
Abstract
Porcine skin is commonly used as a model for human skin injury and as a source material for biologic scaffold materials. Although remarkable similarities between porcine and human skin exist, regional anatomic variations present in human skin are also present in porcine skin. The objective of this study was to evaluate the structure of porcine skin from 11 different anatomic regions in the American Yorkshire crossbreed. Both qualitative and quantitative methods were used, with emphasis on epidermal and dermal thickness, hair follicle density, and collagen and elastin composition and distribution. The results showed that significant regional differences in skin histology exist, particularly with regard to the thickness of the dermis and epidermis and the amount of collagen and elastin within each tissue. Differences were also seen in the distribution of type I and type III collagen within the dermis. Therefore, while porcine skin shares many similarities with human skin, distinct regional differences in composition and morphology exist. This study highlights the importance of appreciating these regional differences to avoid misinterpretation of experimental results when using porcine skin as a human analogue.
Introduction
T
Porcine skin is perhaps the most commonly used animal tissue in biomedical research and regenerative medicine as a model for research and as a source material for biologic scaffold materials.8–20 Previous studies have shown remarkable similarities between porcine and human skin with regard to properties such as percutaneous absorption.21–23 However, it is important to realize that the same regional variations present in human skin are also present in porcine skin. Significant differences between the two species exist, and careful consideration must be made in study design to ensure the porcine skin model is an accurate representation of the equivalent human skin injury. While the regional variations in human skin have been extensively evaluated,1–4,22,24–36 there is a paucity of data on the regional variations in porcine skin structure and composition.
Since the introduction of dermal matrix in 1994, there has been a continual increase in the surgical use of acellular tissue matrices as test materials and medical devices. Dermal matrices, derived from allogeneic and xenogenic sources, are now commonly used for the treatment of extremity wounds, such as diabetic or vascular ulcers or pressure wounds; the treatment of ventral hernias; and surgical reconstruction of extremity trauma.14,37–44 The composition of the dermis and the retention of elastin and collagen fibers after decellularization impart mechanical and viscoelastic properties that enable these dermal products to provide reinforcement to the surgical site, while promoting cellular ingrowth and revascularization as the materials gradually degrade. While acellular dermal matrices (ADM) have shown notable success, a number of complications and failures have been reported.40,45,46 An appreciation of the inherent regional variations in dermal thickness across the body may contribute to improved practices in the preparation of ADM and minimize some of the variability found in these materials.
To improve the reproducibility of animal experiments and minimize the variability in acellular dermal products, a better understanding of select factors in porcine skin and their variation with regard to anatomic location is essential. For example, hair follicle density and epidermal thickness are important mediators of percutaneous absorption and wound healing21,22,24,47–49; epidermal and dermal thickness are important for the development of porcine burn models12,14; and collagen and elastin content and distribution influence wound healing,6,7,50 scarring, and the production of ADM. 51 Thus, the objective of this study was to characterize the regional variations in porcine skin from 11 different anatomic areas by both qualitative and quantitative methods, concentrating on epidermal and dermal thickness, hair follicle density, and collagen and elastin composition and distribution.
Materials and Methods
Preparation of porcine skin samples
Skin samples were obtained from six female domestic pigs, Sus scrofa, of standard American Yorkshire crossbreed stock that had been euthanized as a part of a separate study, not involving the skin. The animals were ∼3–4 months of age, and weighed 120±10 lbs. Full-thickness biopsies of skin, ∼5.0×8.0 cm were removed from 10 locations: anterior neck, inner ear, outer ear, rostral-, mid- and caudal-belly, rostral-, mid- and caudal-back, lateral fore leg, and lateral hind leg (Fig. 1). Each skin biopsy was pinned to a cork mounting board and stored frozen at −80°C until it was processed for histomorphologic assessment.
