
Abstract
Cardiac tissue engineering is a promising method for regenerative medicine. Although we have developed human cardiac cell sheets by integration of cell sheet-based tissue engineering and scalable bioreactor culture, the risk of contamination by induced pluripotent stem (iPS) cells in cardiac cell sheets remains unresolved. In the present study, we established a novel culture method to fabricate human cardiac cell sheets with a decreased risk of iPS cell contamination while maintaining viabilities of iPS cell-derived cells, including cardiomyocytes and fibroblasts, using a methionine-free culture condition. When cultured in the methionine-free condition, human iPS cells did not survive without feeder cells and could not proliferate or form colonies on feeder cells or in coculture with cells for cardiac cell sheet fabrication. When iPS cell-derived cells after the cardiac differentiation were transiently cultured in the methionine-free condition, gene expression of OCT3/4 and NANOG was downregulated significantly compared with that in the standard culture condition. Furthermore, in fabricated cardiac cell sheets, spontaneous and synchronous beating was observed in the whole area while maintaining or upregulating the expression of various cardiac and extracellular matrix genes. These findings suggest that human iPS cells are methionine dependent and a methionine-free culture condition for cardiac cell sheet fabrication might reduce the risk of iPS cell contamination.
Introduction
R
It is well known that the growth of many types of tumors is methionine dependent, 18 one of the essential amino acids, and the molecular imaging of the uptake of methionine has been already used for the diagnosis and staging of some tumors in the brain, lung, and mammary gland, and sarcoma and lymphoma. 19 On the other hand, various types of somatic cells, including human fibroblasts, have been reported to proliferate in a methionine-free condition. 20 Pluripotent stem cells have an unlimited growth capacity and are able to generate benign teratomas or teratocarcinomas upon transplantation. 21 Although the mechanisms of the methionine dependency of cancer cells have not been clarified fully, we hypothesized that the growth of highly replicative cells such as iPS cells might be methionine dependent, whereas that of weakly replicative cells such as cardiomyocytes and fibroblasts might be methionine independent.
The aims of this study were to elucidate the effects of a methionine-free culture condition on iPS cell growth and the viabilities of iPS-derived somatic cells, and to establish human cardiac cell sheets with a low risk of contamination by the remaining iPS cells.
Materials and Methods
Human iPS cell culture
Human iPS cell lines (253G1 and 201B7) were purchased from RIKEN (Tsukuba, Japan) and maintained in Primate ES Cell Medium (ReproCELL, Yokohama, Japan) supplemented with 5 ng/mL basic fibroblast growth factor (bFGF; ReproCELL) on mitomycin C-treated mouse embryonic fibroblasts (MEFs; ReproCELL). Cells were passaged as small clumps every 3–4 days using CTK solution (ReproCELL). For feeder-less culture experiments, iPS cells were adapted and maintained on Matrigel (growth factor reduced, 1:60 dilution; Life Technologies, Carlsbad, CA) in mTeSR-1 (STEMCELL Technologies, Vancouver, Canada). In some experiments, iPS cells were stained with Cell Tracker Orange (Life Technologies) according to the manufacturer's instructions. Images of stained samples were obtained by a fluorescence microscope (Nikon, Tokyo, Japan) with NIS-Elements software (Nikon).
Cardiac differentiation in the bioreactor and cardiac cell sheet preparation
The cardiac differentiation protocol has been described previously 12 and was performed with some modifications. Briefly, at 2 days after starting the culture in the bioreactor system with mTeSR1, EBs were cultured in StemPro34 medium containing 50 μg/mL ascorbic acid (Sigma-Aldrich, St. Louis, MO), 2 mM L-glutamine (Life Technologies), and 400 μM 1-thioglycerol (Sigma-Aldrich). The cells were then treated with 0.5 ng/mL bone morphogenic protein-4 (R&D systems, Minneapolis, MN) (days 2–3), 10 ng/mL bone morphogenic protein-4 (days 3–6), 5 ng/mL bFGF (days 3–6), 3 ng/mL activin A (R&D Systems) (days 3–6), 4 μM IWR-1 (Wako, Osaka, Japan) (days 6–8), 5 ng/mL vascular endothelial growth factor (R&D Systems) (days 8–18), and 10 ng/mL bFGF (days 8–18). Before seeding the cells, the surfaces of temperature-responsive dishes (UpCell; CellSeed, Tokyo, Japan) were coated with FBS for 2 h. After cardiac differentiation, the cells were dissociated with 0.05% trypsin/EDTA, cell aggregates were removed using a strainer (BD Biosciences, San Jose, CA), and single cells were plated onto the UpCell at 2.1×105 cells/cm2 in DMEM supplemented with 10% FBS at 37°C in a humidified atmosphere with 5% CO2.
