
Abstract
In vitro culturing of primary multiple myeloma cells (MMC) has been a major challenge as this plasma cell malignancy depends on the bone marrow environment for its survival. Using a microfluidic platform to emulate the dynamic physiology of the bone marrow microenvironment, we report here a new approach for culturing difficult to preserve primary human MMC. The system uses a three-dimensional ossified tissue to mimic the tumor niche and recapitulate interactions between bone marrow cells and osteoblasts (OSB). To this end, the human fetal OSB cell line hFOB 1.19 was cultured in an eight-chamber microfluidic culture device to facilitate the seeding of mononuclear cells from bone marrow aspirates from three multiple myeloma patients. Optical microscopy, used for real-time monitoring of mononuclear cell interactions with the ossified tissue, confirmed that these are drawn toward the OSB layer. After 3 weeks, cocultures were characterized by flow cytometry to evaluate the amount of expansion of primary MMC (with CD138+ and CD38+CD56+ phenotypes) in this system. For each of the three patients analyzed, bone marrow mononuclear cells underwent, on an average, 2 to 5 expansions; CD38+CD56+ cells underwent 1 to 3 expansions and CD138+ cells underwent 2.5 to 4.6 expansions. This approach is expected to provide a new avenue that can facilitate: (1) testing of personalized therapeutics for multiple myeloma patients; (2) evaluation of new drugs without the need for costly animal models; and (3) studying the biology of multiple myeloma, and in particular, the mechanisms responsible for drug resistance and relapse.
Introduction
M
The replication of the intricate multiple myeloma niche has been examined using both in vitro systems and in vivo models. Yaccoby and Epstein developed the widely used xenograft model capable of supporting the growth of human myeloma,5,6 using a severe combined immunodeficient (SCID) mouse implanted with human fetal bone fragments to create a humanized bone microenvironment (SCIDhu). Animal experiments, however, are time-consuming, costly, and have limited throughput capacity for drug evaluation. Recently, Kirshner et al. 7 developed a three-dimensional (3D) culture system with several key niche elements that allowed in vitro expansion of MMC. However, the conventional macroscale static culture nature of this approach is not suitable for assessing the potential contribution of perfusion-related mass transfer and shear stress effects on the dynamic progression of multiple myeloma tumors. The interactions of MMC within their niche, in a perfused environment that can be controlled to emulate blood flow variations due to vascularization changes and recapitulates the spatial and temporal characteristics of the complex tumor microenvironment, has yet to be incorporated and examined in in vitro models of multiple myeloma.
In this report, we describe the development of a physiological and robust technology for preserving the bone marrow-MMC interactions by customizing a microfluidic 3D ossified tissue system (Fig. 1), which was originally designed to evaluate the efficacy of biomaterials aimed at accelerating orthopedic implant-related wound healing.8,9 We found that this novel patient-specific in vitro multiple myeloma tissue model permits the preservation and proliferation of primary human MMC and recapitulated essential interactions between malignant cells and the recreated endosteal niche, all within a perfused microenvironment intended to mimic the fluidic features of the bone marrow.

Schematics of 3D microfluidic ossified tissue culture system and procedures.
Materials and Methods
Microfluidic chamber
The fabrication and operational procedures of the microfludic device are described elsewhere in detail.8–10 In brief, a microfluidic housing with eight culture chambers shaped as hexagonal prisms (12 mm in length, 6 mm in width, 0.2 mm in depth, and 10 μL in volume) was mounted on a glass slide (Fig. 1). The housing was made out of polydimethylsiloxane (PDMS; Dow Corning Corp.) by soft lithography. PDMS was used as a primary construction material for its biocompatibility and O2 and CO2 permeability desired for long-term cell culture inside a conventional incubator. To ensure that bone marrow mononuclear cells (BMMC) were not flushed out of the culture chambers (Fig. 1), a poly (ɛ-caprolactone) (PCL) fiber mesh was placed as a barrier near the chamber outlet. The chambers were coated with 0.1 mg/mL human fibronectin (Becton, Dickinson and Company) to facilitate OSB seeding and attachment.
