
Abstract
Introduction:
It has been reported that cells or growth factors might improve bone regeneration in nonunion. However, the intrinsic potential of the nonunion tissue to regenerate bone is still unclear; in particular, it is not known whether there are progenitor cells within the nonunion. Thus, in this study, a clinically relevant model has been developed to investigate the nature of the cells in atrophic nonunion tissue and to assess their regenerative potential.
Materials and Methods:
Seventeen Wistar rats underwent the procedure to induce an atrophic nonunion at the tibial midshaft by stripping the periosteum and endosteum as well as creating a small (1.0 mm) noncritical gap. The fracture was stabilized with an external fixator. The proliferation ability of bone marrow-derived cells and nonunion tissue-derived cells was determined using colony forming assays. The differentiation potential of nonunion tissue-derived cells was also investigated.
Results:
Noncritical size defect nonunions were successfully induced in all of the animals. The typical characteristics of atrophic nonunions were demonstrated by radiography, micro-CT, and histology. An atrophic nonunion stimulated a systemic response in the bone marrow with an increase in bone marrow-derived mesenchymal stem cells. In contrast, cells derived from the nonunion gap were not able to form colonies. This indicated that the quiescent or inactive cells in the tissue at the atrophic nonunion gap could be induced to differentiate under osteogenic and chondrogenic conditions, but not under adipogenic conditions. Thus, these cells had the characteristics of osteochondral progenitor cells.
Discussion:
Although there was an increase in the systemic response in the atrophic nonunion model, the intrinsic potential of local tissue from the atrophic nonunion site was impaired. The cells derived from nonunion tissue could be stimulated to differentiate into bone under appropriate conditions. This suggested that there are progenitor cells in atrophic nonunion tissue, which have an intrinsic ability to regenerate bone, but the microenvironment in the atrophic nonunion site might be retarding their function. Consequently, physical or biological agents (such as growth factors or cells) are needed to reactivate the endogenous progenitor cells to make them regenerate bone in atrophic nonunions.
Introduction
T
The scintigraphy work by Weber and Cech 1 is still interpreted as suggesting that the tissue in atrophic nonunions is avascular and therefore nonvital. However, recently, it has been demonstrated that the tissues at the atrophic nonunion are well vascularized2,3 with an increased number of vessels at the fracture gap. 4 Thus, it has been suggested that other components of the fracture healing process such as the local progenitors may be insufficient.
In preclinical studies, it has been reported that an exogenous source of mesenchymal stem cells (MSCs) may improve fracture healing and bone regeneration,5–8 both by providing bone progenitor cells for regenerating bone and also potentially by providing trophic factors, which may stimulate local progenitors to contribute to fracture healing. Cells from nonunion tissue in dogs have been reported to demonstrate osteogenic potential in vitro. 9 In addition, a small case series has reported that, in some patients, the intrinsic progenitors at the nonunion site can be stimulated mechanically using an external fixator to compress the atrophic nonunion. 10 However, little is known about the local progenitors or the systemic response of progenitors to an atrophic nonunion. Hence, a study of the intrinsic progenitors in an atrophic nonunion is required to gain more information about the pathophysiology of atrophic nonunion and to develop novel treatment strategies.
This study tests the hypothesis that bone marrow progenitors respond to an atrophic nonunion and second that local progenitors in the tissue from an atrophic nonunion gap have osteogenic potential. Specifically, this study aims to determine whether there are MSCs or bone progenitors within an atrophic nonunion and whether the bone marrow at a distant site is stimulated by an atrophic nonunion. The objectives of this study are (1) to demonstrate the colony forming ability of remote progenitor bone marrow and of the local tissue from an atrophic nonunion and (2) to evaluate the differentiation potential of progenitor cells from bone marrow and from local tissue at the nonunion gap.
Materials and Methods
Animals used
This study was approved by the United Kingdom Home Office and adhered to the Animals (Scientific Procedures) Act. Twenty-one male Wistar rats (4–5 months of age) were used in this study. Seventeen animals underwent the atrophic nonunion procedure. Four animals were assigned to the control group (without procedure). They were housed under similar conditions. Two animals died because of wound complications and they were not included in the study. Thus, 15 animals remained for analysis from the experimental group. All animals were kept alive for 8 weeks after the surgical fracture procedure and at the end of the experiment; carbon dioxide inhalation and cervical dislocation were used for euthanasia.
