
Abstract
Objectives:
Peripheral blood mononuclear cells (PBMC) are an attractive source for the generation of osteoclasts in vitro, which is an important prerequisite for the examination of resorption and remodeling of biomaterials. In this study, different preparation methods are used to obtain cell populations with a rising content of CD14+ monocytes. We wanted to address the question whether there is a correlation between content of CD14+ cells in the preparation and functionality of formed osteoclasts.
Materials and Methods:
PBMC obtained by density gradient centrifugation with and without further purification by plastic adherence or immunomagnetic separation of CD14+ cells were seeded on both cell culture polystyrene and a calcium phosphate bone cement (CPC) and cultivated under stimulation with macrophage colony-stimulating factor (M-CSF) and receptor activator of nuclear factor-kappa B ligand (RANKL). Cell cultures were characterized by histological and fluorescent staining of multinucleated cells that were positive for tartrate-resistant acid phosphatase (TRAP) activity and the presence of actin rings, respectively. Furthermore, activities of osteoclast marker enzymes TRAP and carbonic anhydrase II (CA II) were quantified. For osteoclasts cultured on CPC, resorption pits were visualized using scanning electron microscopy (SEM).
Results:
Monocytes of all preparations were successfully differentiated into multinucleated osteoclasts showing TRAP and CA II activity on both cell culture plastic and CPC. Preparations involving an additional plastic adherence step exhibited only a minor increase of TRAP and CA II activity in the second week of cultivation. Furthermore, the number of resorption pits on CPC was reduced in these cultures compared with immunomagnetically enriched monocytes and preparations without additional plastic adherence steps. Optimal results with regard to yield, number of multinucleated osteoclasts, activity of TRAP and CA II, and resorption of CPC were obtained by simple density gradient centrifugation.
Conclusion:
All examined monocyte preparation protocols were suitable for the generation of osteoclasts on both polystyrene and CPC. Highly purified monocytes are not mandatory to obtain functional osteoclasts for investigation of biomaterial resorption.
Introduction
D
In this study, we prepare PBMC using different protocols and, accordingly, a different content of CD14+ monocytes. We analyze the potential of the monocytes to differentiate into functional osteoclasts both on cell culture polystyrene and on calcium phosphate bone cement (CPC).
Materials and Methods
Preparation of CPC samples
In this study, a CPC based on α-tricalcium phosphate as originally described by Driessens et al. 21 was used. On mixing with aqueous solution using a liquid-to-powder ratio of 500 μL/g, the cement forms a mouldable paste that sets into Ca-deficient, carbonated hydroxyapatite. Samples for in vitro experiments were prepared as disks of 10 mm diameter and 1 mm height using silicone moulds and allowed to set for 4 days in a water-saturated atmosphere at 37°C. Set cement samples had a micro-porous surface and an average surface roughness of Ra=0.48±0.12 μm as recently described. 22 Samples were sterilized by gamma-irradiation (25 kGy) and immersed in cell culture medium at 24 h before cell seeding to allow medium equilibration.
Isolation of monocytes
Human monocytes were isolated from buffy coats of healthy adult donors (German Red Cross). Each buffy coat was sub-divided into equal parts that were used for the different preparation protocols. Thus, the direct comparison between preparation protocols was not interfered in by individual variances between cells of different donors. In total, buffy coats (75–91 mL, each obtained from 500 mL blood) of nine donors were included into this study. Monocyte adhesion medium contained minimum essential medium, alpha-modification (α-MEM) containing 10% heat-inactivated fetal calf serum (FCS), 100 U/mL penicillin, 100 μg/mL streptomycin, and 2 mM glutamine (all from Biochrom). For isolation of monocytes, the the following protocols were used:
Protocol “RED” (whole PBMC fraction): The buffy coats were diluted 1:2 with phosphate-buffered saline (PBS) containing 2 mM ethylendiamintetraacetate (EDTA) and 0.5% bovine serum albumin (BSA) (both Sigma-Aldrich) (PBS E/B) and layered onto Ficoll-Paque Plus (GE Healthcare) using Leucosep tubes (Greiner). Tubes were centrifuged for 10 min at 1000 g. PBMC fraction was re-suspended in ice-cold PBS E/B and centrifuged at 250 g and 4°C for 15 min. The washing step was repeated once.
Protocol “YELLOW”: In addition to protocol “RED,” remaining erythrocytes were removed in between the washing steps. Therefore, the PBMC pellet was resuspended for 30 s in 2 mL of deionized water before adding 48 mL of ice-cold PBS (E/B) and centrifuged at 250 g for 15 min at 4°C.
Protocol “RED ADH”: After performing protocol “RED,” monocytes were enriched by plastic adherence: Cells were re-suspended in monocyte adhesion medium, transferred into serum-coated T175 cell culture flasks (coated overnight with 10 mL heat-inactivated FCS at 4°C), and incubated for 2 h at 37°C and 5% CO2. Medium was removed, and cells were washed twice with medium to remove nonattached cells. Then, cells were incubated with ice-cold PBS E/B at 4°C on a shaker to detach the adherent cells. After 30–40 min, the cell suspension was pipetted out of the flasks and centrifuged for 15 min at 250 g.
Protocol “YELLOW ADH”: After performing protocol “YELLOW,” monocytes were enriched by plastic adherence, as described earlier in the protocol.
Protocol “CD14”: After performing protocol “RED,” CD14+ cells were magnetically labeled with CD14 microbeads (Miltenyi) and separated by magnetic activated cell sorting technology according to the manufacturer`s instructions.
For all protocols, cell pellets were re-suspended in a suitable amount of PBS E/B for further analysis.
Evaluation of purity of monocyte preparations
A first estimation of monocyte content of the different preparations was obtained by measurement of cell number and size distribution with an automated cell counter (scepter 2.0.; Millipore) applying the Coulter principle. Relative frequency of monocytes was derived from the diameter histogram plot. Cells with a diameter between 7.6 and 12 μm were counted for the monocyte fraction.
Purity of monocyte preparations was further analyzed by fluorescence-activated cell sorting (FACS). One hundred thousand cells were re-suspended in 100 μL PBS, stained with monoclonal anti human CD14-APC- or FITC conjugated antibodies (Miltenyi), and fixed with 4% formaldehyde in PBS. FACS analyses were carried out using an LSR II system (BD).
Seeding and cultivation of monocytes on polystyrene and CPC
CPC discs were incubated overnight in α-MEM containing 10% heat-inactivated FCS, 100 U/mL penicillin, 100 μg/mL streptomycin, and 2 mM glutamine before seeding with cells from different PBMC preparations to ensure pH equilibration. 22 Monocyte content of the different preparations was estimated with the automated cell counter, and seeding density was adapted with regard to the monocyte content of the preparations. Forty-eight-well polystyrene cell culture plates (Nunc) were seeded with 3×105 monocytes, and CPC discs were seeded with 6×105 monocytes in 500 μL medium per sample. Samples that were seeded with preparations “RED” and “YELLOW” were incubated overnight with monocyte adhesion medium (see Isolation of Monocytes section). On the next day, the medium was changed into α-MEM containing 5% heat-inactivated FCS, 5% human A/B serum (Sigma-Aldrich), 100 U/mL penicillin, 100 μg/mL streptomycin, 2 mM glutamine, and 25 ng/mL macrophage colony-stimulating factor (M-CSF) (antibodies-online). Samples seeded with further purified cells (plastic adherence or immunomagnetic sorting) were incubated with the M-CSF-containing medium directly after seeding. After 2 days of cultivation, cells were additionally supplemented with 50 ng/mL receptor activator of nuclear factor-kappa B ligand (RANKL) (antibodies online). Medium was changed twice weekly. Samples for biochemical measurements (day 2, 9, and 16) were frozen at −80°C after washing with PBS. After 16 days, samples for microscopic investigations were fixed with 4% formaldehyde in PBS. All cell culture experiments were performed with triplicate samples and repeated thrice.