Construction of ossified tissue scaffold
To build the 3D ossified tissue, 2×104 human osteoblasts (hFOB 1.19, CRL-11372, purchased from American Type Culture Collection, ATCC, LGC Standards) were cultured in each chamber (of 10 μL volume) for 4 days before the addition of BMMC. The OSB growth medium (Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 1% penicillin/streptomycin) (50 mg) was pumped into each chamber at a flow rate of 0.8 μL/min using a syringe pump (KDS230; KD Scientific).
Patient's BMMC cocultured with 3D ossified tissue
This study was conducted in accordance with the Institutional Review Board–approved protocols at the Hackensack University Medical Center and in accordance with the Declaration of Helsinki. Mononuclear cells and plasma were isolated from three multiple myeloma patient's bone marrow aspirate by Ficoll-Paque gradient centrifugation. Patient disease information is provided in Table 1. BMMC were frozen and stored in liquid nitrogen. Bone marrow culture media (RPMI with L-glutamine, 6.2×10−4 M of CaCl2, 1×10−6 M sodium succinate, 1×10−6 M hydrocortisone, 1 unit/mL heparin) was supplemented with 10% pooled plasma from 10 patients. For proliferation assays, BMMC samples were labeled with 0.5 μM carboxyfluorescein diacetate succinimidyl ester [CFSE; Life Technologies]. Approximately 2×104 BMMC were introduced into the ossified tissue culture chambers. The flow was stopped for 4 h to allow settling of cells and reinstated with the bone marrow growth medium. Microfluidic cultures were kept in a humidified incubator at 37°C in 5% CO2. At the conclusion of the culture, cells were isolated from the chambers by trypsinization. The harvested tissue was dissociated by grinding in microcentrifuge tubes. Single cell suspensions were prepared by filtering the grinded tissue through a 40-μm cell strainer.
Data presented are the average result from two different chambers±SD.
Stage according to the Durie Salmon staging.
% MMC in culture versus corresponding value on day 7 (p<0.05).
% BMMC in culture versus corresponding value on day 7 (p<0.05).
BMMC, bone marrow mononuclear cells; CFSE, carboxyfluorescein diacetate succinimidyl ester; MMC, multiple myeloma cells; 3D, three-dimensional; N/A, data not available.
Real-time monitoring
The interactions of BMMC with the 3D ossified tissue were monitored by real-time microscopy with the technique described elsewhere in detail. 10 Briefly, the microfluidic device was placed on an automated stage of an inverted microscope (Nikon Ti-E) after BMMC seeding. The microscope stage was housed in an environmental chamber (In Vivo Scientific) and maintained at 37°C, 5% CO2, and >90% humidity. To examine behavior of BM cells on the 3D bone tissue, brightfield and florescent images were captured every 20 min to generate time-lapse videos for the first 15 h. In addition, brightfield and florescent images were captured on days 0, 7, and 21. The brightfield and florescent images for the same position were merged into one image.
Confocal microscopy
A confocal laser scanning microscope (Nikon E1000 with Nikon C1-Plus confocal system) was used to examine the cultured tissue. The microfluidic device was placed on the stage of the confocal microscope as is and confocal z-slices were imaged with a 20×objective lens. Three-dimensional reconstructions were performed using Nikon EZ C1 analysis software. Using fluorescence, CFSE-labeled BMMC were identified and could be distinguished from the 3D ossified tissue scaffold.
Flow cytometry and cell proliferation measurements
For flow cytometric analysis, cells were stained in FACS buffer (PBS containing 1% BSA and 0.02% sodium azide) with the following fluorochrome-conjugated mAb (purchased from Beckman Counter or BD Pharmingen): anti-CD138 PE-Cy5, anti-CD38 PE-Cy5, and anti-CD56 PE. A minimum of 10,000 events were acquired and analyzed using a Beckman Coulter FC500 flow cytometer and the CXP cytometer analysis software (Beckman Coulter).