An atrophic nonunion procedure
The surgical procedure was carried out using a previously reported technique, 4 but with modification of the fixator construct (Fig. 1). Briefly, a custom-made external fixator was applied to the right tibia. An anteromedial skin incision was made and the tibia was exposed exploiting the interval between the tibialis anterior and tibialis posterior. A 1-mm osteotomy was created at the midshaft of the tibia with a circular saw. The tibia was irrigated with 0.9% saline to prevent heat and the surrounding soft tissues were protected while the bone was cut. Next, the periosteum was stripped both proximally and distally to the osteotomy site for a length equal to one diameter of the tibia. Finally, the endosteum and intramedullary canal were curetted using a 23-gauge needle to the same length.
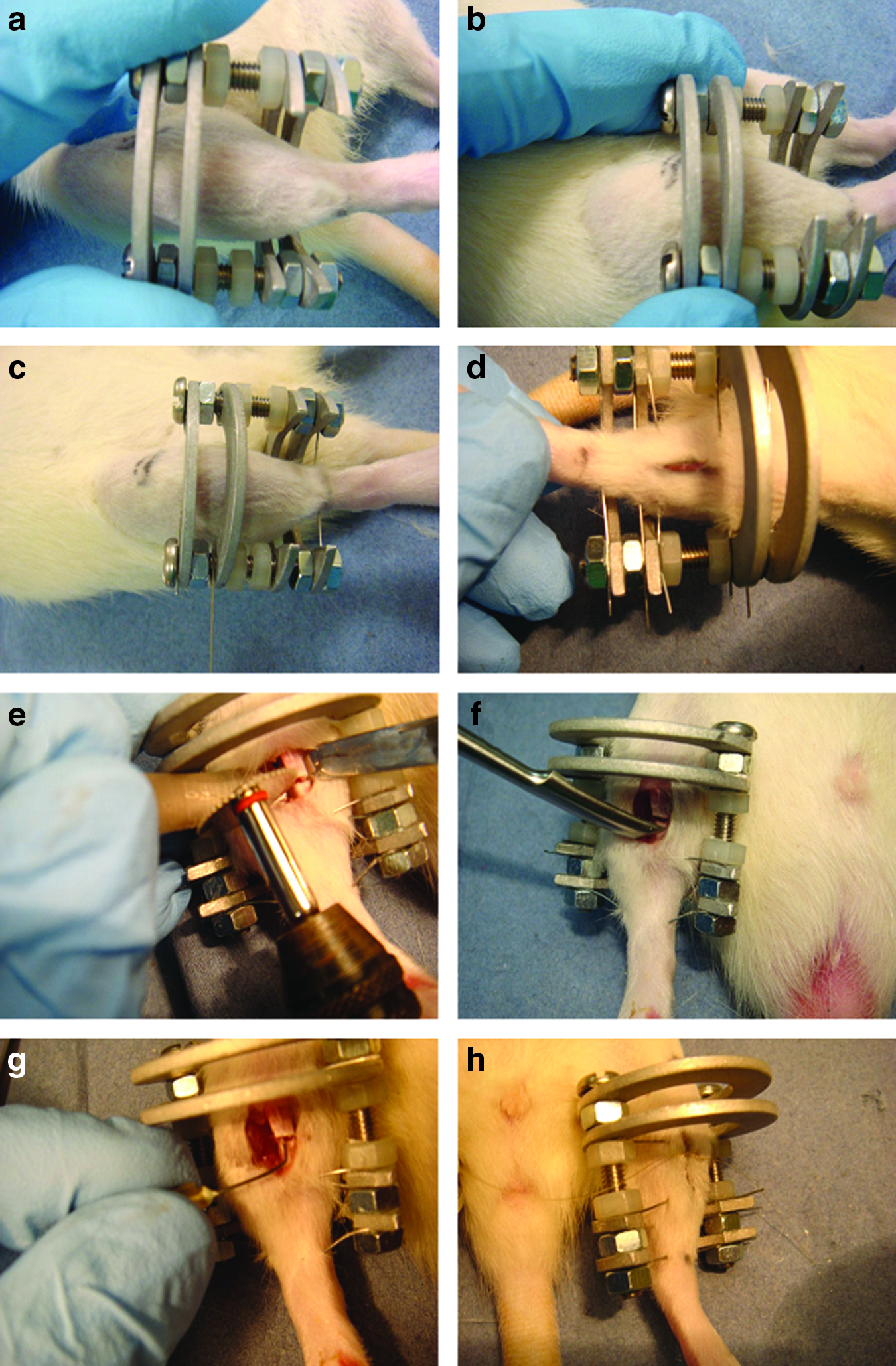
The surgical technique of an atrophic nonunion model. The position of the fixation was in the middle of the tibia; the line of knee and ankle were as the reference position
Evaluation of the atrophic nonunion of the fracture
Atrophic nonunion was diagnosed in all animals using serial radiographs with digital imaging every 2 weeks. The diagnoses were judged by two independent orthopedic surgeons. At 8 weeks, the external fixator was carefully removed, and the animals were examined by a manual clinical assessment, which confirmed the presence of motion at the fracture site. The fracture site was also scanned with a micro-CT (Skyscan Ltd.) using a voltage of 60 kV (n=7), to assess the connectivity of the fracture site and to compare the minimal bone area fraction of the fracture site to the contralateral side using a previously reported technique. 11 Four of the 15 nonunion specimens were randomly selected and processed for histological examination. Hematoxylin and eosin staining was used for general histology, Masson's trichrome for new bone formation. Stained sections at the fracture gap were viewed using light microscopy under 10×and 20×magnification.
