
Abstract
Mesenchymal stromal cells (MSC) and factors secreted by them are essential components of the hematopoietic stem cell (HSC) niche within the bone marrow microenvironment. It has been shown that the extracellular matrix (ECM) can influence HSC-supportive potential of MSC and is a prerequisite for the proper signaling of morphogens. Therefore, we aimed at the identification of ECM components and candidate morphogens capable of enhancing the expression of HSC-supportive proteins in human MSC, namely, angiopoietin-1 (Ang-1) and stromal cell-derived factor 1 (SDF-1). For this purpose, highly sensitive secreted dual reporter constructs for Ang-1 and SDF-1 were established. These newly designed dual reporter systems enable continuous monitoring of the Ang-1 and SDF-1 promoter activity in an immortalized human MSC line cultured on ECM/morphogen microarrays. Reporter arrays showed that Ang-1 and SDF-1 expression can be induced by different ECM/morphogen combinations. In addition, continuous monitoring of promoter activity allows delineating time-dependent effects of the ECM and morphogens. Thus, we identified that collagen I and vitronectin in combination with Wnt3a favored SDF-1 expression over time, while only transiently inducing the expression of Ang-1. Taken together, the newly developed reporter systems allow for the monitoring of Ang-1 and SDF-1 promoter activity induced by morphogens and the ECM in a combinatorial and high-throughput manner. This technology might therefore be helpful to optimize culture conditions, which favor the activity of MSC as feeder cells for various types of stem and progenitor cells.
Introduction
M
ECM components do not only control mechanical properties but can also regulate the cell fate decisions through interactions with integrins, the receptors mediating the cell-ECM and cell–cell interactions. 15 Various ECM components have been shown to modulate gene and protein expression profile, proliferation, and differentiation of BM-derived MSC.16–18 Cell-free MSC-derived ECM preparations, containing several components of incorporated ECM, favor MSC proliferation, self-renewal, and multipotential differentiation potential,19–21 suggesting that the ECM secreted by MSC modulates their behavior in an autonomous manner. Besides direct regulation of cell behavior through integrins, ECM components can store growth factors and present them to the cells in a biologically active form. 22 Thus, fibronectin has been shown to enhance the biological activity of platelet-derived growth factor, 23 vascular endothelial growth factor, 24 and hepatocyte growth factor. 25 Furthermore, the activity of hepatocyte growth factor can be augmented by interactions with vitronectin. 25 These findings suggest an essential multifunctional role of the ECM in the regulation of the cellular behavior in different tissues and organs. Being the essential part of the BM microenvironment, the ECM, however, is relatively poor investigated and its role in BM remains to be elucidated.
In this study, we have addressed whether ECM components alone or in combination with morphogens from the BM microenvironment, namely, Jagged-1, Wnt3a, and N-cadherin, might regulate the HSC-supportive potential of MSC. We established reporter systems for Ang-1 and SDF-1 to identify ECM/morphogen combinations capable of inducing the expression of these HSC-supportive genes in an immortalized human MSC line (single-cell-picked clone 1 [SCP-1]). The newly designed reporter system allowed continuous monitoring of Ang-1 and SDF-1 expression in SCP-1 cultured on prepared protein microarrays. Reporter arrays identified a range of ECM/morphogen combinations capable of inducing Ang-1 and SDF-1 expression in SCP-1. Furthermore, continuous monitoring of gene expression allowed delineating time-dependent effects of ECM and morphogens.
Materials and Methods
Culturing of immortalized human BM-derived MSC
SCP-1 immortalized human BM-derived MSC overexpressing the human telomerase reverse transcriptase were kindly provided by Matthias Schieker, München, Germany. 26 SCP-1 was cultured in the Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal calf serum (FCS; Biochrom, Cambridge, United Kingdom) at 37°C and 5% CO2.