Immunocytochemistry
Cells were fixed with 4% paraformaldehyde, and the immunostaining methods have been described previously. 22 Samples were imaged by Image Express (Molecular Device, Sunnyvale, CA) with MetaXpress and AcuityXpress software (Molecular Device). The following antibodies were used: mouse monoclonal anti-cardiac troponin T (Thermo Scientific, Rockford, IL), rabbit polyclonal anti-SM22 (Abcam, Cambridge, United Kingdom), and goat polyclonal anti-Oct3/4 (R&D systems). Secondary antibodies were purchased from Jackson ImmunoResearch Laboratories (West Grove, PA).
RNA extraction and quantitative reverse transcriptase-polymerase chain reaction
Total RNA extraction and reverse transcriptase-polymerase chain reaction (RT-PCR) were performed as described previously. 23 Primer pairs and TaqMan MGB probes were designed for human OCT3/4, NANOG, MYL2, MYL7, TNNT2, COL1A1, COL3A1, NPPA, NPPB, ACTB, and GAPDH with the TaqMan gene expression assay (Table 1) (Applied Biosystems [ABI], Foster City, CA). Quantitative PCR was performed with a 7300 Real-Time PCR System (Applied Biosystems). The relative mRNA expression level was calculated using a standard curve of ACTB or GAPDH mRNA levels.
ABI, Applied Biosystems.
Statistical analyses
Data are presented as the mean±standard deviation. Statistical analyses were performed with Student's t-test for comparison between two groups. Multiple group comparisons were performed by one-way analysis of variance followed by Tukey's or Dunnett's procedure for comparison of means. A value of p<0.05 was considered statistically significant.
Results
Methionine is critical for the survival and growth of human iPS cells
First, we examined whether iPS cells could be cultured using a methionine-free medium. At 2 days after starting the culture with mTeSR1 on Matrigel-coated plates, iPS cells (253G1) were assigned to the following conditions: mTeSR1, RPMI1640+B27 supplement [RPMI1640 with methionine, R(+)], or RPMI1640 (methionine-free)+B27 supplement [RPMI1640 without methionine, R(−)] (Fig. 1A). The next day, human iPS cells formed colonies in not only mTeSR1 but also R(+) (Fig. 1C). On the other hand, almost all iPS colonies had disappeared when iPS cells were cultured with R(−) (Fig. 1C). These findings suggest that methionine might be critical for iPS cell survival. Next, we examined whether methionine was also necessary for the growth and survival of iPS cells cultured on feeder cells. At 2 days after starting the culture on MEFs with primate ES medium containing bFGF, iPS cells (253G1) were assigned to the following conditions: primate ES medium with bFGF, R(+), or R(−) (Fig. 1B). Many iPS cell colonies were observed in the preparation with primate ES medium with bFGF and R(+), whereas a low number of small iPS colonies were observed when iPS cells were cultured with R(−) (Fig. 1C). These phenomena were also observed for the other iPS cell line (201B7) (Fig. 1C). These findings suggest that methionine might be important for the growth and survival of iPS cells in feeder and feeder-less conditions.

Methionine is indispensable for human iPS cell growth and survival.
Methionine is necessary for human iPS cell growth in cardiac cell sheet preparation
Human cardiac cell sheets were fabricated by culturing human iPS cell-derived cells after the cardiac differentiation in DMEM supplemented with 10% FBS. Because contamination by the remaining iPS cells increases the risk of tumor formation after transplantation, we examined the growth of human iPS cells in coculture with human iPS cell-derived cells after the cardiac differentiation. At 1 day after starting the culture of human iPS cell-derived cells, small aggregates of human iPS cells were seeded and cultured in primate ES medium with bFGF for 1 day. Then, the cells were assigned to the following conditions: primate ES medium with bFGF, 10% FBS DMEM, R(+), or R(−) (Fig. 2A). As shown in Figure 2B, large iPS cell colonies were observed when cultured in not only primate ES medium with bFGF but also 10% FBS DMEM and R(+). These findings indicate the possibility that the remaining iPS cells are able to grow in the bioengineered cardiac tissue even under unconventional culture conditions for human iPS cells. On the other hand, when iPS cells were cocultured with human iPS cell-derived cells in R(−), we observed very small colonies at day 1 (Fig. 2B). Furthermore, the number of Oct3/4-positive cells were significantly decreased in R(−) condition compared with other methionine-containing conditions (Fig. 2C) at day 1. Moreover, the number of Oct3/4-positive cells was decreased in R(−) condition in a time-dependent manner until day 2 (Fig. 2D). These findings suggest that a methionine-free condition might prevent the growth of human iPS cells in cardiac cell sheet preparation.