Bone marrow isolation and colony forming assay
Bone marrow-derived MSCs from the contralateral femurs of nine atrophic nonunion rats and four age-matched normal rats (control) were isolated using a previously reported technique. 12 In brief, using a sterile technique, the soft tissues were cleaned from the femurs and then the bones were flushed using a 21-gauge needle and the cell suspension produced was filtrated with 100 μL and then 70 μL. The filtrated suspension was added to 10 mL of basal medium (10% FBS, low-glucose DMEM with 1% penicillin/streptomycin) (Gibco) and centrifuged at 1200 rpm for 5 min. The mononuclear cells were counted and processed for colony forming assessment. 13 The mononuclear cells obtained from bone marrow isolation were seeded in six-well plates at varying densities (1×105, 0.5×106, and 1×106 per cm2) in duplicate in the basal medium for 2 weeks without change of medium.
Nonunion tissue isolation and colony forming assay
To isolate the mononuclear cells from tissue at the nonunion gap (n=3), a dissociation technique was modified from the previously reported method. 14 The tissues from the atrophic nonunion gap were collected and digested using a 0.1% collagenase type 1 solution (Sigma) at 37°C for 45 min. The suspension obtained from digestion was centrifuged at 1200 rpm for 5 min. The pellet was suspended in 10 mL of standard medium and filtrated with 100 and then 70 μL. The mononuclear cells were counted and processed for colony forming assessment. An equal number of mononuclear cells from the nonunion gap were seeded directly at a concentration of 0.5×106 per cm2 in six-well plates in duplicate in the standard culture medium for 3 weeks without the medium being changed. These cultures were left for a longer period than the bone marrow cell culture to ensure that plastic adhered cells with colony forming ability were able to develop.
Giemsa staining for colony counting
Following culture of the cells for 2 weeks (bone marrow) or 3 weeks (tissues from fracture gap) without a change of media, the cultures were fixed in 100% methanol for 5 min and stained with Giemsa. The number of colonies, which contained more than 50 cells, was counted using light microscopy.
Trilineage differentiation
The isolated cells from bone marrow and nonunion tissue were maintained in culture with a media change every 3 days. The second–third passaged cells were used for the differentiation assay. To induce cell differentiation, cells were cultured under (1) osteogenic, (2) chondrongenic, and (3) adipogenic conditions as previously reported. 15 All supplements were obtained from Sigma-Aldrich® unless otherwise stated. The osteogenic medium contained the basal culture medium with a supplement of 100 nM dexamethasone, 10 mM β-glycerolphosphate, and 0.3 μM ascorbic acid. Alkaline phosphatase (ALP) deposits were detected using an ALP assay kit from Sigma-Aldrich. To demonstrate chondrogenic differentiation, a micromass culture technique was used. The chondrogenic differentiation medium contained high-glucose DMEM (4 g/L) with a supplement of 100 nM dexamethasone, 1×insulin-transferrin-selenium plus premix (ITS Premix; from BD®); final concentration: 6.25 μg/mL bovine insulin, 6.25 μg/mL transferrin, 6.25 μg/mL selenous acid, 5.33 μg/mL linoleic acid, and 1.25 μg/mL bovine serum albumin, 50 μg/mL ascorbic acid, 100 μg/mL sodium pyruvate, 50 μg/mL proline, and 20 ng/mL transforming growth factor-β3 (TGF-β3). The presence of proteoglycans was evaluated using 1% Alcian blue staining for 30 min. The adipogenic differentiation medium contained the basal medium with a supplement of 1 μM dexamethasone, 10 μg/mL insulin, 0.5 μM isobutylemethylxanthine, and 200 μM indomethacin. The cells were stained with 0.2% Oil Red O for 30 min to reveal the intracellular accumulation of lipid-rich vacuoles. The differentiation assay was performed in triplicate.
Statistical analysis
The mean of minimal bone area fraction and colony number were analyzed using either the paired or unpaired t-test as appropriate. The GraphPad Prism® 5.0 was used for analysis. The differences were considered to be statistically significant when the p-value was less than 0.05.