Generation of Ang-1 and SDF-1 reporter plasmids
Bacterial artificial chromosome (BAC) containing the human Ang-1 (RP11-608F23) or SDF-1 promoter (RP11-691N23) were purchased from BACPAC Resources (Oakland, CA). A cassette containing the Gaussia luciferase gene and the neomycin gene under control of the PGK promoter was inserted into each BAC by homologous recombineering directly after the transcription start site of an appropriate promoter region. Next, the fragment containing 1.4 kb of the SDF-1 promoter region and the cassette were subcloned into the XhoI/SalI cloning sites of the lentiviral transfer vector pRRL.SIN.cPPT.SFFV.GFP.WPRE (kindly provided by Christopher Baum, Hannover, Germany27,28). The pSEAP control vector was purchased from Clontech (Mountain View, CA). A fragment containing secreted alkaline phosphatase (SEAP) under the control of the SV40 promoter was amplified by polymerase chain reaction (PCR) and subcloned into each lentiviral reporter construct downstream of Gaussia luciferase and upstream of the PGK promoter to generate a dual reporter lentiviral construct. Schematics of the reporter constructs are illustrated on Figure 1A.
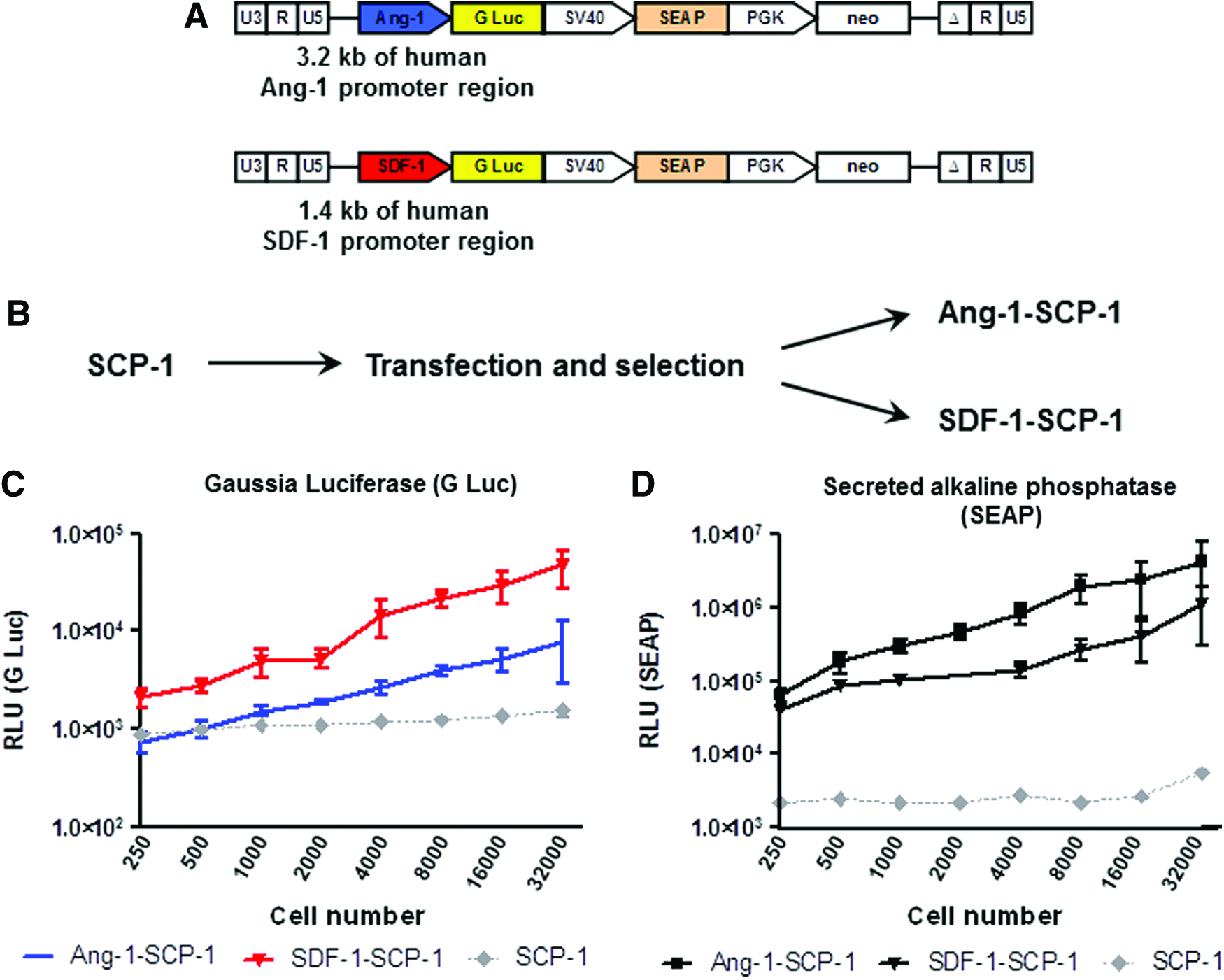
Generation of human reporter MSC.