TRAP staining and light microscopy
Samples for light and fluorescence microscopy were washed with prewarmed PBS at the respective time points and fixed with 4% formaldehyde in PBS for 15 min at room temperature. Excess fixation solution was removed by washing with PBS. TRAP staining was performed for 10 min at 37°C by immersion in 0.3 mg/mL Fast Red Violet LB (Sigma-Aldrich) in an aqueous staining buffer containing 0.05 M sodium acetate (Sigma-Aldrich), 0.05 M acetic acid (Sigma-Aldrich), 0.03 M sodium tartrate (Roth), 0.1 mg/mL naphthol AS-MX phosphate disodium salt (Sigma-Aldrich), and 0.1% Triton X-100 (Sigma-Aldrich). After washing with PBS, cell nuclei were stained for 10 min using Mayers Haemalaun solution (AppliChem) followed by rinsing in tap water. Stained samples were photographed using a Zeiss Axiovert C40 microscope that was equipped with a Sony NEX-5 camera.
Fluorescence staining and confocal laser scanning microscopy
For fluorescence staining, fixed cells (see Seeding and Cultivation of Monocytes on Polystyrene and CPC section) were permeabilized by incubation in 0.2% Triton X-100 in PBS for 3 min. Then, samples were washed in PBS again and blocking was performed using 1% BSA in PBS for 45 min to prevent nonspecific binding of fluorescence dyes. Simultaneous staining of cell nuclei and actin cytoskeleton was done using 20 ng/mL 4′,6-diamidin-2-phenylindol (DAPI; Invitrogen) and 25 μL/mL AlexaFluor 488® phalloidin (Invitrogen), in 1% BSA in PBS for 45 min in the dark. After washing in PBS, stained cells were imaged using confocal laser scanning microscopy (Leica TCS SP5; Leica Microsystems), located in the “MTZ imaging” light microscopy facility of the Medical Faculty of the Technische Universität Dresden.
Scanning electron microscopy
Cells were removed from the CPC surfaces by addition of 1% Triton X-100 in PBS. To support cell lysis, the samples were sonicated for 10 min. Samples were washed with deionized water and dried at 60°C for 12 h. After drying, samples were mounted on stubs, sputter coated with carbon, imaged using a Philips XL 30/ESEM with FEG (field emission gun), and operated in scanning electron microscopy (SEM) mode at 10 mm working distance and 6 kV using an SE detector.
Determination of DNA concentration, TRAP and CA II activity
Cell lysis
Frozen cell-seeded samples were thawed for 20 min on ice followed by lysis with 1% Triton X-100 in PBS for 50 min on ice. Cell lysis was supported by sonication for 10 min in an ice-cooled ultrasonic bath. Cell lysates were transferred to test tubes and stored on ice for subsequent analysis.
DNA content
Cell lysates were mixed with Picogreen ds DNA quantitation reagent (Molecular Probes) diluted 1:800 in TE buffer (10 mM TRIS and 1 mM EDTA) and incubated for 5 min in the dark. The intensity of fluorescence was measured with a multifunction microplate reader (Infinite® M200 Pro; Tecan) at an excitation and emission wavelength of 485/535 nm. DNA content was calculated from a calibration line of calf thymus DNA (Sigma-Aldrich).
TRAP activity
According to Janckila et al., 23 TRAP reaction buffer was composed of 2.5 mM naphthol-ASBI-phosphate (Sigma-Aldrich) in 100 mM sodium acetate and 50 mM disodium tartrate pH 6.1 (both from Roth) containing 1% ethylenglycolmonomethylether and 2% Nonidet NP-40 (both from Fluka). Cell lysates (10 μL) were mixed with TRAP reaction buffer (50 μL) and incubated at 37°C for 30 min. Finally, 0.1 M NaOH (Roth) containing 0.05% Nonidet P40 (150 μL) was added to stop the enzymatic reaction. The intensity of fluorescence was measured with the multifunction microplate reader at an excitation and emission wavelength of 405/520 nm. Calibrator solutions with different TRAP concentrations (0.6–12 U/L) taken from a commercially available ELISA kit (BoneTRAP) were used to correlate the fluorescence intensity with TRAP activity. TRAP activity was related to DNA content of the respective sample.
Carbonic anhydrase II activity
Cell lysates (50 μL) were mixed with carbonic anhydrase II (CA II) reaction buffer containing 12.5 mM TRIS pH 7.5 (Roth), 75 mM NaCl (Sigma-Aldrich), and 2 mM 4-nitrophenylacetate (50 μL; Sigma-Aldrich). Absorbance was monitored at 400 nm for 5 min. Conversion to 4-nitrophenol was calculated from the slope of the absorbance plot using a calibration line from different dilutions of 1 mM 4-nitrophenol (Sigma-Aldrich). CA II activity was related to DNA content of the respective sample.
Statistics
All cell culture experiments were performed using triplicates, and the results are presented as means±standard deviation. Analysis of variance (two-way ANOVA, Origin 8.5.0G; OriginLab) for repeated measures was used to evaluate statistical significance at a level of p<0.05. Post hoc analysis was performed to determine multiple comparisons using the Tukey method (Origin 8.5.0G).