CFSE+ cells in dot plots were gated as BMMC. CFSE+CD38+CD56+ cells and CFSE+CD138+ cells were gated and identified as multiple myeloma subpopulations. MMC proliferation was determined by analyzing the CFSE intensity, as each cell division results in a 50% decrease in the florescence intensity of this dye. Division peaks were labeled from 0 to n. Since a single cell dividing n times will generate 2
n
daughter cells, and if the total number of cells resulting from three divisions (n=3) is eight, then exactly one mother cell had to divide three times to generate these eight daughter cells (23=8)
11
. Making use of this mathematical relationship, the number of divisions of mother cells was extrapolated from the number of daughters under each division peak. The BMMC daughter cell percentage under each cell division peak was obtained from the flow cytometry histogram analysis. The average cell expansion was then calculated as follows:
Where
Statistical analysis
Flow cytometric data for each patient are expressed as mean±standard deviation of the percentage of MMC (from dot plots) obtained from two separate chambers at each time point. For multiple comparisons (>2 groups), one-way ANOVA followed by the Tukey's multiple comparison test was performed to determine significant differences among groups. The Student's t-test was used when comparing two groups. A p<0.05 was considered significant. Prism (GraphPad software) was used to perform analyses.
Results
Interaction between patient-derived BMMC and the 3D ossified tissue
The 3D ossified tissue in our model represents the bone marrow endosteal surface. Its growth can be described as follows: (1) adhesion and spreading of OSB and formation of a confluent layer as early as 24 h after seeding, (2) stacking of cells by day 2, (3) production of measurable extracellular matrix (ECM) and introduction of patient CFSE-labeled BMMC on day 4.
Temporal behavior of cells in 3D culture evaluated by time-lapsed images revealed that, during the first 4 h after seeding, BMMC settled on the external perimeter of the 3D ossified tissue; between 4–8 h, BMMC were seen to adhere to the OSB layer. Subsequently, cells integrated themselves into the forming ossified tissue (∼8 h) and became uniformly dispersed within its surface in the first 24 h (Supplementary Video S1; Supplementary Data are available online at
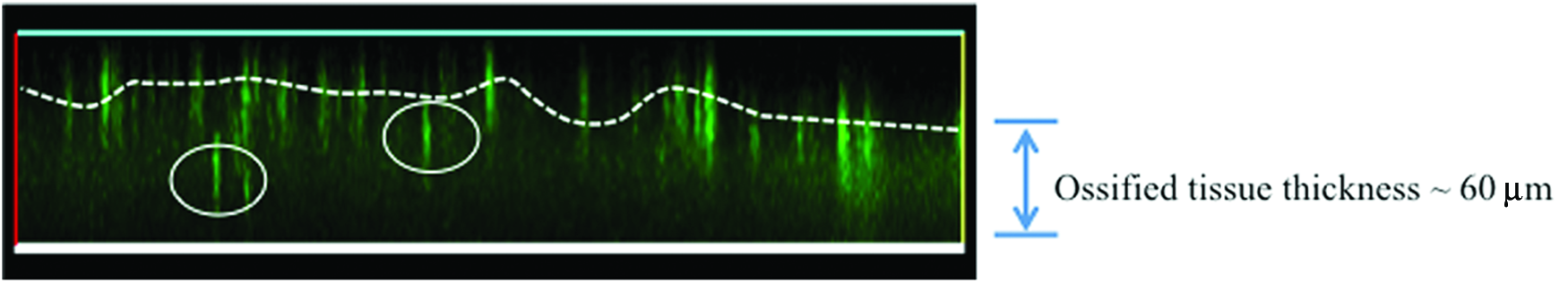
Confocal imaging of BMMC in the 3D ossified tissue. The stratification of BMMC in the 3D tissue was assessed by confocal microscopy (day 21). Diffused green area represents the ECM thickness of the 3D bone tissue (∼60 μm). CFSE+BMMC can be seen as bright green spots. The dashed white line follows average location of BMMC in the upper surface of the 3D tissue. On day 21, cells could be found deeper in the tissue (white circles). ECM, extracellular matrix. Color images available online at
Representative fluorescent microscopy images in Figure 3a and b show that BMMC were well dispersed and had a uniform CFSE intensity by day 1. From day 7, divided BMMC were darker as the CFSE intensity decreased due to proliferation (Fig. 3c). Some spreading of BMMC was also observed (insert on Fig. 3c), suggesting close interaction of these cells with the underlying OSB/ECM substrate. By day 21, most of the BMMC had divided and displayed a much decreased CFSE intensity (i.e., CFSElow) and only those cells that had retained their CFSE labeling could be detected. Cell colonies containing 20 to 40 cells were observed (insert on Fig. 3d), indicating that some BMMC underwent four to five rounds of expansion over 21 days.