Stable transfection of SCP-1 with reporter constructs
SCP-1 was plated at a density of 1×104/cm2 and cultured in the DMEM with 10% FCS. After 24 h, cells were transfected with 1 μg of Ang-1 or SDF-1 dual reporter lentiviral construct using Lipofectamine 2000 (Life Technologies, Carlsbad, CA) according to the manufacturer's instructions. After a 24-h transfection, the culture medium was replaced with the fresh DMEM with 10% FCS. Stably transfected cells were selected using 0.5 mg/mL G418 (Sigma-Aldrich, St. Louis, MO) for 1 week. Batch cultures were used for further applications. Stably transfected Ang-1 and SDF-1 in SCP-1 were constantly maintained in a growth medium containing 0.5 mg/mL G418.
Fabrication of ECM/morphogen microarray plates
Three hundred eighty four-well immobilizer plates were purchased from Nunc (Roskilde, Denmark). Stock solutions of human collagen I (Millipore, Billerica, MA), fibronectin (Roche, Mannheim, Germany), vitronectin (Millipore), and laminin (R&D Systems, Minneapolis, MN) were suspended at 100 μg/mL in phosphate-buffered saline (PBS). Stock solutions of human signaling proteins (R&D Systems) were suspended in PBS at the following concentrations: Jagged-1 5 μg/mL, N-cadherin 5 μg/mL, and Wnt3a 2.5 μg/mL. The BSA (Sigma-Aldrich) solution was prepared at 100 μg/mL in PBS.
A twofold serial dilution of each ECM component was prepared using 100 μg/mL of BSA solution to acquire equal total protein concentrations between all dilutions. Thus, ECM dilutions cover a concentration range from 0 to 100 μg/mL. Dilutions with 0 μg/mL of ECM protein were presented by 100 μg/mL of BSA solution only and PBS only (blank) and used as controls.
All dilutions of collagen I, fibronectin, vitronectin, and laminin were mixed with equal volumes of PBS (Ø), Jagged-1, N-cadherin, or Wnt3a solutions. The final concentration range of ECM proteins was from 0 to 50 μg/mL. Each ECM dilution used for immobilization contained, correspondingly, no signaling proteins, 2.5 μg/mL Jagged-1, 2.5 μg/mL N-cadherin, or 1.25 μg/mL Wnt3a. Each protein mixture was dispensed in triplicate into 384-well immobilizer plates at 50 μL/well. The plate layout is illustrated in Figure 2A.

Fabrication of ECM/morphogen microarray plates.
The plates were incubated with protein solutions for 2 h at room temperature with gentle agitation. Then, wells were aspirated, washed twice with PBS/0.05%Tween 20 and once with PBS. After washing, 50 μL of PBS was added to each well, and plates were subjected to electron beam sterilization with 25 kGy. Dilution of ECM, mixture of ECM and signaling proteins, dispense of defined combinations into the 384-well plates, and washing of the plates were performed using an automated Freedom EVO75 workstation (Tecan, Männedorf, Switzerland).