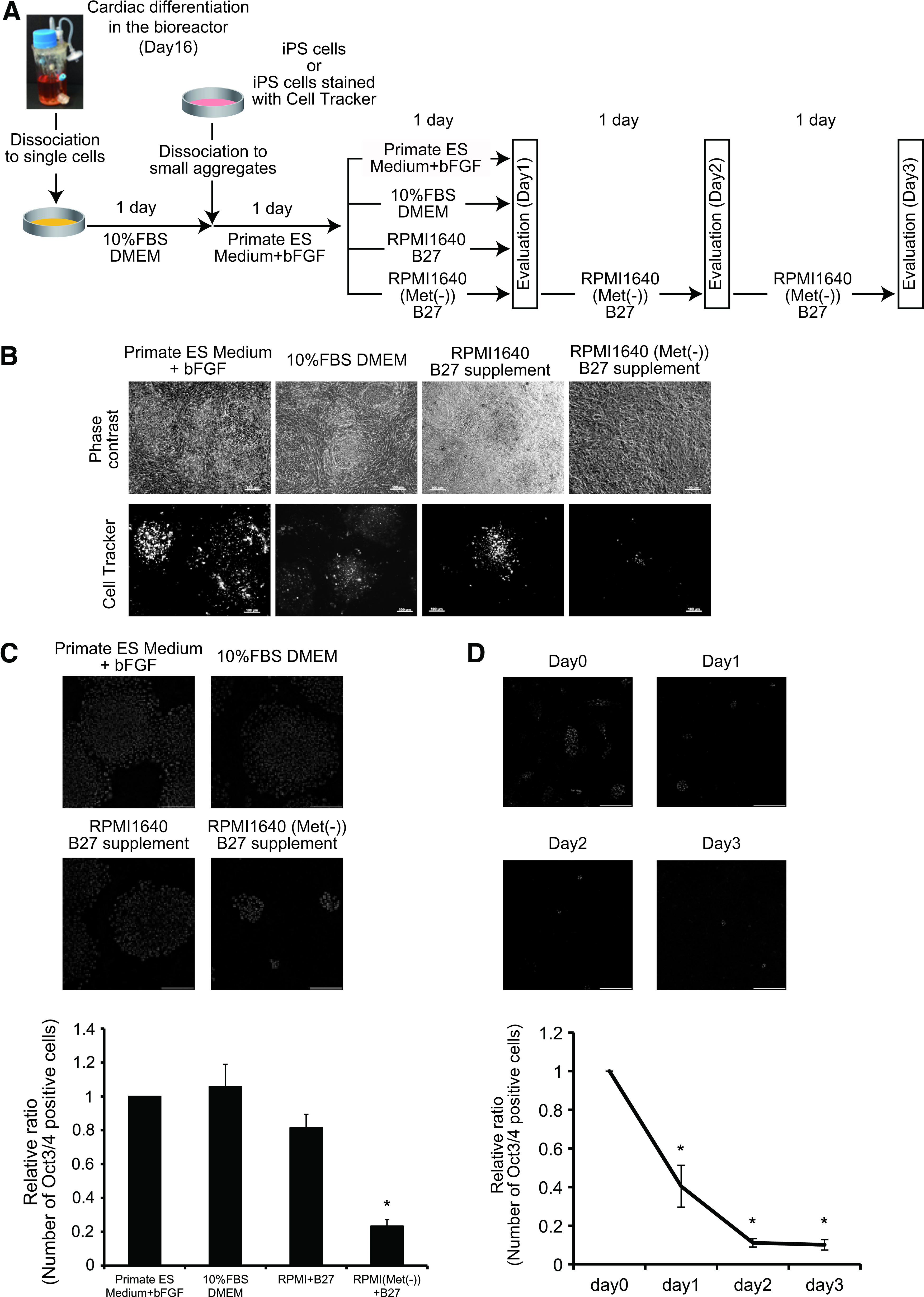
Methionine is necessary for iPS cell growth in cardiac cell sheets.