Results
Characteristics of the atrophic nonunion model
At 8 weeks after operation, all animals (n=15) had developed atrophic nonunions. There were two animals with wound complications 3 days after surgery, necessitating euthanasia before completion of the experiment. All of the remaining animals remained well with no pin tract infection or any signs of wound infection. The fracture gaps were grossly palpable at the osteotomy site. After removal of the external fixator, the tibias showed gross motion on clinical examination, that is, they deflected under their own weight when held horizontally. Figure 2 shows the gross morphology of an atrophic nonunion.

Atrophic nonunion model at 8 weeks. Gross morphology
In all of the animals (n=15), serial radiographs revealed a persistent gap. There was an absence of callus formation from both the proximal and the distal fragments (Fig. 3). The immediate postoperative radiographic images showed good bone alignment and the required position of the pins and the external fixator needed to maintain the 1 mm gap without significant movement of the bony fragments. Two weeks after operation, radiographic images showed no significant difference from immediate postoperative images. The 1 mm gap was maintained and there was still no callus formation and no change in the size of the fracture gap. Four weeks after operation, there continued to be no change in the radiographic appearance in all the animals. Six weeks after operation, radiographic images showed an increase in gap size in some animals. Both the proximal and the distal bone ends developed rounded blunt edges. Eight weeks after operation, the bone ends remained rounded and the gap had increased. These X-rays demonstrated the pattern of development of an atrophic nonunion.
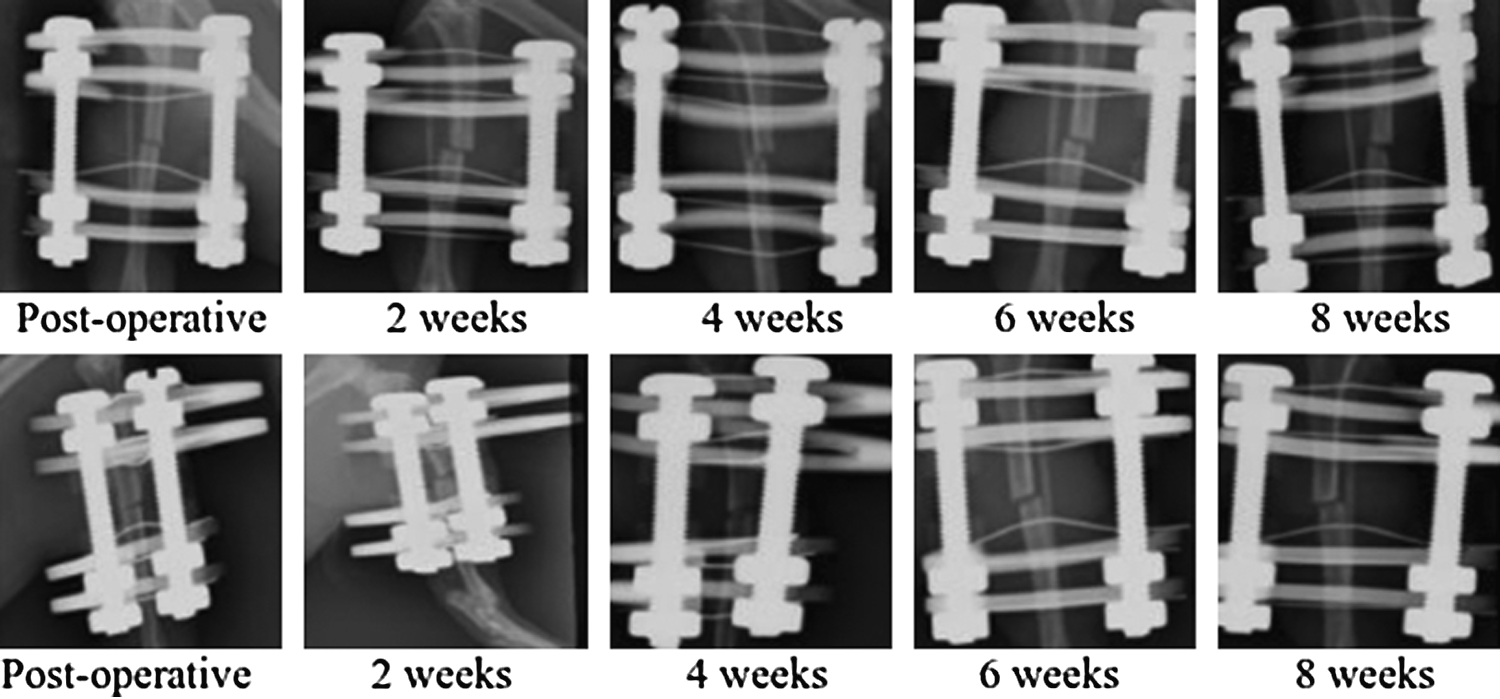
Serial radiographic; antero posterior (above) and oblique views (below) every 2 weeks after operation to 8 weeks.