In experiments aimed at verification of the reporter arrays, 100 μg/mL fibronectin and 100 μg/mL vitronectin solutions containing no signaling proteins or 1.25 μg/mL Wnt3a were used for immobilization as described above. Each protein mixture was dispensed in duplicate.
Culture of reporter SCP-1 in 384-well plates
Reporter SCP-1 was plated at a density of 2×103 cells per well in 80 μL of DMEM with 10% FCS. Cells were cultured for 6–8 days, and culture supernatants were collected repeatedly every 2 days. At day 6 or 8, cells were washed once with ice-cold PBS, plates were aspirated and kept at −80°C for further total RNA extraction as described below. The supernatant collected on day 2, 4, and 6 was subjected to Gaussia luciferase and SEAP activity measurement.
Gaussia luciferase and SEAP reporter assays
The Gaussia luciferase activity was measured using the BioLux Gaussia Luciferase Assay Kit (New England Biolabs, Ipswich, MA) according to the manufacturer's instructions. The SEAP activity was measured using the NovaBright™ Phospha-Light™ EXP Assay Kit (Life Technologies) according to the manufacturer's instructions. The Gaussia luciferase activity was normalized to the SEAP activity.
Detection of immobilization proteins in 384-well immobilizer plates
Mixtures of Jagged-1 (2.5 and 5.0 μg/mL), N-cadherin (1.25 and 2.5 μg/mL), or Wnt3a (1.25 and 2.5 μg/mL) with fibronectin (100 μg/mL) were dispensed in duplicate into 384-well immobilizer plates at 50 μL/mL. After 2 h of incubation at room temperature, the plate was washed as described previously. Immobilized Jagged-1, N-cadherin, and Wnt3a were detected using rabbit anti-Jagged-1 (1:100; Cell Signaling Technology, Danvers, MA), mouse anti-N-cadherin (1:100; BioLegend, San Diego, CA), or mouse anti-Wnt3a (1:200; LifeSpan BioSciences, Seattle, WA) primary antibodies and, correspondently, anti-rabbit HRP-conjugated (1:200; GE Healthcare, Chalfont St. Giles, United Kingdom) or anti-mouse HRP-conjugated (1:200; Thermo Scientific) secondary antibodies. A substrate solution (Sigma-Aldrich) was used to develop the reaction product. The optical density was measured at 405 nm with a reference filter 570 nm using a microplate reader (Tecan).
Quantitative real-time PCR
Total RNA isolation from Ang-1 and SDF-1 reporter SCP-1 cells cultured in 384-well plates was performed using the RNeasy Micro Kit (Qiagen, Hilden, Germany). Briefly, cells were washed with PBS and lysed with the RLT buffer containing β-mercaptoethanol. The cell lysates from triplicates or duplicates were pulled and proceeded according to the manufacturer's instructions. Single-stranded cDNA was synthesized with oligo-dT primers using a RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific) according to the manufacturer's instructions. Quantitative real-time PCR was performed using a SYBR Green PCR Master Mix (Thermo Scientific) on an AB7500 Fast Real-Time PCR System (Life Technologies). The housekeeping gene GAPDH was used as a reference gene. The gene-specific primer sets are GAPDH forward 5′-GAAGGTGAAGGTCGGAGTC-3′, reverse 5′-GAAGATGGTGATGGGATTTC-3′; Ang-1 forward 5′-GAAGGGAACCGAGCCTATTC-3′, reverse 5′-GCTCTGTTTTCCTGCTGTCC-3′; SDF-1 forward 5′-AGAGCCAACGTCAAGCATCT-3′, reverse 5′-CTTTAGCTTCGGGTCAATGC-3′. All primers were synthesized by Life Technologies.
Data analysis and statistics
All results are reported as mean±SEM. Changes in the luciferase activity derived from independent experiments have been analyzed using a two-way analysis of variance with a Bonferroni multiple comparison test or two-sided paired Student's t-test, using GraphPad Prism 4.03. Statistical significance was established as *p<0.05, **p<0.01, or ***p<0.001.