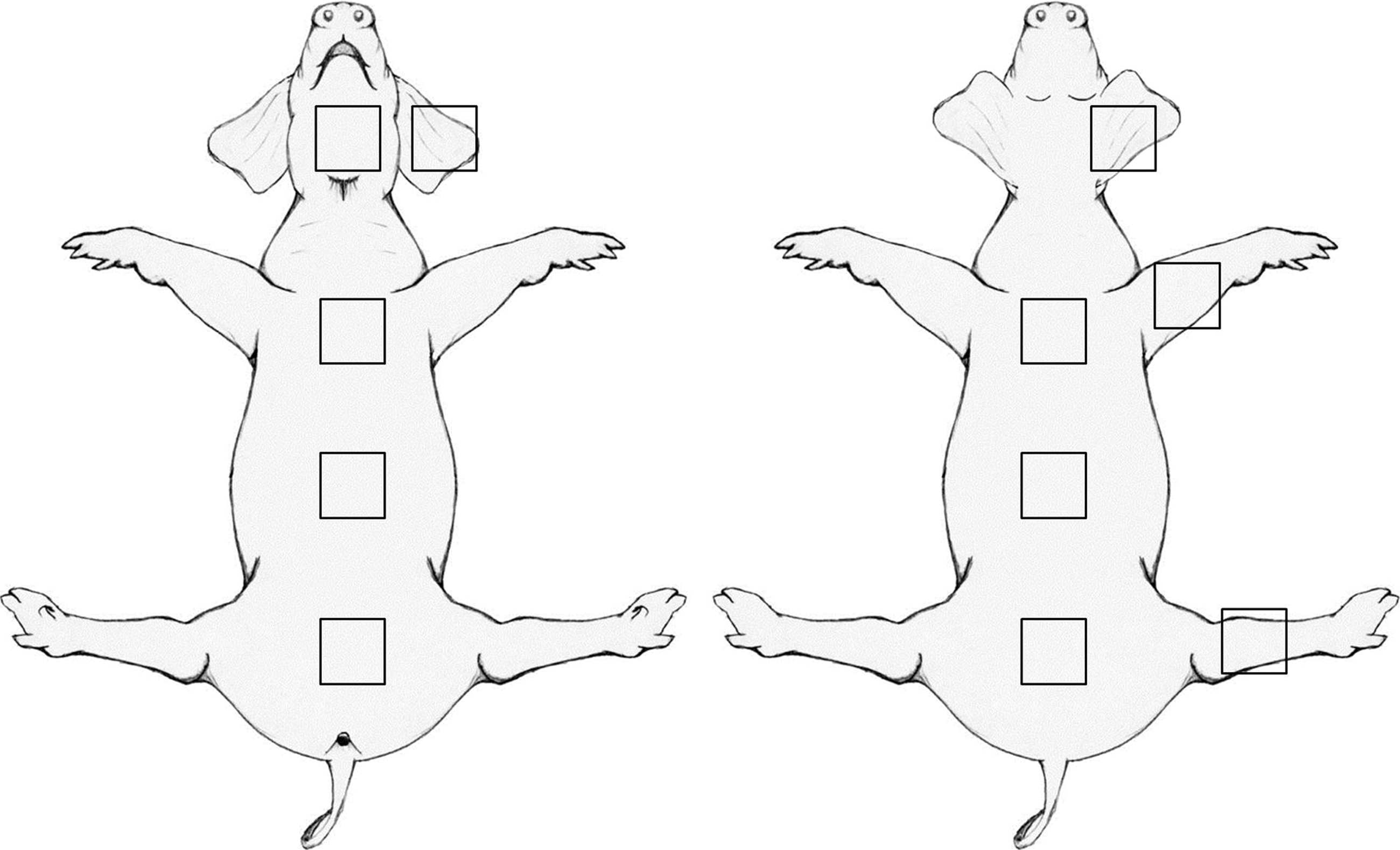
Diagram illustrating the locations on the body where skin samples were obtained.
Quantification of hair follicle density
Two 8.0 mm punch biopsies were obtained from each skin sample to assess hair follicle density. The biopsies were taken from random areas across each skin sample. The biopsies were then viewed under a stereo dissecting microscope (Nikon Instruments, Melville, NY), and the number of individual hair follicles per biopsy was recorded. Hair follicle density was then calculated as number of hair follicle per cm2.
Morphometric analysis
Morphometric analysis was performed on paraffin-embedded tissue sections stained with hematoxylin and eosin (H&E), Masson's Trichrome, Verhoeff's vanGieson, and Herovici's polychrome stains. Briefly, the frozen skin samples were thawed at 4°C overnight, and two 2.0×1.0 cm full-thickness biopsies were cut from random areas of the skin sample. For each skin sample, there was at least 1 cm of tissue separating the two biopsy sites to ensure that the biopsies were representative of the entire skin sample. The biopsies were pinned to a cork mount and fixed in 10% neutral-buffered formalin for 48 h before being processed for routine paraffin embedding. After embedding, 5 μm thick sections were cut and stained with H&E, Masson's Trichrome, Verhoeff's vanGieson, and Herovici's polychrome stains.
Stained slides were viewed using an axio-observer Z1 microscope (Carl Zeiss Microscopy, Thornwood, NY) at ×100 or ×200 magnification. Measurements of total skin thickness, and individual measurements of the corneum, epidermis, and dermis were obtained using Axiovision software (Carl Zeiss Microscopy) with measurements calibrated to the microscope and objectives used to obtain the images. The number of cell layers of the corneum, total number of Langerhans cells per field, and total number of melanocytes per field were also recorded in ×1000 magnification in six representative fields of view per specimen.