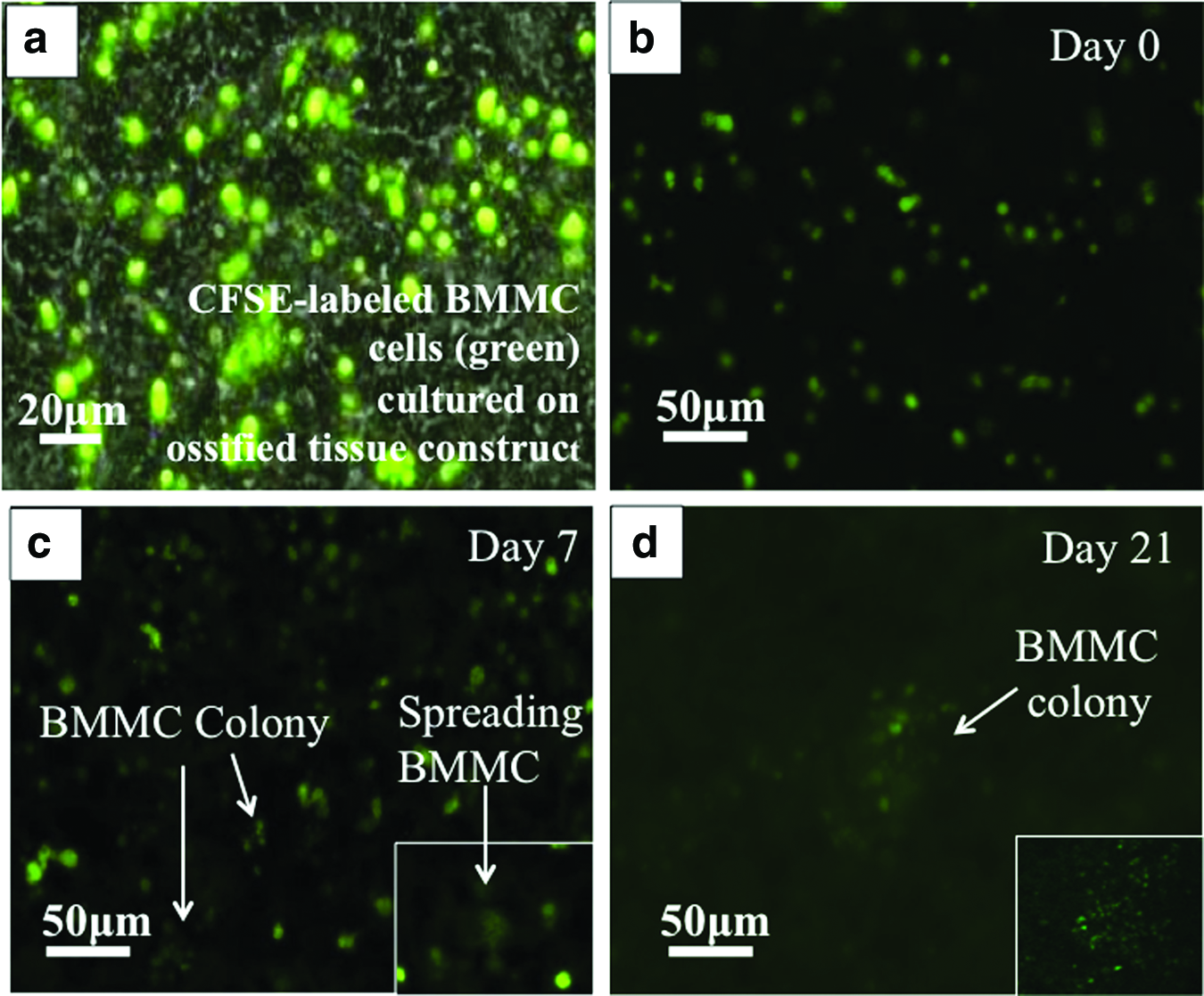
Representative imaging of CFSE-labeled BMMC biospecimens from MM patient B cultured in 3D ossified tissue.