Results
Monocyte content and yield of the different preparations
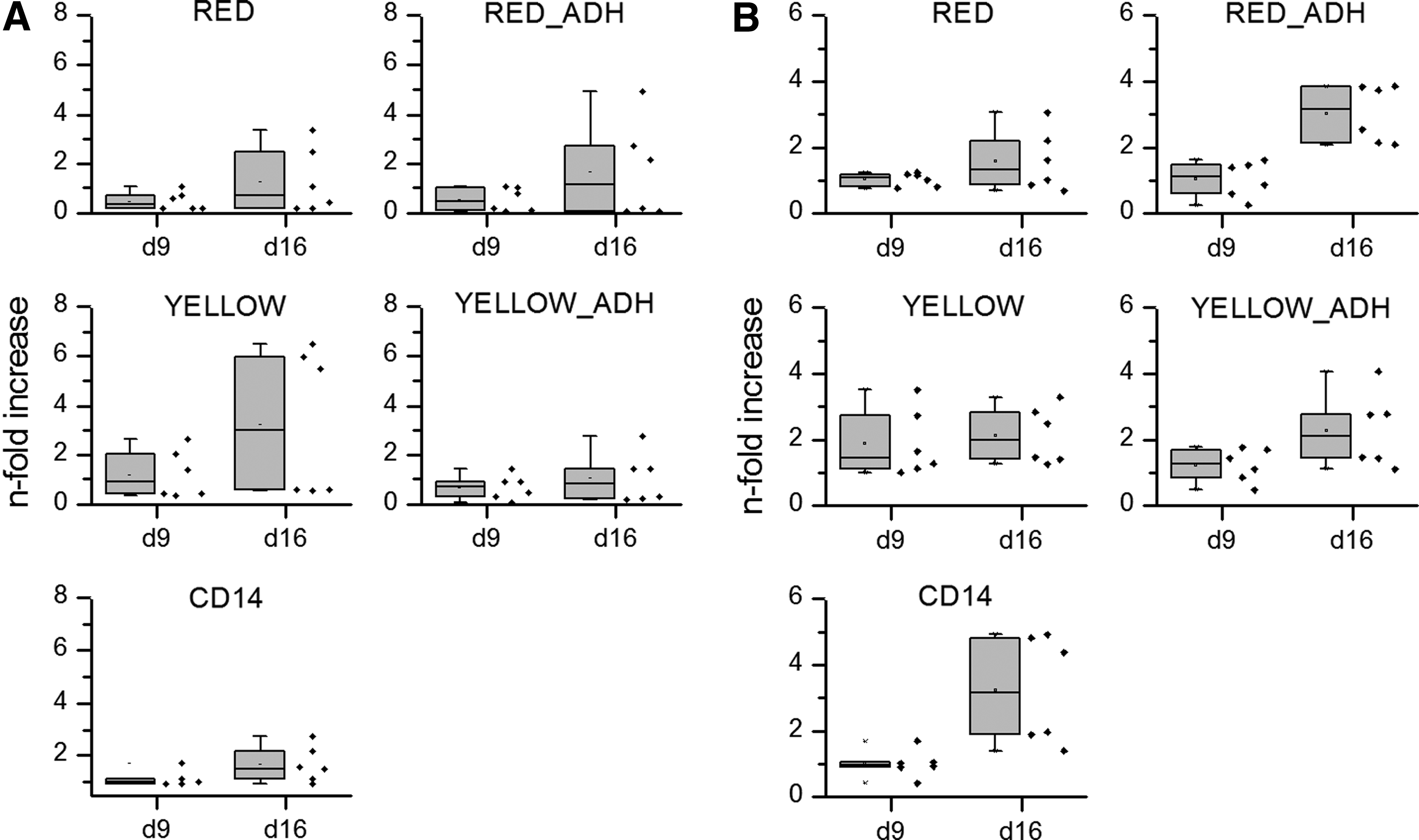
Monocyte content of the different preparations was analyzed by two different methods. We used an automated cell counter to obtain size-based histograms as a function of cell diameter as well as FACS. In size-based analysis, monocyte fraction always showed a peak with an average diameter of 9–10 μm and the monocyte content of the preparation was calculated by integrating the peak area between 7.5 and 12 μm. As expected, lowest monocyte content was obtained for PBMC fraction without further treatment (“RED”), whereas highest monocyte content was observed for magnetically purified CD14 fraction. These values were, by trend, in agreement to the content of CD14+ cells determined by FACS analysis (Table 1 and Fig. 1), although the size-based measurement revealed a somewhat higher monocyte content. Furthermore, we always detected a very low content of CD14+ cells in PBMC (“RED”), which was in contrast to already published data and the size-based measurements. Immunomagnetically enriched CD14+ monocytes (“CD14”) were not involved in FACS analysis, as the CD14 protein on the surface is blocked by the CD14 antibody-labeled magnetic beads (beads will be detached from the cells during the first days of cultivation, but not in freshly prepared cell suspensions).
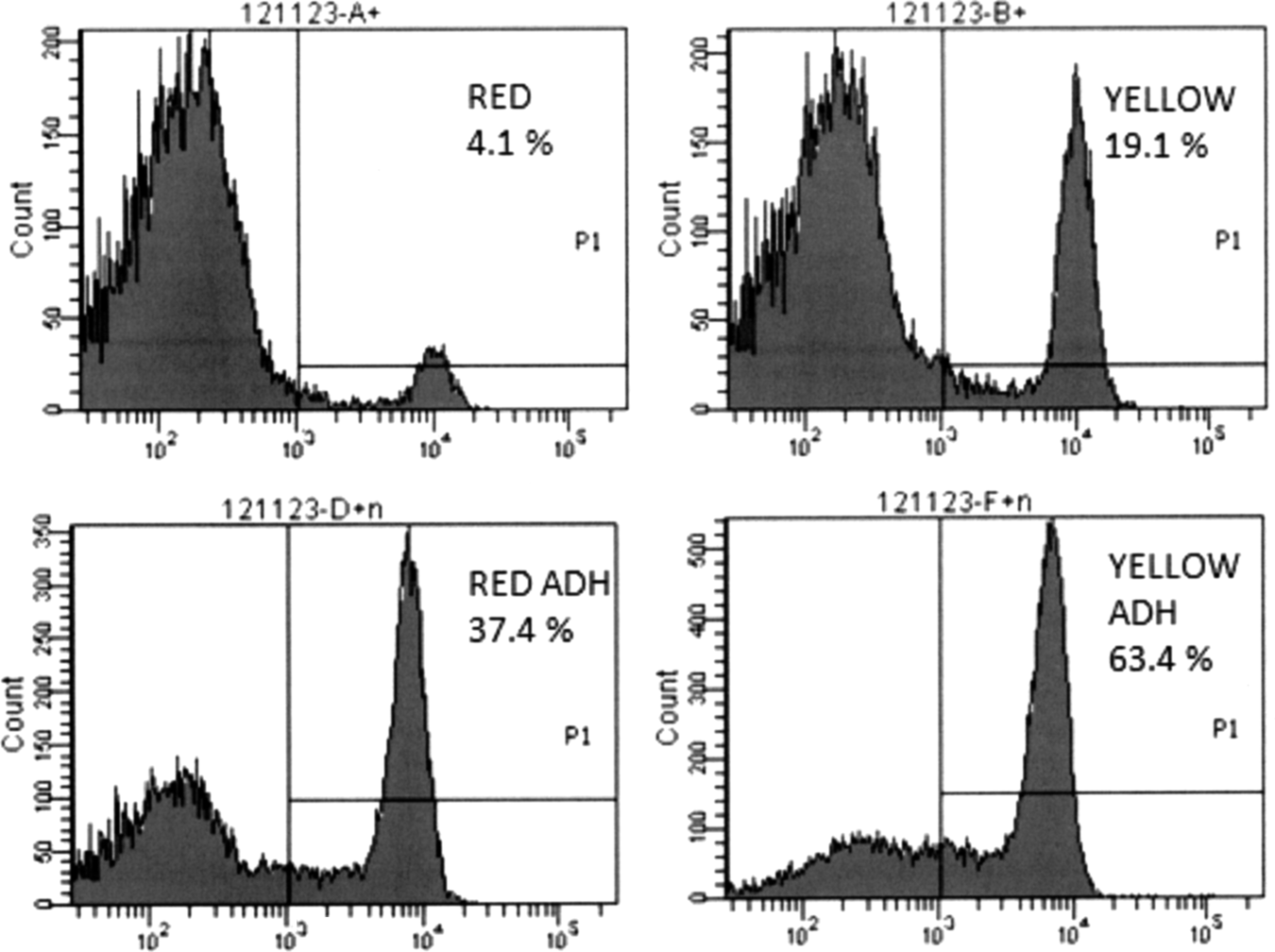
Representative flow cytometry histograms showing the purity of different monocyte preparation methods concerning positive staining for CD14.