Results
Generation of dual-reporter MSC to study the expression of Ang-1 and SDF-1
Ang-1 and SDF-1 dual-reporter constructs (Fig. 1A) were used for lipotransfection of SCP-1 to generate Ang-1- and SDF-1-SCP-1, respectively (Fig. 1B). To evaluate the sensitivity of the detection methods, reporter cells were plated at multiple densities. The level of Gaussia luciferase (Fig. 1C) and SEAP (Fig. 1D) derived from Ang-1- and SDF-1-SCP-1 increased in a cell number-dependent manner with a detection limit of 1000 plated cells for Ang-1-SCP-1 and 250 plated cells for SDF-1-SCP-1.
Fabrication and characterization of ECM/morphogen microarray plates
Next, we sought to screen the effects of the ECM proteins and their combinations with different morphogens on the expression of Ang-1 and SDF-1 using established reporter cells. For fabrication of ECM/morphogen microarray plates, we used commercially available 384-well plates with high protein binding affinity. Collagen I, fibronectin, vitronectin, and laminin were immobilized alone or in combination with morphogens: Jagged-1, Wnt3a, and N-cadherin. The ECM was immobilized in six different concentrations between 1.56 and 50 μg/mL, and the total protein concentration in the immobilization solution was kept constant using BSA as balance. The plate layout is illustrated in Figure 2A. Wells coated with BSA only and with a blank solution (PBS only) served as control wells. Each condition was reproduced in triplicate, and overall, 104 different conditions were available for analysis in one microarray plate.
To verify protein immobilization quantitatively and to assess whether immobilization of the ECM occurs in a gradient manner, we used TAMRA-labeled fibronectin in a concentration range between 6.25 and 100 μg/mL. The fluorescence measurement showed that the compound was immobilized in a gradient-dependent manner as intended (Fig. 2B). Immobilization of morphogens was verified using different concentrations of the morphogens in the presence of 100 μg/mL fibronectin and measured by immunochemistry (Fig. 2C).
Dose-dependent effect of ECM proteins on expression of Ang-1 and SDF-1 in SCP-1
The protein microarray plates were seeded with Ang-1- and SDF-1-SCP-1, and supernatants were subjected to Gaussia luciferase and SEAP assays at day 2, 4, and 6. Results of the reporter assays showed a pronounced dose-dependent effect of all ECM proteins (alone and in combination with morphogens) with a higher ECM concentration resulting in a higher level of Ang-1 (Supplementary Fig. S1; Supplementary Data are available online at

Expression of Ang-1 mRNA and SDF-1 mRNA confirms the results of the reporter assays.
Effect of ECM proteins and their combination with morphogens on expression of Ang-1 and SDF-1 is dynamic
Based on the initial results, we included the following conditions in repeated arrays: maximal concentrations of ECM component alone (50 μg/mL) and in combination with morphogens and morphogens alone (overall, 20 conditions were analyzed repeatedly). Results of two independent arrays are presented in Supplementary Figure S3 for Ang-1 and Figure 4 for SDF-1 (day 2, 4, and 6 of the assay).