To further evaluate potential patterns of cell–cell interactions between seeded BMMC and OSB, a smaller number of OSB (2×103 cells) were seeded in the microfluidic chamber for easier visualization and cultured for 4 days before seeding of BMMC. Analyses of real-time images showed that during the first 8 h postseeding, BMMC move toward and adhered to OSB cells (Fig. 4a). Cells that did not adhere were flushed toward the PCL barrier and began to die after 3 days in culture (data not shown).

BMMC-OSB cell–cell interactions.
In OSB cultures alone, cells proliferated and produced the ECM during the 21-day culture period. Figure 4b shows that in the absence of patient BMMC, the ossified tissue presented with more mineralization particles (black arrow). In contrast, the presence of BMMC appeared to disrupt the deposition of ECM by OSB, and hence, the more homogenous, less spotted appearance of the ossified tissue.
Quantification of multiple myeloma proliferation
To determine whether BMMC-OSB coculture could support the survival and proliferation of multiple myeloma, cells were removed from the chamber and stained as described above. The percentage of CD138+ and CD38+CD56+ MMC at the beginning of the culture and on days 7 and 21 is summarized in Table 1. Representative dot plot images of flow cytometric results can be found in Figure 5. Figure 5a shows CFSE+BMMC on days 7 and 21. Figure 5b and c shows the percentage of positively stained MMC. For all three patients studied, BMMC were retained within the ossified tissue chamber, that is, the percentage of BMMC in culture remained the same (Patient A and C) or was significantly increased (Patient B), between days 7 and 21 (Table 1). On average, BMMC accounted for 1.01% (day 7) and 1.8% (day 21) (range=0.4–3.5%) of the total cell number (BMMC+OSB) in the culture chambers. The percentage of CD138+ and CD38+CD56+ cells (within the CFSE+ population) was much higher on day 7 compared with day 0 before seeding of BMMC in the microfluidic device; however, statistical significance between these two time points was regarded as inconsequential since the BMMC within the coculture was subjected to flow conditions that likely changed the relative proportion of MMC within the plated cell population. On the other hand, comparison between consecutive culture time points was of relevance because it assessed the retention and expansion of BMMC and MMC within our system. This analysis showed that the overall percentage of MMC in culture was conserved (Patient C) or decreased between days 7 and 21. For patient A, this reduction was statistically significant in both CFSE+CD138+ and CFSE+CD38+CD56+ populations, whereas in Patient B, only the percentage of CFSE+CD138+ cells decreased significantly.