Cardiac cell sheet fabrication using a methionine-free culture condition
Next, we elucidated whether cardiac cell sheets could be fabricated in a methionine-free condition. EBs at day 16 of cardiac differentiation in the bioreactor were dissociated to a single cell suspension and cultured on temperature-responsive culture dishes in DMEM supplemented with 10% FBS for 6 h. The cells were then assigned to the following conditions: condition 1, 10% FBS DMEM for 4 days; condition 2, R(−) for 2 days and 10% FBS DMEM for the next 2 days; condition 3, R(−) for 4 days; and condition 4, R(+) for 4 days. As shown in Figure 3A, monolayer cardiac cell sheets were detached from the culture dishes when cultured in conditions 1 and 2. However, when cells were cultured in conditions 3 and 4, the cells were maintained in a confluent condition, but did not detach from the culture surface after lowering the culture temperature, which led to the failure to fabricate monolayer cell sheets (Fig. 3A). These findings suggest that serum-free culture conditions, regardless of the existence of methionine just before cell sheet harvest, did not work for fabrication of cell sheets under the applied conditions. However, the transient methionine-free culture at the early stage of cell sheet fabrication such as condition 2 might be applicable. Therefore, we performed the following experiments using cells from conditions 1 and 2.
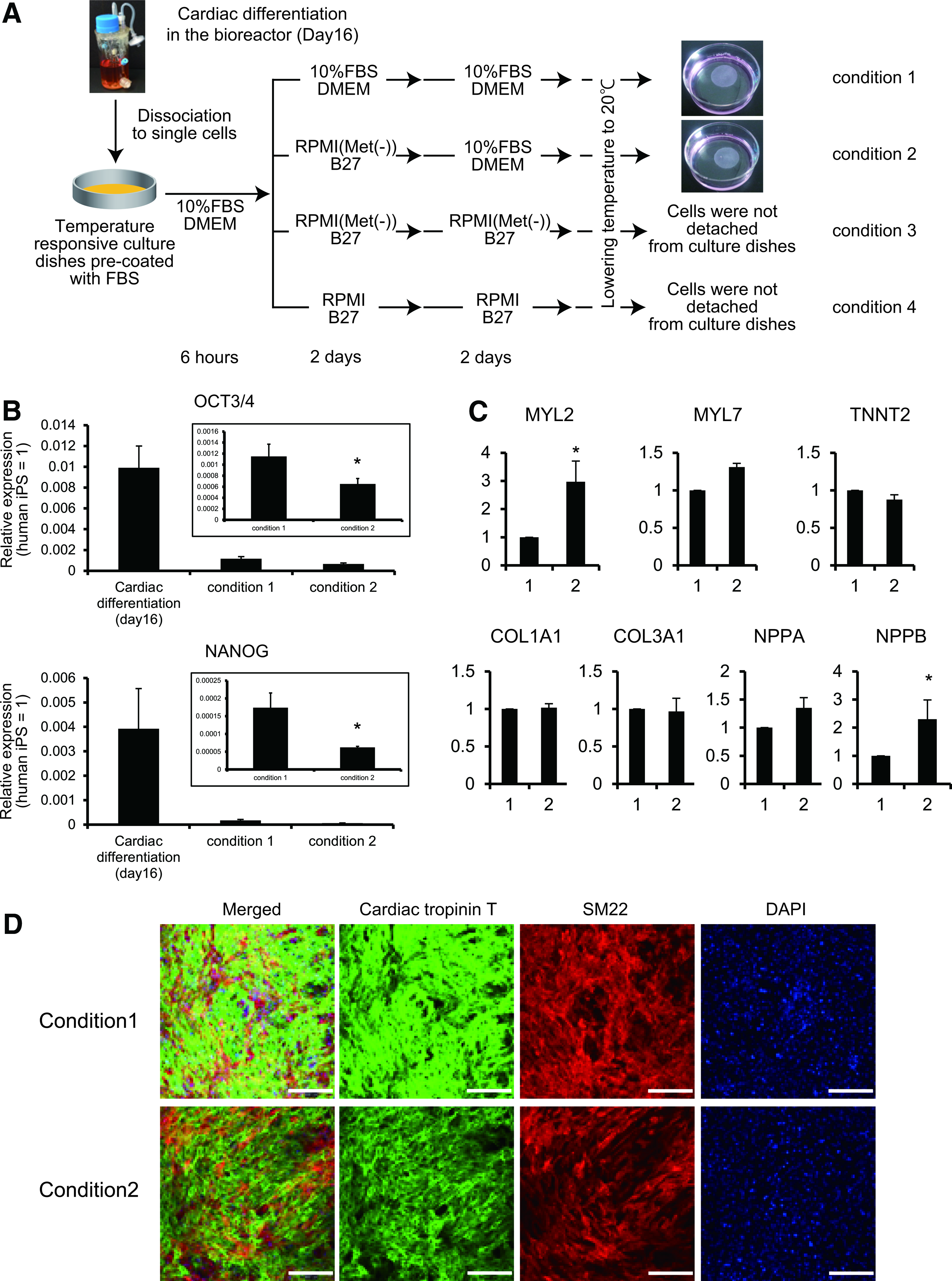
Cardiac tissue engineering using the methionine-free condition.