Four of the 15 tibiae were randomly selected for histological assessment (Fig. 4). In all of the selected samples, the fracture site had formed an atrophic nonunion with rounded bone ends and fibrous tissue present in the interfragmentary gap. There was no woven bone or cartilaginous callus at the fracture gap. Masson's trichrome staining confirmed the absence of new bone formation.
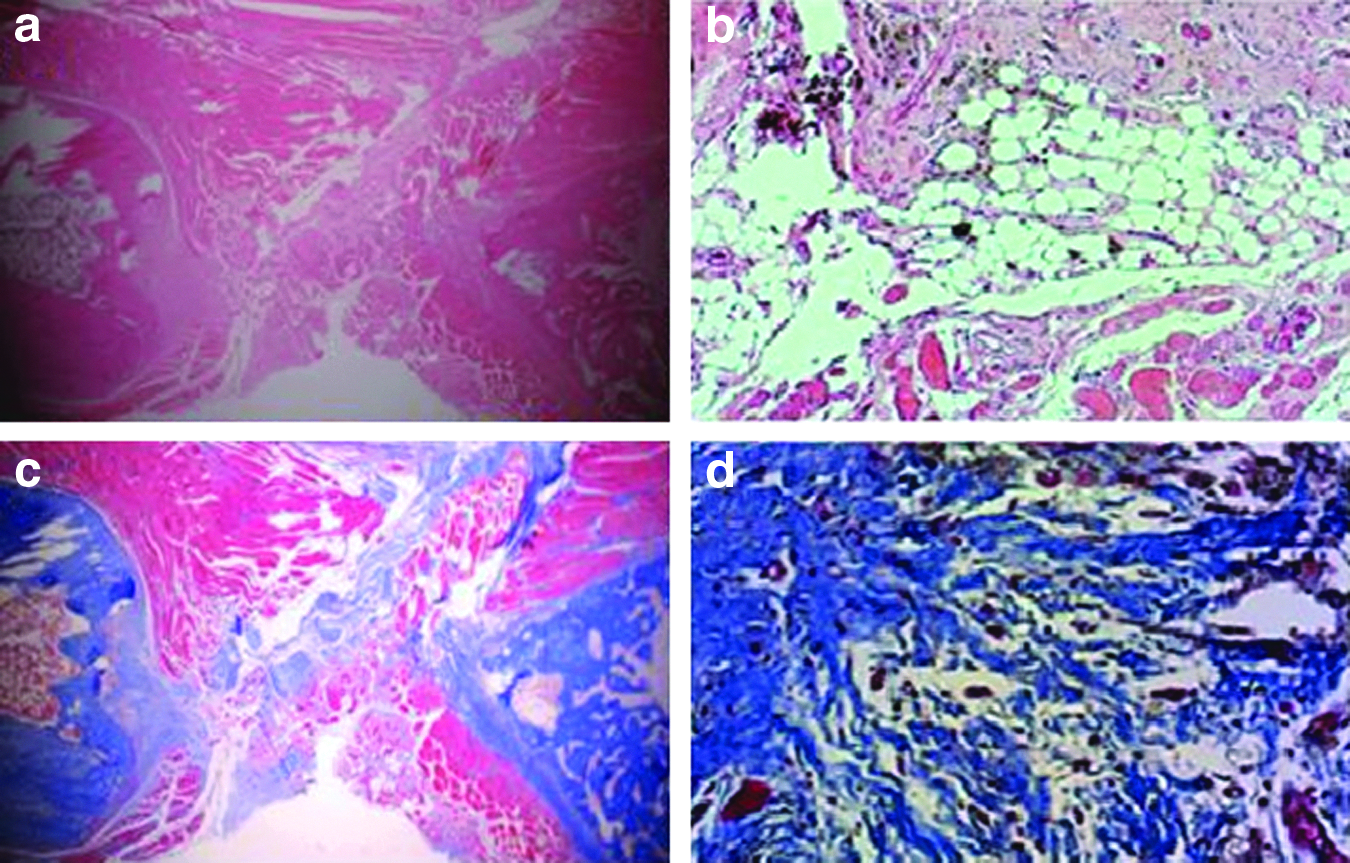
Histology assessments of atrophic nonunion model. Hematoxylin and eosin: 10×
Confirmation of atrophic nonunion with micro-CT
Three-dimensional images from micro-CT showed the fracture gap clearly. Neither an external callus formation nor bone bridges were present. The fracture ends became round, reflecting the atrophic nature of the nonunion. The shape of the nonunion was also demonstrated by the bone area fraction, that is, the fraction of area formed by the external borders of the diaphysis and the continuation of these borders across the fracture site that was filled with bone. This was plotted across the fracture site from proximally to distally along the region of interest. Proximal and distal to the atrophic nonunion, the bone area fraction of the proximal fragment was approximately 0.9. The bone area fraction dropped to a minimum at the fracture site (Fig. 5). The bone area fraction from the region of interest in the atrophic nonunion was significantly less than from the equivalent position from the unoperated side (0.84 vs. 0.12, p<0.01)

Bone area fraction from proximal to distal part demonstrated by micro-CT; normal bone
Colony forming units of MSCs in an atrophic nonunion
Assessment of the increase in the number of colony forming units from bone marrow from a remote site to the atrophic nonunion