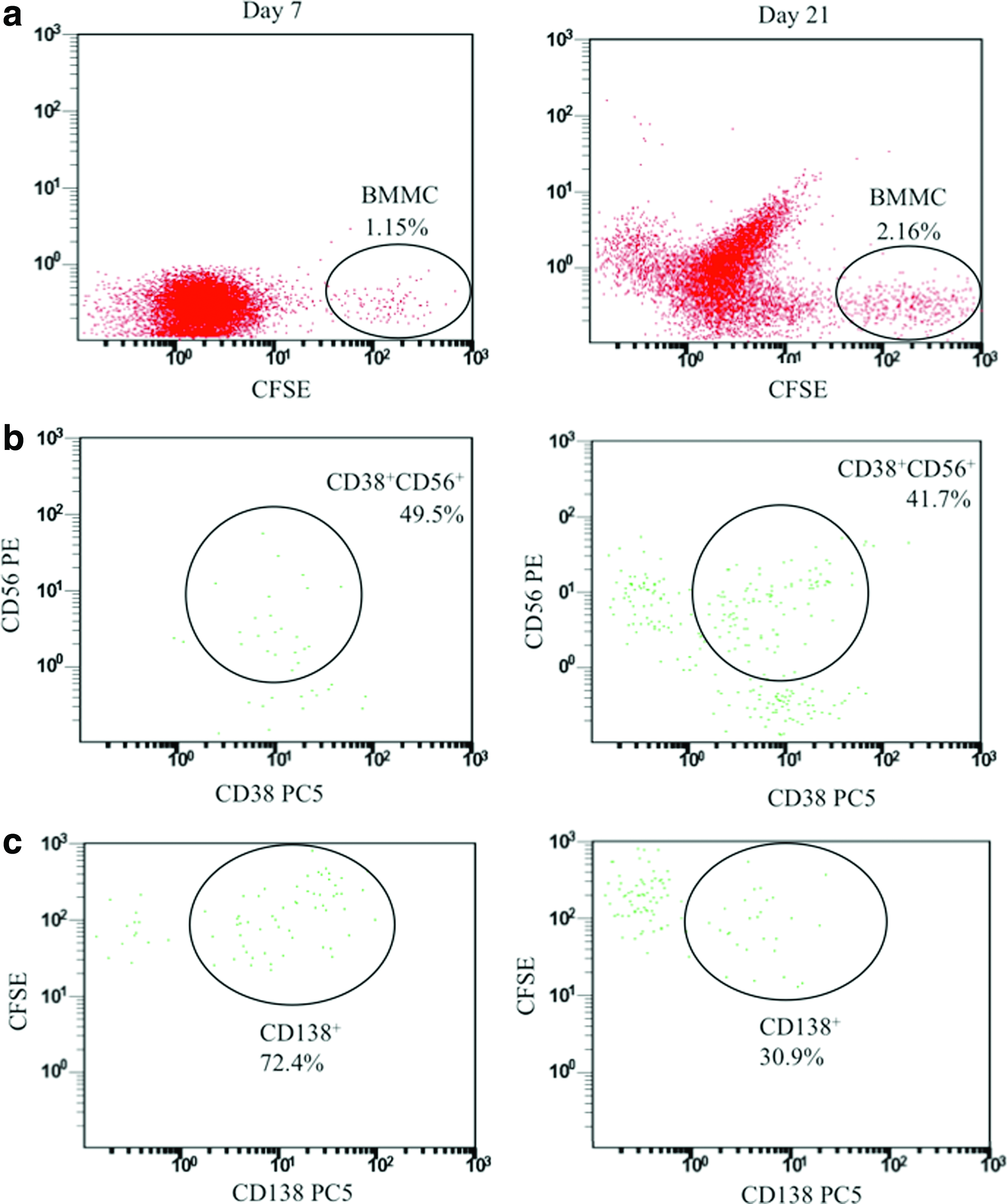
Representative dot plot images from flow cytometric analyses of patient B on days 7 and 21 of culture.
Additionally, the average expansion of BMMC and MMC for each of the three patient samples was also analyzed [Table 2, Eqn. (1)]. An example (data from one chamber) of the percentage of BMMC daughter cells under each division peak utilized to calculate the average cell expansion can be found in Table 3. For each of the three patients, BMMC underwent, on an average, 2 to 5 expansions; CD38+CD56+CFSE+ MMC underwent 1 to 3 expansions and CD138+CFSE+ MMC underwent 2.5 to 4.6 expansions. More importantly, expansion rates were maintained overtime as no statistical difference was found between day 7 and 21 cultures.
Data presented are the average result from two different chambers±SD.
Calculation example: If the percentage of cells is 20% for peak # 0, 30% for peak # 1, and 50% for peak # 2, using Equation (1), the average cell expansion=1/(0.2+0.15+0.125)=2.11.
Day 21 expansion was not statistically different to that of day 7.
Data sample from analysis of one chamber. Values were obtained from histogram plots.
Discussion
In the current study, we demonstrated that primary human MMC could be propagated in vitro by directly culturing BMMC from multiple myeloma patient biopsies in a 3D microfluidic ossified tissue constructed by a human OSB cell line. This culture system was designed to include key components of the bone/bone marrow microenvironment such as the endosteal surface (i.e., 3D OSB) and its respective secreted ECM, growth factors and cytokines from the patient's plasma, and other putative patient-derived bone marrow stromal cells, all within a microperfused environment. We found that the reconstructed endosteal layer was particularly critical for the ex vivo preservation of MMC and could properly retain these cells within the device, as other chamber designs that lacked OSB failed to do so (data not shown). In addition, the use of a premade ossified construct has an off-the-shelf added feature that allows for direct seeding of primary cells, setting this technology in line with the goals of personalized medicine therapeutics and can also facilitate the study of osteolytic and osteblastic disease, which characterizes the progression of tumors (primary and metastatic) that reside within the constrains of the bone marrow microenvironment.
