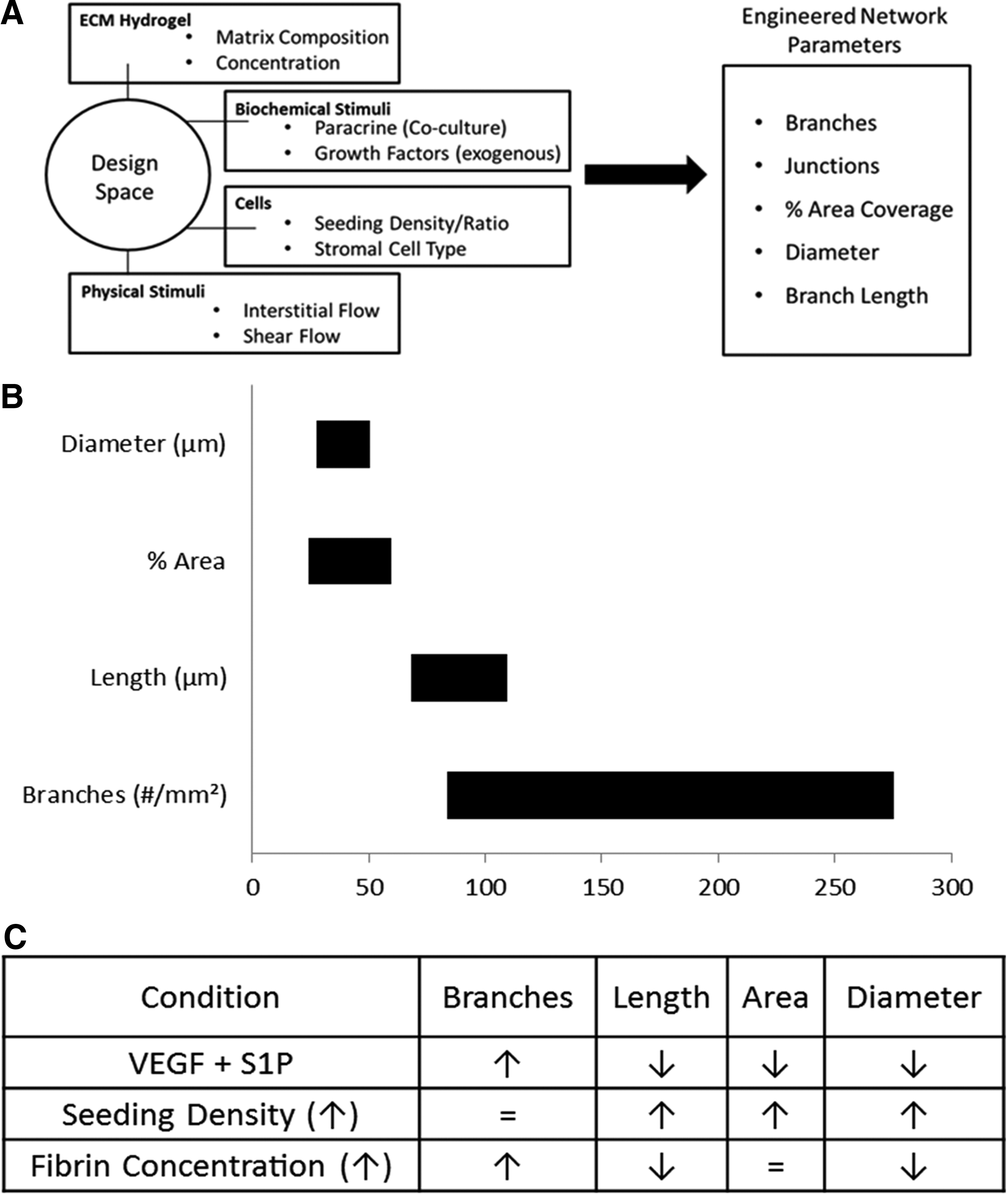
We produced a table of results (Fig. 6), which captures our design approach to microvascular engineering and summarizes our findings. In this study, we mapped out the design space for the creation of MVNs with prescribed morphological properties. The controllable inputs we studied were cell seeding densities, ECM composition, and paracrine signaling factors. The output metrics we analyzed were the number of branches, average branch length, area fraction of vascularization, and effective diameter. These properties proved sufficient to quantitatively describe the significant differences in network morphology observed by varying environmental conditions. The list of environmental factors studied here is not exhaustive, and future work will utilize this system and approach to expand in both breadth and depth. Additionally, to further probe the mechanisms underlying the observations found in this study, it would be useful to pair these experiments with a computational modeling approach.42–44 In closing, this study provides a significant contribution to the effort of quantifying the science of microvascular tissue engineering and creating MVNs with prescribed morphological properties for tissue-specific applications.
Footnotes
Acknowledgments
Support from the National Science Foundation Science and Technology Center for Emergent Behaviors of Integrated Cellular Systems (CBET-0939511) and a NSF Fellowship to JW are gratefully acknowledged.
Disclosure Statement
The authors wish to disclose that RDK has financial interests in a company, AIM Biotech, which develops microfluidic systems of the type used in this study.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.