The densities of colonies of the well plates seeded at 5×105 cells (0.5×105 cells per cm2) were found to be appropriate for counting. The colony forming units from both atrophic nonunion and normal controls had a similar morphology and the fibroblast-like cells in the colonies were also similar. The number of colony forming unit counts from the remote bone marrow of atrophic nonunion animals was significantly higher compared with the controls. The mean number of colony forming from the atrophic nonunion group was 10.4±1.1 (mean±SEM, n=9), whereas for the control group, it was 5.8±0.7 (mean±SEM, n=4); (p-value=0.0041) (Fig. 6).
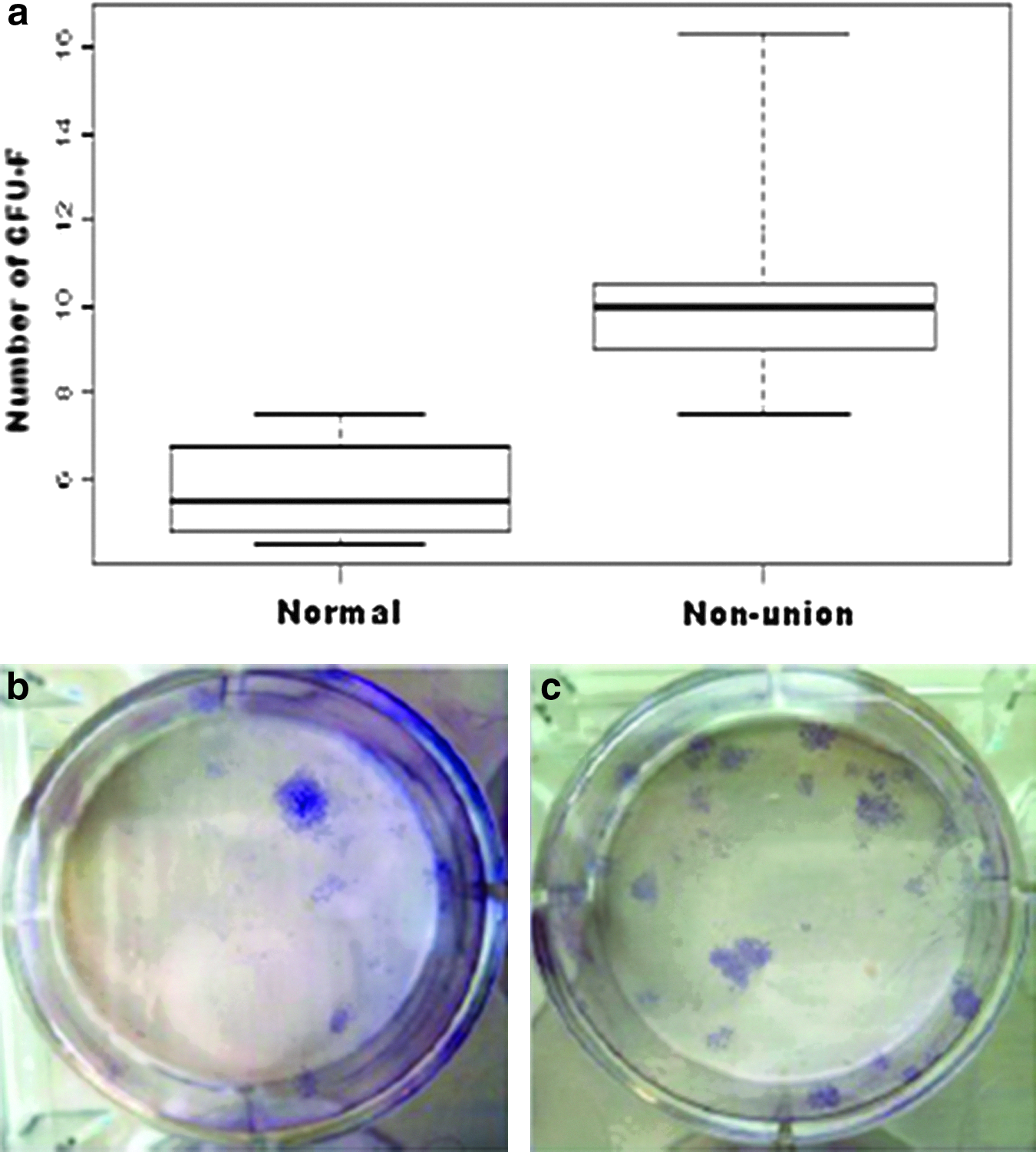
The colony forming assay for remote bone marrow from the atrophic nonunion model. The mean number of colony forming units from the bone marrow from the contralateral femur of the atrophic nonunions and also for normal control animals
Assessment of atrophic nonunion tissue for colony forming units of mononuclear cells
Atrophic nonunion tissues from three samples were dissociated with collagenase type 1 and colony forming assays were performed. Following digestion of the tissues with collagenase, 500,000 mononuclear cells from the tissue from the atrophic nonunion were seeded onto six-well plates (0.5×105 cells per cm2) and the media were left unchanged. No colony forming units were obtained from any of the samples.
The differentiation potential of progenitor cells in atrophic nonunion model
Cells were successfully derived from the femoral bone marrow of all of the samples (n=4/4), whereas cells could only be isolated from three out of five of samples (n=3/5) from atrophic nonunion tissue. Cultured cells that had undergone two to three passages from both bone marrow and nonunion tissue were induced under osteogenic, chondrogenic, and adipogenic conditions. Cells derived from bone marrow demonstrated trilineage differentiation potential and therefore could be considered to be MSCs according to the International Society for Stem Cell Research (ISSCR). 16 Cultured cells from the atrophic nonunion tissue were only capable of osteogenesis and chondrogenesis, but not adipogenesis (Table 1). Thus, the cells derived from the nonunion tissue could be defined as osteochondral progenitor cells.