Flow cytometric analysis and microscopy images showed that CD138+ and CD38+CD56+ populations were capable of proliferating in our 3D microfluidic ossified tissue system, for all three patients studied, regardless of the initial percentage of malignant cells present in the biopsy samples (Table 1). Proliferation determination was consistent with fluorescent imaging studies; for patient B, for example, the number of cells observed within a BMMC colony (20–40 cells, counted manually, Fig. 3d) was predictive of the daughter cell number generated by 5 cell divisions measured by flow cytometry (i.e., 25=32 daughter cells).
The data also suggests the existence of different proliferation rates among different cell types, that is, other BMMC might have divided faster than MMC, leading to a reduction in the overall percentage of CFSE+ cells and concomitantly, a decrease in the CFSE+ multiple myeloma populations between days 7 and 21. Although loss of CFSE labeling due to >7 rounds of proliferation could have accounted, in part, for the decrease of CFSE+MMC+ from day 7 to 21 (Table 1), only occasionally we found that the maximum number of division peaks detected was=7 (Table 3). Most likely, because of the nature of this dynamic culture system, nonadherent (hematopoietic) cells that were not trapped within the ossified tissue matrix or by the PCL barrier left the chambers during the first 24 h of culture (data not shown). Consequently, the percentage of MMC is expected to increase within the first 7 days of culture compared with the initially seeded sample, explaining the preferential retention of MMC in the ossified tissue (Table 1). Last, we also observed that most of the proliferation occurred within the first 7 days of culture, as there was no statistical difference in the cell expansion between days 7 and 21 (Table 2). Taken together, these system properties make the technology ideal for drug screening before patient treatment in the clinical setting when timely intervention is critical before disease progression takes its course.
It is well documented that the OSB activity is suppressed in multiple myeloma. 3 In our system, we also observed less ECM production by OSB cells (Fig. 4b), alluding to a functional decline in the osteoblastogenic activity. More importantly, to the best of our knowledge, this is the first time that the affinity of MMC for OSB was documented using real-time imaging (Supplementary Video S1). This behavior is consistent with previous reports showing that MMC tend to have close contact with OSB.7,12 An important consequence of these direct adhesive interactions between the bone marrow stroma and MMC is the development of drug resistance. This phenomenon termed “cell adhesion-mediated drug resistance” is thought to be one of the major mechanisms by which MMC escape the cytotoxic effects of therapeutic agents.2,13 The 3D microfluidic culture system provides a unique platform for the study of these interactions and can aid further understanding of multiple myeloma and development of novel therapeutics to abrogate microenvironment-driven drug resistance. Furthermore, the conservation of the bone marrow microenvironment and, in particular, the endosteal zone can have broader utility in the study of other blood cancers like leukemias and solid tumors such as prostate and breast cancer that also reside or metastasize to the bone marrow.
MMC are known to grow poorly outside their bone marrow milieu and attempts to optimize their maintenance have been hampered by a lack of known conditions that allow for their ex vivo survival. 14 Aside from more recently developed xenograft models,15–17 there has been only one group to date to report on creating an in vitro model capable of supporting the proliferation and survival of MMC. 7 Our unique approach, however, uses a premade bone-like 3D biological scaffold that facilitates the seeding of BMMC and is fabricated in a microfluidic device to provide a perfusion microenvironment that favors the proliferation of MMC within the first 7 days of culture. From a clinical perspective, we anticipate that the developed technology should facilitate the creation of a physiologically relevant system which (1) maximizes sample use by requiring very small amounts of patient-derived bone marrow cells (<1×106 cells, for an eight-chamber microfluidic device) and serum (<2 mL/culture/week) and (2) accelerates the evaluation of new therapeutics for the treatment of multiple myeloma.
Footnotes
Acknowledgments
This research was supported jointly by the Center for Healthcare Innovation at the Stevens Institute of Technology and the John Theurer Cancer Center at Hackensack university Medical Center.
Disclosure Statement
The authors declare no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.