Cells with 7.5–12 μm diameter, extrapolated to the whole buffy coat, originating from 500 mL blood.
FACS, fluorescence-activated cell sorting; PBMC, peripheral blood mononuclear cells.
Size-based histograms were also applied to calculate the overall yield of monocytic cells (defined here as cells with a diameter between 7.5 and 12 μm) from the different preparation methods (Table 1). The highest monocyte yield (up to 80×107 cells/buffy coat) was obtained after simple density gradient centrifugation (protocol “RED”). Protocols involving a step of plastic adherence (“RED ADH” and “YELLOW ADH”) gained much lower monocyte yields, as only 14–33% of the cells were recovered after the adherence step. Concomitantly, considerably higher purities were obtained compared with the PBMC (“RED”) fraction. In the case of immunomagnetically enriched CD14+ cells (“CD14” purification protocol), the highest purities combined with relatively high monocyte yields were achieved.
Osteoclast morphology on polystyrene
Multinucleated cells positively stained for TRAP activity were formed in all preparations after 9 and 16 days of cultivation with M-CSF and RANKL. As depicted in Figure 2, “CD14”-isolated cells exhibited typical osteoclast-like morphology after 9 days of culture. Large (∼100 μm) multinucleated cells with intense staining for TRAP activity were found. Similar results were obtained for cell cultures derived from monocytes after additional plastic adherence purification “(“RED ADH” and “YELLOW ADH”). Osteoclast cultures derived from monocytes applied after simple density gradient centrifugation (“RED” and “YELLOW”) also exhibited strong TRAP staining, but the osteoclast size was slightly reduced compared with the other cultures.
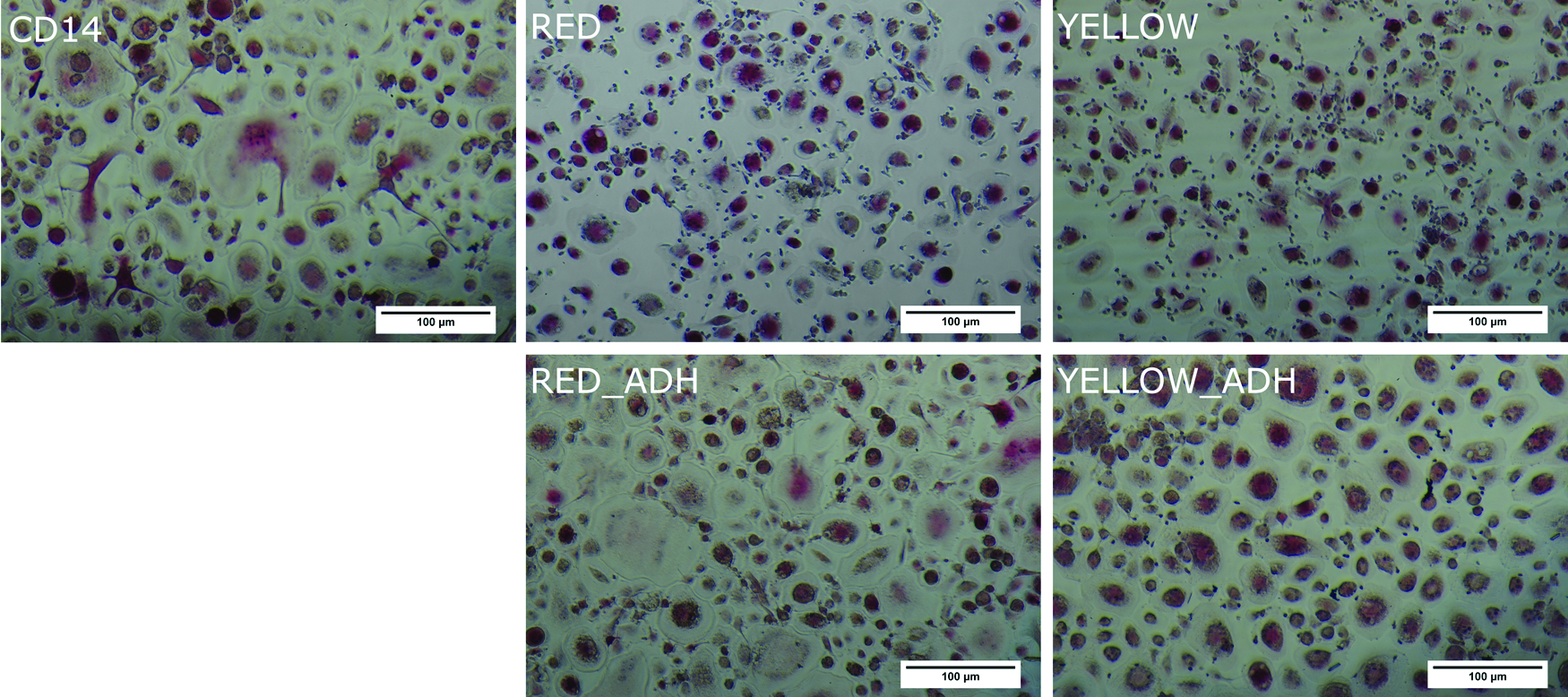
Tartrate-resistant acid phosphatase (TRAP) staining of cells cultured for 9 days under the influence of M-CSF and RANKL (haemalaun counterstaining). M-CSF, macrophage colony-stimulating factor; RANKL, receptor activator of nuclear factor-kappa B ligand. Color images available online at
In comparison to freshly prepared cells, we also used cell preparations after cryopreservation. PBMC (“RED”) barely formed multinucleated cells after thawing. Cells purified by plastic adherence as well as immunomagnetically purified cells (“RED ADH,” “YELLOW ADH,” and “CD14”) were able to form multinucleated TRAP-positive cells after thawing and cultivation with M-CSF/RANKL (data not shown).
TRAP and CA II activity on polystyrene
After 2 days of cultivation with M-CSF but before addition of RANKL, we did not detect any measureable TRAP activity in all examined samples. Likewise, CA II activity was significantly lower (p<0.05) in all examined samples after 2 days of cultivation compared with 9 and 16 days of cultivation. After 7 days of cultivation with M-CSF/RANKL (day 9 of cultivation in total), both TRAP and CA II activity were significantly increased in all samples and partly increased further until day 16 of cultivation. Figure 3 depicts a typical progress of TRAP and CA II activity in one of the experiments with immunomagnetically purified monocytes (“CD14”).