We previously reported that a very small number of iPS cells remain after bioreactor-based cardiac differentiation culture. 12 However, after enzymatic dissociation of EBs to a single cell suspension and 4–5 days of culture in DMEM supplemented with 10% FBS, Oct3/4-expressing undifferentiated iPS cells are undetectable. 13 Consistent with these previous findings, the expression of OCT3/4 and NANOG was downregulated in both condition 1 and 2 compared with that in cells just after cardiac differentiation (Fig. 3B). Moreover, OCT3/4 and NANOG expression in condition 2 was significantly lower than that in condition 1 (Fig. 3B). These findings suggest that a methionine-free condition in the process of cardiac cell sheet fabrication might further decrease the risk of contamination by the remaining iPS cells.
Although a methionine-free condition might be useful to inhibit the growth of iPS cells in cardiac cell sheets, the influence of methionine depletion on cardiomyocyte and noncardiomyocyte viabilities is unclear. As shown in Supplementary Videos S1 and S2 (Supplementary Data are available online at
Discussion
In this study, we have reported for the first time that methionine-free culture in the process of human cardiac cell sheet fabrication is useful to reduce the risk of contamination by the remaining undifferentiated human iPS cells. Although the methionine-free culture condition significantly prohibited the growth of iPS cells in monoculture or coculture with iPS cell-derived cells after the cardiac differentiation, the viabilities of cardiomyocytes and noncardiomyocytes were maintained and cardiac cell sheets were fabricated.
Methionine is one of the essential amino acids and contributes to various types of mammalian metabolisms such as protein synthesis as reviewed elsewhere. 25 Although methionine is important for normal growth and development, it is well known that the growth of some types of tumors, including breast cancer, colon cancer, and glioma is dependent on methionine. 18 Halpern et al. reported that human monocytic leukemia cells cannot grow in a medium supplemented with homocysteine in place of methionine, whereas the growth of human breast and prostate fibroblasts is not affected in such a culture condition. The mechanisms of the methionine dependency of tumor cells have not been clarified fully. One possible mechanism is the loss of methylthioadenosine phosphorylase (MTAP) expression, which is important for salvage of adenine and methionine. 26 Christopher et al. have reported that reintroduction of MTAP into MTAP-deficient MCF-7 breast cancer cells inhibits their growth. 27 Pluripotent stem cells are known to generate benign teratomas or teratocarcinomas upon transplantation. 21 The various possible mechanisms of malignant tumorigenicity of iPS cells have been reported, including epigenetic transformation, aneuploidy, somatic mutation, viral integration, and culture adaptation. 21 In this study, the methionine-free culture condition inhibited the survival and growth of human iPS cells in monoculture and coculture. Consistent with the evidence that adult stem cells are quiescent under normal conditions, the presence of MTAP has been reported in human hematopoietic stem cells. 28 To date, there are no reports on the presence or function of MTAP in pluripotent stem cells. Because pluripotent stem cells are quite different from adult stem cells in terms of their unlimited growth capacity, MTAP deficiency in pluripotent stem cells might regulate their growth and methionine dependency to a certain extent.