The osteogenic (alkaline phosphatase staining), chondrogenic (Alcian blue staining), and adipogenic (Oil Red O staining) differentiation potential are displayed. Color images available online at
Discussion and Conclusion
The aim of this study was to determine the colony forming ability of cells (1) from nonunion tissue and (2) from remote bone marrow in an atrophic nonunion model. The results demonstrated that the number of colony forming units from the remote bone marrow increased in the atrophic nonunion model, 8 weeks after operation. No colony forming cells could be isolated from atrophic nonunion tissues. However, culture cells from nonunion tissue were capable of osteogenesis and chondrogenesis, but not adipogenesis.
A number of studies have evaluated the bone regeneration potential in a segmental defect model,5–7 however, these defect models do not simulate either the biological or mechanical condition of a clinical nonunion. The induction of a noncritical defect atrophic nonunion can be performed by cauterization, or introducing of foreign material. The technique of cauterization for inducing an atrophic nonunion was reported by Kaspar et al. 17 in a rat femur model. The animals had a monolateral external fixator applied and a femoral osteotomy. They used cautery to damage the perisoteum and the tissue around the fracture sites. However, thermal destruction of the periosteum is not relevant clinically. Interposition of foreign material has been used to create an atrophic nonunion in small and large animal models. It has been reported that the silicone foil used to wrap around the femoral diaphysis was a reliable technique to induce atrophic nonunion. 11 However, it is also not representative of an atrophic nonunion in the clinical setting.