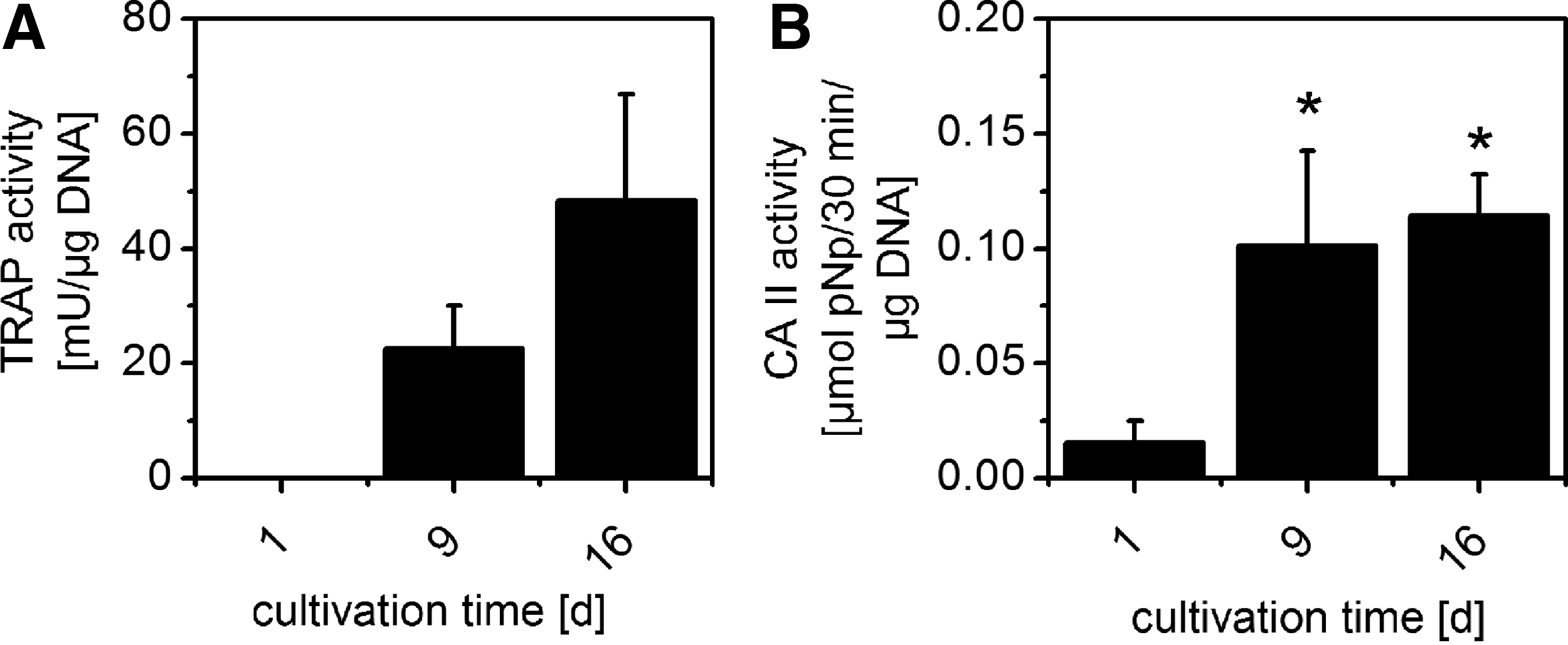
Enzyme activities of “CD14” purified monocytes under stimulation with M-CSF and RANKL on cell culture polystyrene, measured after cell lysis and 1 and 2 weeks of stimulation, respectively. RANKL was added to the samples starting from the second day of cultivation.
In each experiment, cells from the same donor were subjected to five different preparation protocols. To compare between the different independent experiments, carried out with cells from different donors, we normalized all measured TRAP and CA II activities with those of “CD14” purified cells at day 9, which was set to 1 for all experiments on polystyrene.
An increase of TRAP activity was found on day 9 and 16 in all samples; however, this was only statistically significant in cultures prepared by the protocols “CD14” and “YELLOW” (Fig. 4A). CA II activity was highest in CD14 samples at all time points. Interestingly, for CA II, a significant increase over time was, besides immunomagnetically enriched monocytes, found for cell fractions after additional plastic adherence (Fig. 4B).
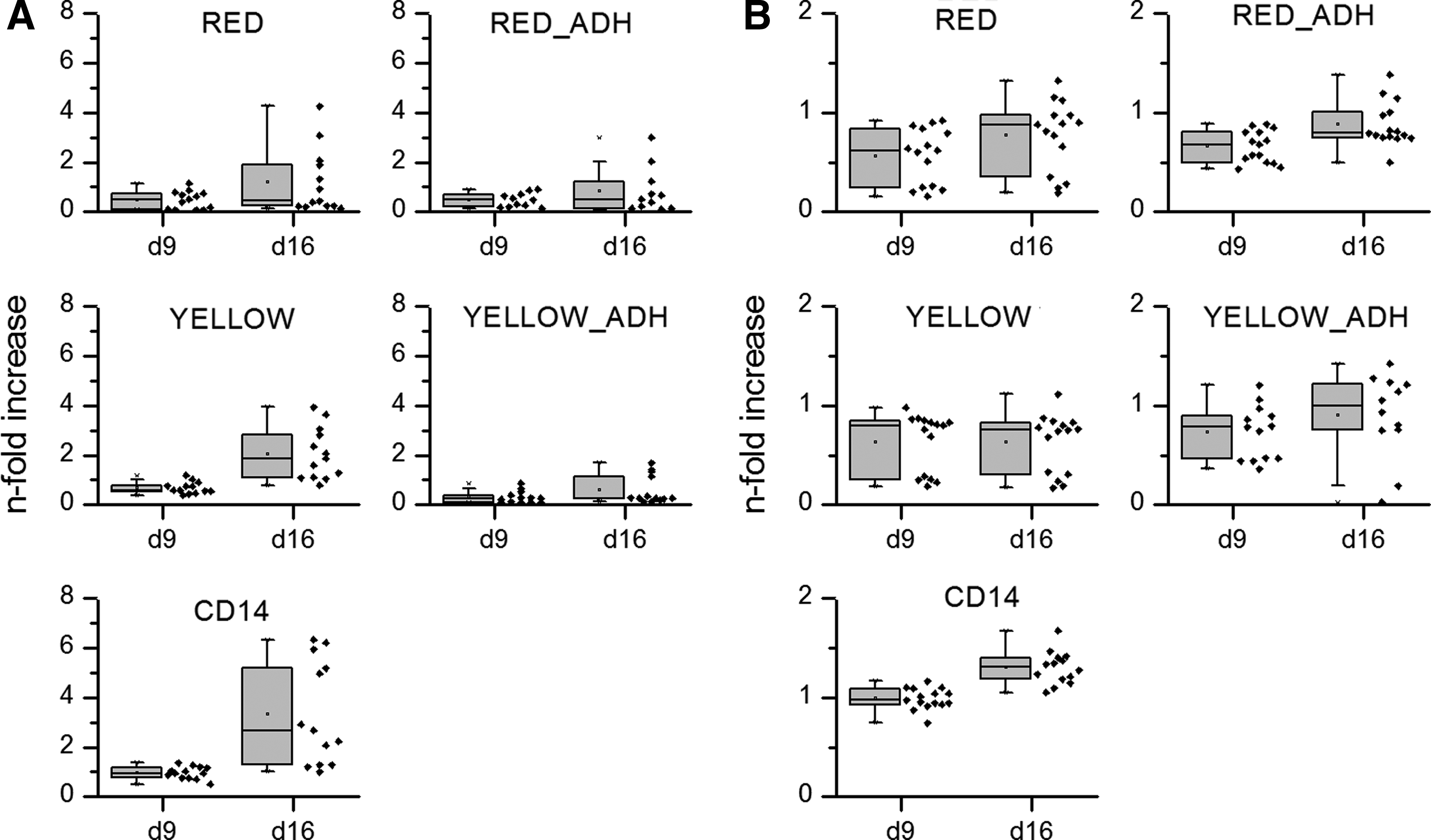
TRAP
On day 9, both TRAP and CA II activity barely differed between the respective isolation methods, while the highest activity was measured for immunomagnetically enriched cells in both cases (Fig. 5). At day 16, a significant increase in TRAP activity compared with day 9 was found in cultures derived from immunomagnetically enriched monocytes as well as from PBMC after erythrocyte lysis (“YELLOW”). Interestingly, CA II activity was significantly increased in cultures derived from immunomagnetically enriched monocytes and from cell fractions after additional plastic adherence, suggesting an inverse dependency of TRAP and CA II on the tendency of the cells to express TRAP and CA II.