It is a prerequisite to develop suitable methods to purify target cells after differentiation and eliminate contamination of undifferentiated iPS cells for iPS cell-based regenerative medicine. Recently, several methods have been reported to purify human cardiomyocytes after cardiac differentiation of human pluripotent stem cells.14–17 Purified cardiomyocytes are indispensable for drug screening and disease research. However, purified cardiomyocytes might be insufficient for use in regenerative medicine, because purified cardiomyocytes are not able to construct cardiac tissue without fibroblasts.11,29 Furthermore, cell transplantation as a single cell suspension, but not tissue, into an infarcted heart shows limited engraftment and subsequent marginal improvement of cardiac function.22,30 In this study, we focused on development of culture methods to eliminate the remaining undifferentiated iPS cells without affecting the viabilities of cardiomyocytes and noncardiomyocytes and cardiac cell sheet fabrication. When iPS cell-derived cells after the cardiac differentiation were cultured in conditions 1 and 2 (Fig. 3A), spontaneous and synchronous beating of cardiomyocytes and expression of many cardiac and ECM genes were identical between conditions. Furthermore, the expression of MYL2 and NPPB was significantly upregulated in condition 2 compared with condition 1. Consistent with our recent findings that the percentage of ventricular type myosin heavy chain-expressing cardiomyocytes-derived human iPS cells was increased following culture on cell culture plates, 31 transient culture in a methionine-free condition might promote the cardiomyocytes maturation. On the other hand, although cell sheets were not detached from the culture surface when cultured in condition 3 (Fig. 3A), a longer period of culture in the methionine-free condition resulted in spontaneous and synchronous beating (Supplementary Video S3). This observation suggests that the methionine-free condition might not affect cardiomyocyte function. Moreover, because cell sheets were not detached from the culture surface when cultured in condition 4 (Fig. 3A), which was similar to condition 3 except for the presence of methionine, a methionine-free condition may not be the principal reason for the inability to harvest cell sheets. The detachment of cells from the temperature-responsive culture surface depends on the amount of grafted temperature-responsive polymer, Poly(N-isopropylacrylamide) (PIPAAm). 32 In the case of culture on surfaces grafted with a small amount of PIPAAm, cells can adhere to the surface at 37°C (hydrophobic surface), but cannot detach from the culture surfaces even after lowering the culture temperature (20°C, hydrophilic surface). Conversely, in the case of culture on surfaces grafted with a large amount of PIPAAm, cells cannot adhere to the surface even at 37°C. Herein, a longer culture period in serum-depleted conditions such as conditions 3 and 4 might affect ECM deposition and quantity in the basal region of cells, which leads to making the cell harvest difficult from a certain amount of PIPAAm-mediated hydrophilic surface at 20°C.
There are a few limitations in the present study. We examined the effects of methionine-free culture on the growth of two lines of human iPS cells. However, the iPS cells used in this study were produced using retroviral vectors,1,33 and methionine dependency has not been determined in genome integration-free iPS cells. 34 Although transient methionine-free culture in the process of cardiac cell sheet fabrication significantly decreased the expression OCT3/4 and NANOG, the efficacy of this method to reduce the tumorigenic risk should be elucidated in vivo with appropriate cell numbers in the future.
During the preparation of this article, the importance of methionine for the self-renewal of human pluripotent stem cells has been reported. 35 They have shown that methionine regulates intracellular S-adenosylmethionine level, the expression of p53 and NANOG, and the phosphorylation of p38, and when cultured in the methionine-free medium, many of the human pluripotent stem cells result in apoptosis and cell cycle arrest. We showed for the first time that methionine-free culture in the process of cardiac cell sheet fabrication is useful for elimination of remaining undifferentiated human iPS cells while maintaining the viabilities of cardiomyocytes and noncardiomyocytes in cardiac cell sheets.
In the present study, we have demonstrated a useful strategy to eliminate the remaining undifferentiated iPS cells in human cardiac cell sheets while maintaining cardiomyocyte and noncardiomyocytes viabilities using a methionine-free culture condition. Further understanding of the molecular mechanisms of methionine dependency in human iPS cells and the differences of metabolisms related to methionine in iPS cells and iPS cell-derived somatic cells might enable us to fabricate safer and more effective regenerative medicine products.
Footnotes
Acknowledgments
The authors would like to thank Dr. Akiyama and Dr. Iimuro for the fruitful discussion. This work was funded by a grant from the Japan Society for the Promotion of Science through the Funding Program for World-Leading Innovative R&D on Science and Technology (FIRST Program) initiated by the Council for Science and Technology Policy and a grant from Projects for Technological Development in Research Center Network for Realization of Regenerative Medicine of the Japan Science and Technology Agency.
Disclosure Statement
Teruo Okano is a founder and director of the board of Cell Seed, Inc., licensing technologies and patents from Tokyo Women's Medical University. Teruo Okano and Tatsuya Shimizu are stakeholders of Cell Seed, Inc. Tokyo Women's Medical University is receiving research funds from CellSeed, Inc. Teruo Okano, Tatsuya Shimizu, and Katsuhisa Matsuura are inventors of bioreactor systems.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.