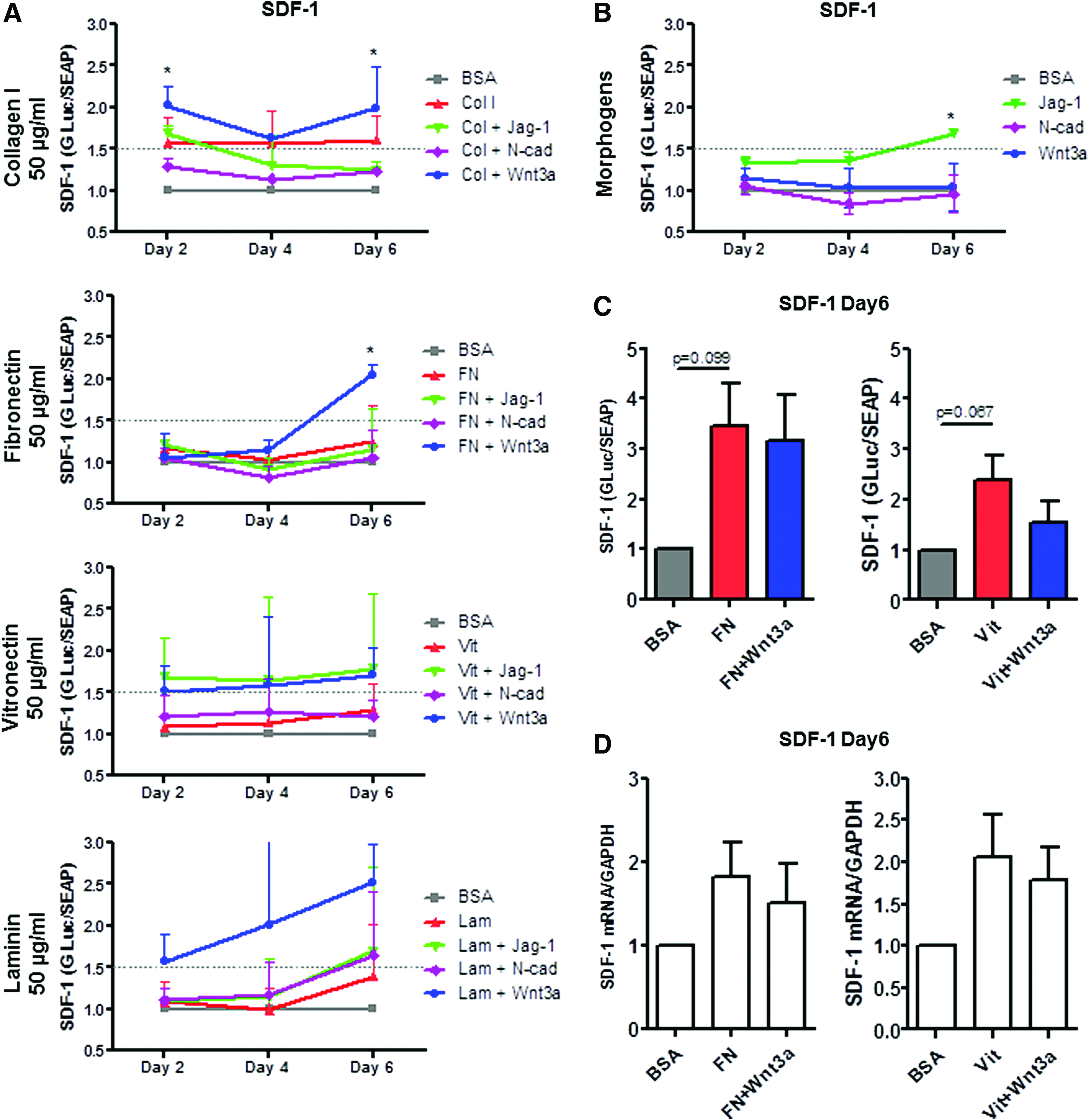
Effects of ECM and morphogens on SDF-1 expression.
Results of the arrays revealed that collagen I+Wnt3a, fibronectin+Wnt3a, and vitronectin+Wnt3a were maximally inducing Ang-1 at day 4 of the assay compared with control (collagen I+Wnt3a vs. control 1.76±0.14-fold induction, p<0.001, fibronectin+Wnt3a vs. control 1.76±0.06-fold induction, p<0.05, and vitronectin+wnt3a vs. control 1.69±0.17-fold induction, p<0.05) with no or weaker induction potential observed at day 2 and 6 (Supplementary Fig. S3A; 1.5-fold and higher induction compared with control was considered). Other conditions demonstrated no favorable effect on Ang-1 expression (Supplementary Fig. S3A).