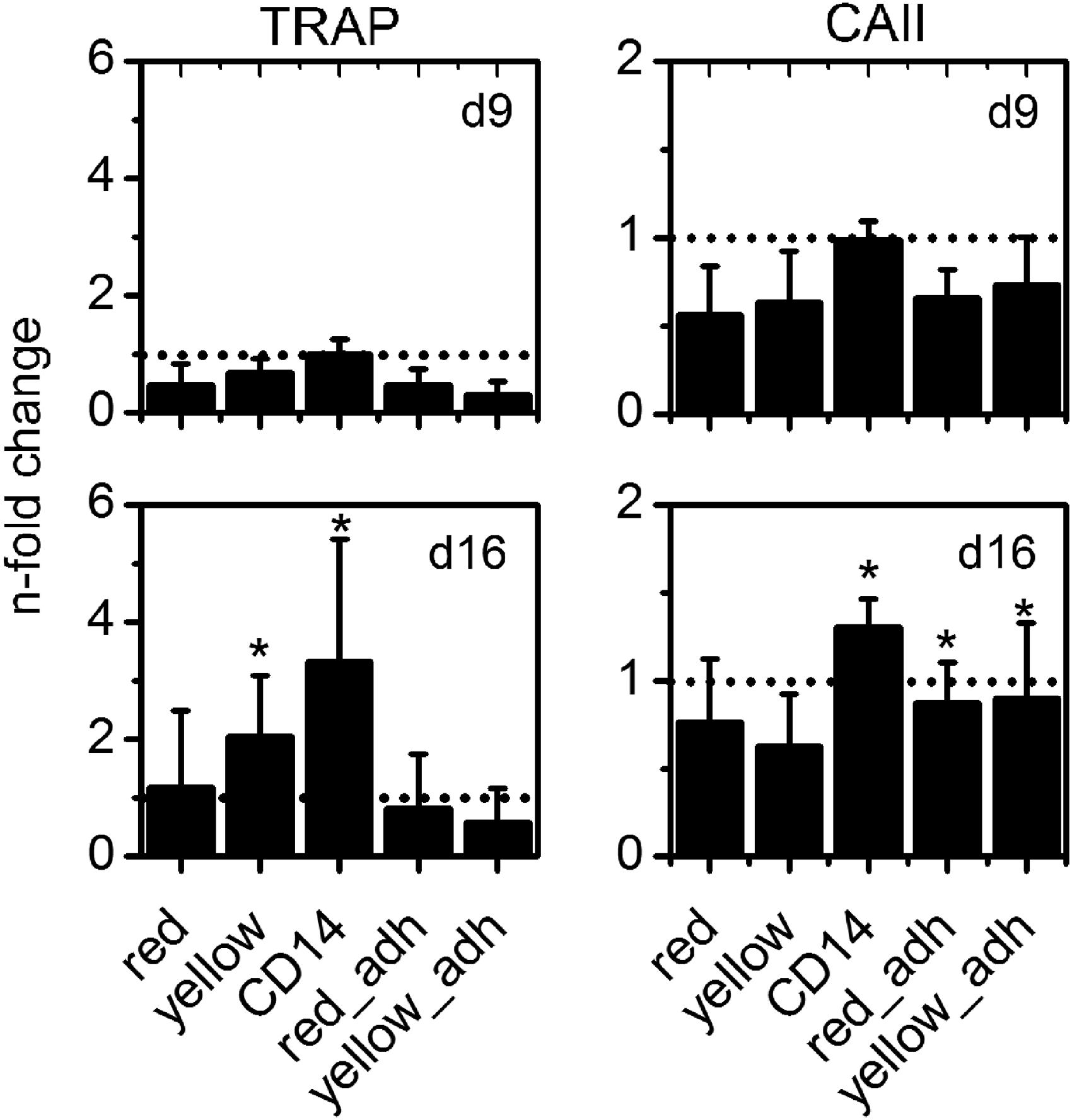
Relative TRAP and CA II activity (compared with CD14 on day 9) measured for differently isolated monocytes under stimulation with RANKL and M-CSF on polystyrene. * Indicate significant (p<0.05) changes in enzyme activity compared with those measured for CD14 samples on day 9.
Osteoclast morphology and resorption on CPC
After 2 weeks of cultivation with M-CSF/RANKL on CPC, all monocyte preparations formed multinucleated cells with actin rings, as visualized by confocal laser scanning microscopy after fluorescence staining of nuclei and actin (Fig. 6). There were no obvious differences between the different monocyte isolation methods, neither concerning size nor density of multinucleated osteoclasts.

Confocal laser scanning microscopic images of differently prepared monocytes, differentiated for 16 days under stimulation with M-CSF and RANKL on calcium phosphate bone cements (CPCs). Nuclei are stained in blue, and actin cytoskeleton is stained in green. Color images available online at
Resorption pits on the CPC were analyzed by SEM after removal of the cells (Fig. 7). The surface of the CPC shows a relatively low roughness (see Preparation of CPC Samples section), and resorption pits were easily detectable in all preparations. The highest density of pits was observed in samples, whose precursor cells were purified with simple density gradient centrifugation (protocols “RED” and “YELLOW”). Cement samples that were cultivated with osteoclasts derived from immunomagnetically enriched monocytes exhibited a lower density of resorption pits, as was also found for cement samples cultivated with osteoclasts derived from “RED ADH” purified osteoclasts. However, the size of resorption pits was larger in those samples, which is in agreement to the TRAP staining on polystyrene (Fig. 2) that also revealed osteoclasts with larger dimensions for cells derived from plastic adhesion purified and immunomagnetically enriched monocytes.

Typical scanning electron microscopy micrographs of CPC cultivated with different monocyte preparations for 16 days. Cells were removed with 1% Triton X-100 to enable the visualization of resorption pits. Scale bar=100 μm.
TRAP and CA II activity on CPC
After seeding of differently purified precursor cells on CPC and cultivation under stimulation with M-CSF and RANKL, we observed a similar progress of TRAP and CA II activity as shown for cells on cell culture polystyrene (see TRAP and CA II Activity on Polystyrene section). Both enzyme activities were not or barely detectable at 2 days after seeding of the cells on the material, followed by a strong increase of activity during the first week of cultivation (data not shown). Similar to the experiments on polystyrene (PS), we observed distinct donor variability. Therefore, all measured TRAP and CA II activities were again normalized to that of “CD14” purified cells at day 9, which was set to 1 for all CPC experiments.
In contrast to osteoclasts on polystyrene, TRAP activity on CPC did not significantly increase between day 9 and 16 of cultivation (Fig. 8). Furthermore, we observed a higher variation between samples of the same group, both for TRAP and CA II activities as well as for DNA content (DNA data not shown). Nevertheless, it can be stated that cells of all preparations showed a significant increase of TRAP and CA II activity between day 2 and 9 of cultivation.