This study was conducted using a well characterized noncritical size defect model. It represented the clinical scenario of a stiff atrophic nonunion. 18 The surgical technique was modified from that originally developed by Reed et al. 4 The fixator constructs were made from aluminium as the rats chewed the plastic rings. There was no fixation failure from this design of the external fixator. This model of an atrophic nonunion did not require a large segmental defect. The model was induced by creating a noncritical size 1 mm gap in the tibial midshaft in association with periosteal stripping and medullary canal curettage for a distance of one tibial diameter. This procedure was reproducible and reliable. All of the operated animals developed atrophic nonunions. The diagnosis was confirmed with serial radiographs and histology. The histology from the fracture gap showed fibrous tissue across the fracture site, which provided a certain amount of stiffness at the nonunion site. Thus, this model should be classified as a stiff atrophic nonunion model according to the review by Mills and Simpson. 17 Furthermore, micro-CT was used to demonstrate the connectivity across the fracture site. From the CT images, the bone fraction area was derived. This technique provided a quantitative measurement of the shape of the fracture repair. 11
The regeneration potential of the native progenitors from remote bone marrow (the contralateral femur) of animals with an atrophic nonunion model of tibia was investigated. The number of MSCs present was determined using their characteristic of plastic adherence and their ability to form colonies when seeded at low densities. 13 The results showed that the colony forming capacity from remote bone marrow (contralateral femur) of animals with an atrophic nonunion was significantly higher than normal animals. This study has shown that remote bone marrow yields an increased number of MSCs in the presence of an atrophic nonunion. Thus, the nonunion appeared to stimulate a systemic response. A clinical study of Marchelli et al., has demonstrated that the serum OPG, which is a bone formation marker, was significantly higher in patients with an atrophic nonunion of a shaft fracture indicating that there was a systemic response to the nonunion. 19 However, it has been reported that the number of colony forming units from bone marrow in an established atrophic nonunion patient was significantly less than in the normal control patient. 20 It is possible that the systemic response to the fracture or nonunion may depend on the stage of the healing process 21 and the results from one time point cannot be extrapolated to other time points.
Finally, the local cells in the nonunion tissue were investigated. Colony forming units could not be detected from cells isolated from the tissue at the atrophic nonunion gap. In this study, enzyme digestion using a collagenase solution, which is a common method for cell isolation, was used to isolate the cells from nonunion tissue. It has been reported that the collagenase solution can be used to extract the adipose-derived stem cells from fat, 22 and the tissue-derived stem cells from the periosteum and synovium.23,24 The cell viability and cell number were also checked using 0.4% tryphan blue solution before seeding. It was postulated that the systemic response in atrophic nonunion patients might be higher, whereas the local tissue was unable to contribute sufficiently to the healing process as the local MSCs and progenitors were insufficient. In this study, atrophic nonunion tissue-derived cells could be isolated from three out of five cases. They grew more slowly than bone marrow-derived MSC and also they did not have the trilineage differentiation capacity, whereas the cells that had been derived from remote bone marrow were able to differentiate into bone, cartilage, and adipose. Based on the results from this study, it is asserted that the nonunion-derived cells were not MSCs as they were not able to form colonies and they did not have multipotent differentiation. It has been reported that cells isolated from human nonunion tissues demonstrated increased levels of cell senescence and reduced capacity to form osteoblasts, which were associated with significantly elevated secretion of Dickkopf-1 (Dkk-1). 14 In addition, there were significantly downregulated factors, including canonical Wnt-, IGF-, TGF-β-, and FGF-signaling pathways in nonunion osteoblasts. These factors have been reported to be involved in the proliferation and differentiation of bone progenitors. 25 The results presented here suggest that the tissue from atrophic nonunions has an insufficient number of functional progenitors and that the cells at the nonunion site might be committed osteochondral progenitors, but not MSCs. However, these cells might have a potential role to contribute to bone healing under appropriate mechanical or biological stimulation.
Atrophic nonunions are sometimes considered to be avascular. However, the mean vessel counts of biopsies from the fracture gap of patients with healing fractures, hypertrophic nonunions, and atrophic nonunions demonstrated no difference in vascularity between the three groups. 2 These findings were supported by preclinical models, which yielded consistent results demonstrating that the number of vessels was not reduced in atrophic nonunion tissue compared with normal healing.3,4 The results from our study extend our understanding of the pathophysiology of atrophic nonunion and suggest that atrophic nonunions may fail to heal, because of the biological impairment of progenitors at the local site.
There were some limitations in this study. First, this study did not demonstrate the temporal change of yield of MSCs in bone marrow during the developing of an atrophic nonunion. Second, the characteristics of isolated cells were based on their general morphology and their function in the in vitro study, and this might not reflect the in vivo characteristic of the cells in the model. Although the markers of MSCs in vitro have been investigated, 26 the identification of MSCs in vivo is still controversial. 27 If markers of in vivo MSC markers could be specifically identified, this would allow researchers to have a better understanding of the pathobiology of MSCs in atrophic nonunion.
In conclusion, in atrophic nonunion patients, (1) there was an elevated systemic response of the MSCs of remote bone marrow and (2) the local cells derived from the nonunion tissues were capable of osteogenesis and chondrogenesis, but not adipogenesis, indicating that they were osteochondral progenitor cells rather than stem cells.
Footnotes
Acknowledgments
The experimental work presented in this article was partly performed in the Scottish Centre for Regenerative Medicine (SCRM) Building and Queen medical research institution (QMRI), the University of Edinburgh, United Kingdom.
Disclosure Statement
No competing financial interests exist.