SDF-1 expression was induced constantly over time by collagen I, collagen I+Wnt3a, vitronectin+Wnt3a, vitronectin+Jag-1, and laminin+Wnt3a (Fig. 4A), although the results, except collagen I+Wnt3a at day 2 und 6, did not reached the level of statistical significance. Furthermore, by day 6, fibnonectin+Wnt3a favored SDF-1 expression compared with control (2.1±0.12-fold induction, p<0.05, Fig. 4A and Table 1).
Effects of the highest concentrations of ECM and their combinations with morphogens on Ang-1 and SDF-1 expression at day 6. Results are presented as fold increase relatively to control (BSA only). Induction of 1.5-fold and higher is highlighted by bold. Results are provided as mean±SEM derived from two independent assays. ap<0.05.
Ang-1, angiopoietin-1; SDF-1, stromal cell-derived factor 1.
Effect of morphogens on Ang-1 and SDF-1 expression in SCP-1
Among tested morphogens, Wnt3a induced Ang-1 expression at day 2 with no induction effect at later time points (Supplementary Fig. S3B), while Jagged-1 favored SDF-1 expression at day 6 of the assay compared with control (Fig. 4B). N-cadherin affected neither Ang-1 nor SDF-1 expression.
Validation of the reporter assays
To verify the finding of the arrays, we further analyzed the effect of fibronectin (100 μg/mL) and vitronectin (100 μg/mL) alone and in combination with Wnt3a on Ang-1 and SDF-1 expression. The reporter assay revealed, as intended, no induction of Ang-1 expression (data not shown), while SDF-1 was favored by fibronectin, vitronectin, and their combinations with Wnt3a (Fig. 4C; although the difference did not reach the level of statistical significance). Analyses of Ang-1 mRNA expression also revealed no induction of Ang-1 (data not shown). The level of SDF-1 mRNA correlated with the results of the reporter assay and revealed an upregulation of SDF-1 mRNA in cells cultured with fibronectin, fibronectin+Wnt3a, vitronectin, and vitronectin+Wnt3a (Fig. 4D, although the difference did not reach the level of statistical significance).
Discussion
In the current study, we aimed at identifying the effect of ECM components and ECM/morphogen combinations on the expression of HSC-supportive genes in human MSC. The choice of the ECM (collagen I, fibronectin, vitronectin, and laminin) was based on the following rationales: (i) all ECM glycoproteins analyzed here are natural components of the bone marrow microenvironment11,12 and (ii) human MSC expressed integrin receptors to all ECM selected for the current study. 15 Morphogens (Jagged-1, Wnt3a, and N-cadherin) analyzed here are essential components of the BM microenvironment, which not only support HSC self-renewal8,29,30 but also affect MSC behavior.9,31,32 Thus, Jagged-1 through activation of Notch signaling induces the osteogenic differentiation potential of MSC, 31 while N-cadherin increases adipogenic differentiation of MSC. 32 Treatment of mouse and human MSC with Wnt3a enhances the expression of decorin and improves HSC supportive potential of MSC. 9 As the number of potential ECM/morphogen combinations can be enormous, high-throughput technologies are preferable to identify optimal combination partners, which allow mimicking the niche environment in an optimum fashion.
High-throughput ECM microarrays enable screening of cell-ECM interactions with great efficiency compared with standard approaches such as coating of typical culture plates with one of the ECM components. High-throughput approaches are also useful for screening of ECM glycoproteins and other niche signaling components in a combinatorial manner. 33 Several groups have addressed the generation of ECM/signaling protein microarrays to investigate interactions with embryonic stem cells,34,35 neural precursors, 36 or mouse HSC (LSK-CD150+ cell population). 37 Microarray techniques enable screening of cell behavior in response to a large number of protein combinations, although most often these techniques only allow endpoint analysis of cellular functions. To overcome this limitation, novel approaches incorporating sensitive and biologically relevant cellular sensors are warranted.