Relative TRAP
CA II activity at day 16 of cultivation was significantly increased in cultures derived from immunomagnetically enriched monocytes as well as from PBMC after plastic adherence (RED ADH and YELLOW ADH, Fig. 9).
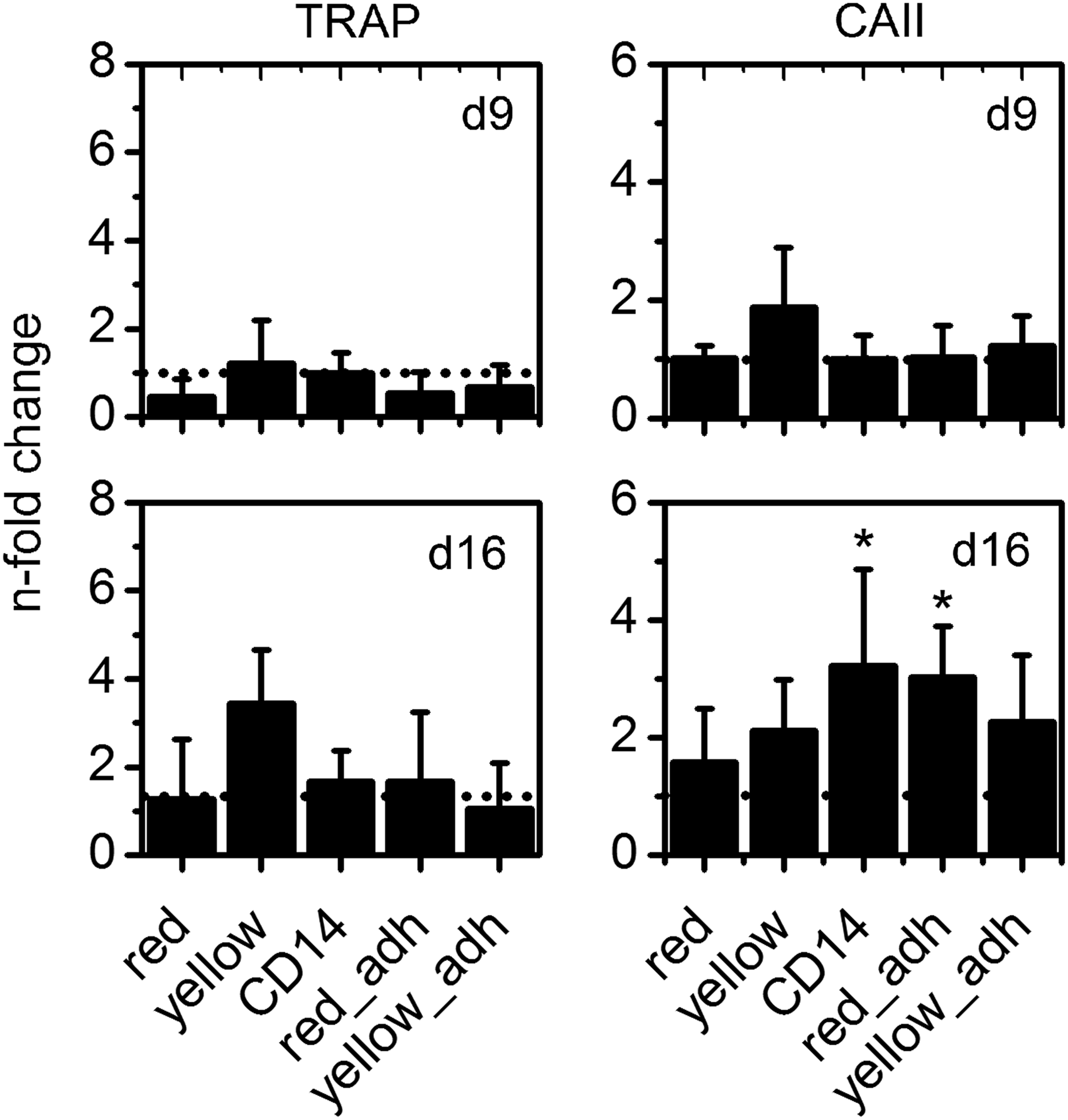
Relative TRAP and CA II activity (compared with immunomagnetically enriched cells “CD14” on day 9) measured for differently isolated monocytes under stimulation with RANKL and M-CSF on CPC. * Indicate significant (p<0.05) changes in enzyme activity compared with those measured for CD14 samples on day 9.
Discussion
The optimal monocyte density for osteoclast formation in vitro was reported to be 107 cells/mL. 24 Therefore, resorption studies with bulky biomaterials require high quantities of osteoclast precursor cells. It is well accepted that the so called classical monocytes which are strongly positive for CD14 and negative for CD16 are osteoclast precursors among the PBMC fraction of blood.12,25 An obvious implication is, therefore, that enrichment of this monocyte fraction should enable a particularly efficient osteoclastogenesis in vitro. On the contrary, nonfractionated PBMC are reported to have a high potential for in vitro osteoclastogenesis. Shalhoub et al. 13 describe the formation of resorbing osteoclasts from unsorted PBMC under stimulation with M-CSF even without additional supplementation of RANKL. Similar findings were reported a decade later by Costa-Rodrigues et al., 14 who also observed some spontaneous osteoclastogenesis in PBMC cultures in basal medium without M-CSF and RANKL. In this study, we did not analyze osteoclastogenesis without supplementation of RANKL. However, we wanted to analyze the impact of purity of different monocyte preparations and of the isolation protocols themselves on osteoclastogenesis and resorption in vitro. The preparation of pure CD14+ monocytes remains quite costly because of the application of immunomagnetic labeling techniques. Concomitantly, there is some evidence that highly purified CD14+ monocytes may perform inferior to PBMC regarding the formation of osteoclasts in some approaches. In this regard, smaller size of formed osteoclasts 19 and lower resorption activity and TRAP activity 14 were reported for osteoclasts from pure CD14+ monocyte preparations compared with PBMC. This study compares not only PBMC and CD14+ monocytes, but also other cell preparations with intermediate content of CD14+ monocytes for the formation of functional osteoclasts on a biomaterial. We wanted to address the question of whether further enriched PBMC might be an alternative to immunomagnetically purified CD14+ monocytes.
PBMC preparations obtained after density centrifugation of blood or leucocyte enriched “buffy coat” comprise monocytes, T-lymphocytes, B-cells, and minor contents of residual platelets, granulocytes, and erythrocytes. Among these cells, monocytes have the strongest potential to adhere to cell culture polystyrene, glass, and other surfaces. Nevertheless, other PBMC may also adhere weakly, resulting in a population of contaminating lymphocytes at least during the early stages of cultivation. 18 T-lymphocytes express soluble RANKL, 26 which might be an explanation for the spontaneous osteoclastogenesis in PBMC observed in the studies mentioned earlier. 27
As expected, different preparation methods attained cell populations with different monocyte contents. We applied two methods for evaluation of the monocyte content: size-based evaluation utilizing the Coulter principle and FACS analysis for the CD14+ cell population. The percentage of CD14+ cells matched quite well the percentage of cells in the size range of 7.5–12 μm diameter. By tendency, the size-based measurement obtained slightly higher values, indicating that not all cells with a diameter above 7.5 μm are CD14+. The only exception was the small percentage of CD14+ cells in the PBMC fraction, which is in contrast to size-based measurements as well as to published data. We hypothesize on hampered antibody binding caused by possible CD14 ligands present in the PBMC but not in the other monocyte preparations. Nevertheless, the size-based measurement seems to be a good alternative for FACS, as it is fast (<1 min) and saves expenses for fluorescence-labeled antibodies. It is of enormous importance to know the monocyte content of a cell preparation before seeding to a material. Different authors have stated that sufficient density of precursor cells is a crucial requirement for osteoclastogenesis.12,24 Consequently, when seeding cell preparations with low monocyte content, a higher initial cell density is required. Accordingly, Nicholson et al. 12 reported the necessity to seed the 10-fold amount of PBMC compared with immunomagnetically enriched CD14+ monocytes on bone slices to get comparable amounts of TRAP-positive osteoclasts. Since monocyte content in the different preparations is highly variable (typically between 8% and 90% due to donor variability and preparation method), we highly recommend the determination of monocyte content and further adaptation of seeding cell number.
An important hallmark for the evaluation of different monocyte isolation protocols is the monocyte recovery per mL blood or buffy coat. In fact, the number of monocytes at first depends on the monocyte content of the respective blood sample. Nevertheless, also the preparation method strongly influences the total number of monocytes obtained. The highest monocyte yields were always obtained for the two PBMC fractions (“RED” and “YELLOW”), which is reasonable, as only a few preparation steps are performed in these protocols. The obtained cell numbers from our PBMC preparations (estimated from size-based histograms) are in a similar range of already published data.16,17,24 On the contrary, an additional step of plastic adherence considerably decreased monocyte yield to ∼10% compared with the PBMC samples. Zhou et al. 28 also compared different monocyte purification protocols with regard to purity, yield, and functionality in a phagocytosis assay. For monocytes isolated by plastic adherence compared with immunomagnetically selected CD14+ cells, the authors stated lower purity and yield as well as higher variances due to difficulties in standardizing the washing steps. Monocytes after simple density gradient centrifugation were not considered in their study. One advantage of monocyte preparations after additional plastic adherence is a better performance in cryo-conservation compared with PBMC. Monocytes, which were purified by plastic adherence, can be easily cryo-conserved and functional osteoclasts are obtained after thawing and cultivation. In our experiments, cryo-conserved PBMC showed considerably lower numbers of osteoclasts after thawing, removal of nonadherent cells, and cultivation with M-CSF and RANKL compared with fresh PBMC (data not shown). Apparently, a higher purity of the cell population supports recovery after freezing.
The main focus of this work was the suitability of cell populations with different content of monocytes for osteoclastogenesis in vitro, especially on CPC. Only a few publications report on the osteoclastic differentiation of human monocytes on CPC.24,29–31 In this study, we applied a well-established α-TCP-based CPC to analyze the formation and functionality of osteoclasts with special regard to the preparation of monocyte precursor cells. All preparations gave rise to multinucleated osteoclasts with TRAP and CA II activity, on both cell culture polystyrene and CPC. It has been shown that activity of TRAP correlates significantly with the number of osteoclasts, even in the absence of resorbable material. 32 Thus, the measurement of TRAP activity enables the comparison of osteoclast number between different preparation protocols. Another characteristic marker of mature osteoclasts is CA II,33,34 being responsible for extracellular acidification, which is a prerequisite for dissolution of bone mineral. While many studies focus on expression analysis of the CA II gene, its enzyme activity is also regarded an osteoclast marker. 35 In contrast to TRAP activity, there seems to be no distinct correlation between CA II activity and osteoclast number. We detected a significant increase of CA II activity in all examined samples between day 2 and 9 of cultivation. However, during the second week of cultivation, elevated TRAP activity was not necessarily accompanied by elevated CA II activity and vice versa. Nevertheless, CA II activity might be a useful indicator for the formation of osteoclasts, as it is barely detectable in PBMC and monocytes. Further studies are necessary to clarify the correlation between the enzyme activities of TRAP and CA II with osteoclast number and resorption on different biomaterials. Variations in TRAP and CA II activity between cells of different donors were higher than variations between different purification protocols, indicating that all examined protocols for the enrichment of monocytes are suitable for osteoclastogenesis on both cell culture plastic and CPC. A direct comparison of osteoclast formation on polystyrene and CPC was not the main focus of this study. We rather intended to analyze the osteoclast-forming potential of the different monocyte preparations at first on a well-defined, easily observable, and well-investigated surface. Since cell culture polystyrene is neither resorbable nor suitable for implantation, we extended our investigation on a resorbable bone graft material. Nevertheless, we want to briefly discuss one observation from the comparison of polystyrene and CPC. While osteoclasts on polystyrene showed a further increase of TRAP activity between day 9 and 16 of cultivation, TRAP activity of osteoclasts on CPC was not significantly increased within this period. Assuming that absolute TRAP activity in a lysate sample correlates to osteoclast cell number, 32 these findings would indicate a higher osteoclast number on polystyrene compared with CPC after 16 days of cultivation, which is contradicting to our microscopic analyses on both substrates. On the other hand, resorbing and nonresorbing osteoclasts differ in organization of the actin cytoskeleton 36 as well as in gene expression of osteoclast markers cathepsin K and RANK. 37 Possibly, TRAP activity is also influenced by the substrate and may not in all cases reflect the osteoclast number or even resorption. Therefore, further investigations on resorbable and nonresorbable substrates are necessary to clarify the role of TRAP activity as a resorption marker. Resorption of CPC was demonstrated by visualization of resorption pits for all preparations. However, the monocytes that were purified through an additional plastic adherence step generated fewer resorption pits on the calcium phosphate material. After all examinations, we do not favor purification by plastic adherence for different reasons: besides the possibly reduced resorption activity of the obtained osteoclasts, the recovery of monocytes after plastic adherence is quite poor. Even though we succeeded in increasing monocyte recovery by coating cell culture flasks with FCS, which also reduced the amount of damaged cells as scraping was not necessary to detach the cells, recovered cells tend to agglomerate. Cell agglomerates complicate uniform distribution of the monocytes on the cements, which might be unfavorable for osteoclast formation. Direct adherence of monocytes directly after isolation of PBMC (“RED”) or erythrocyte-depleted PBMC (“YELLOW”) to the biomaterial followed by removal of nonadherent cells with the next medium change is regarded as the preferred method for the generation of osteoclasts on biomaterials. Highly purified monocytes (“CD14”) enriched by immunomagnetic selection do not provide better results with regard to osteoclast formation and functionality, at least for cell culture on polystyrene and CPC.
Conclusion
The five examined purification protocols for human monocytes from leukocyte-enriched blood samples were suitable for the generation of osteoclasts on both cell culture polystyrene and CPC. Optimal results concerning yield, number of multinucleated osteoclasts, activity of TRAP and CA II, and resorption of CPC were obtained by simple density gradient centrifugation.
Footnotes
Acknowledgments
The authors wish to thank Hella Luksch (Pediatric Clinic of the University Hospital Dresden) and Angela Jacobi for FACS measurements. They are grateful to InnoTERE GmbH (Radebeul, Germany) for providing CPC precursor powder and acknowledge the technical assistance of Sophie Brueggemeier, Anna Rumiantseva, and Kristin Faulwasser and the financial support rendered by DFG (SFB/TR79).
Disclosure statement
No competing financial interests exist.