To monitor the effect of ECM glycoproteins and their combinations over time, we initially generated reporter constructs encoding two reporter proteins: Gaussia luciferase under the control of the human Ang-1 or SDF-1 promoters and SEAP under the control of a constitutive promoter. As Gaussia luciferase and SEAP are highly sensitive secreted reporter proteins,38,39 application of a dual reporter system allowed continuous, real-time monitoring of Ang-1 and SDF-1 gene expression in immortalized MSC in a high-throughput manner.
To screen for synergistic combinations of ECM and morphogens, we generated ECM/signaling protein microarrays. We used an automated workstation to immobilize varying concentrations of ECM components, namely, collagen I, fibronectin, laminin, and vitronectin each alone and also in combination with morphogens, in particular, Jagged-1, Wnt3a, or N-cadherin. A 384-well plate was used for each type of reporter cells; overall, more than 100 conditions could be investigated simultaneously in a 384-well plate, each in triplicate. The results of the reporter arrays allowed the identification of proteins relevant for both Ang-1 and SDF-1 expression. Combinations capable of upregulating the various HSC-supporting genes in MSC could provide superior induction of MSC feeder properties.
The reporter assay revealed a dose-dependent effect of the ECM on both Ang-1 and SDF-1 expression. In most cases, the highest concentrations resulted in the highest levels of gene expression; the only exception being, an Ang-1 induction at lower concentrations of fibronectin in combination with Wnt3a. Identification of optimal ECM concentrations may provide better control of MSC feeder potential.
The advantage of the new dual reporter culture systems, designed and developed for this project, is that they enabled dynamic monitoring of gene expression over time. They allowed identification of conditions that demonstrated long-term and short-term induction of SDF-1 and Ang-1 expression. Of the combinations screened, collagen I in combination with Wnt3a and vitronectin in combination with Wnt3a favored SDF-1 expression over time, while only transiently inducing the expression of Ang-1. Of interest, collagen alone did not favor Ang-1 expression at any time point, but induced SDF-1 expression at all time points. These results correlate with previously published data: Seib et al. studied the effect of engineered ECM components (collagen I and tropocollagen) on human MSC gene expression and observed that SDF-1, but not Ang-1 was upregulated in cells cultured on collagen I. 16 Thus, monitoring of gene expression allows the short-term and long-term effects on gene expression to be distinguished. This should be taken into consideration when developing an experimental design to achieve superior induction of the HSC-supportive potential of MSC.
With except of Wnt3a, which induces Ang-1 expression at day 2, and Jagged-1, which induces SDF-1 expression at day 6, morphogens alone do not significantly affect Ang-1 and SDF-1 expression. At the same time, reporter arrays revealed that ECM components combined with Wnt3a favored SDF-1 and to some certain extent, Ang-1 expression compared with the ECM only. These results suggest that ECM and Wnt3a can synergistically enhance expression of both Ang-1 and SDF-1 compared with ECM alone. Interestingly, a higher concentration of the ECM used for immobilization, namely, 100 μg/mL of vitronectin and 100 μg/mL fibronectin, induced SDF-1 expression to a comparable level to that observed on lower ECM protein concentrations combined with Wnt3a. These data suggest that the synergism between the ECM and Wnt3a requires an optimal ECM concentration.
In summary, the reporter systems developed by us enable investigators to systematically screen for the effects of various factors on the expression of Ang-1 and SDF-1 in MSC. The induction of these genes seems to be modulated by the ECM and morphogens in a combinatorial and time-dependent manner. Further refinements are needed to identify conditions that induce maximum support of HSC by MSC cultured under the respective conditions.
Footnotes
Acknowledgments
This work was supported by the DFG grant Collaborative Research Center 655 “Cells into tissues: Stem cell and progenitor commitment and interactions during tissue formation” (Projects B1 to K.A., B2 to M.B. and C.W., and B6 to S.B.) and by the DFG Research Group-1586 “SKELMET” (M.B. and M.W.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.