Assessment of collagen composition
Collagen composition and organization was assessed using slides stained with Herovici's polychrome and focused on collagen types I and III. Herovici's polychrome staining was performed using a modified protocol developed in our laboratory, 52 compared with that first described by Herovici. 53 Images of stained slides at ×200 magnification were obtained using a Nuance FX multispectral imaging system (Perkins Elmer, Cambridge, MA) connected to a standard light microscope (Nikon Eclipse E600; Nikon Instruments). A spectral library for Herovici's polychrome was prepared using control slides and comprised the unique spectra of acid fucshin (purple/red), methyl blue (blue), and picric acid (yellow).
After spectral unmixing, the individual color components of the image were quantified using image analysis software built in to the Nuance FX system. Color thresholding of Herovici's polychrome-stained samples was used to identify areas corresponding to type I and type III collagen within the image. The resulting thresholding values were saved with the spectral library and used to analyze all subsequent images in this study. For each skin sample, a region of interest was defined by the epidermal: dermal and dermal: subcutis borders. The percentage of image pixels corresponding to either type I or type III collagen after spectral unmixing and color thresholding was recorded for three fields of view per sample with each sample repeated in duplicate.
Assessment of elastin composition
Elastin composition and organization was assessed using two methods. The first method was a histologic assessment and quantification using Verhoeff's vanGieson staining to label the elastin fibers within the dermis and epidermis. The second method used a colorimetric assay (Fastin; Biocolor Life Sciences, County Antrim, United Kingdom) to quantify the elastin content extracted from skin samples.
Verhoeff's vanGieson staining was performed using standard histologic methods, resulting in red staining collagen and black staining elastin fibers. Images of stained slides at×200 magnification were obtained using an axio-observer microscope and axiovision acquisition software (Carl Zeiss Microscopy). The captured images were imported into Photoshop CS2 (Adobe Systems, Inc., San Jose, CA), and the replace color tool was used to remove the red collagen staining from the image to leave only the black staining of the elastin fibers. These corrected images were then imported into ImageJ, 54 where a region of interest was defined by the epidermis and dermal: subcutis border. The image was thresholded, and the percentage of black and white pixels was calculated using a macro supplied with ImageJ. The percentage of black pixels was taken to represent the percentage of elastic tissue within the skin section.
The colorimetric assay for elastin quantification utilized 5,10,15,20-tetraphenyl-21H,23H-porphine dye to analyze solubilized elastin polypeptides extracted from the skin samples. For each skin sample, one 8.0 mm punch biopsy was obtained and lyophilized until completely dry. The mass of the dried tissue samples was recorded and then, the tissue samples were solubilized in 750 μL 0.25 M oxalic acid at100°C for 60 min. The specimens were then centrifuged at10,000 rpm for 10 min, and the supernatant was retained for analysis. This solubilization step was further repeated twice on the residual tissue to ensure complete elastin extraction. Quantification of the solubilized elastin was performed using the Fastin Elastin Assay (Biocolor Life Sciences) following the manufacturer's recommended protocol with elastin content expressed as μg elastin/mg of dry weight tissue.
Statistical analysis
For all quantitative assays, for which data passed tests for normality and equal variance, two-way analysis of variance (ANOVA) was performed to determine differences between sample sites and whether any difference existed between animals. If no difference between animals was found, a one-way ANOVA with Tukey's post hoc test was performed to identify specific differences between body sites. Where data failed tests for normality, a Kruskal–Wallis one-way analysis of variance on ranks was performed. A value of p≤0.05 was considered significant. All tests were performed using SigmaPlot software (Systat Software, Inc., San Jose, CA).
Results
A two-way ANOVA showed no significant differences between animals for all statistical tests.
Histology of porcine skin
Porcine skin samples were evaluated by histologic methods on paraffin-embedded sections stained with H&E and Masson's Trichrome. A keratinized outer layer and epidermal and dermal layers could be clearly identified in all specimens. It was not possible to discern a distinct papillary or reticular layer within the dermis by H&E or Masson's staining. All skin samples were devoid of eccrine glands, although occasional sebaceous glands and apocrine glands were present surrounding hair follicles. Representative images of skin structure from each body site are shown in Figure 2.

Summary of the histologic appearance of porcine skin from different body locations
The keratinized layer of the epidermis at each body site generally consisted of between 5 and 17 cell layers with the average thickness being 18.88±2.58 μm. The keratinized layer was thickest in the hind leg and thinnest in the mid back with average thicknesses of 24.37 and 15.41 μm, respectively. However, counting the number of cell layers showed that the caudal belly skin had the most layers with 15.93±0.44, while the mid back had the smallest number of layers with 4.97±1.01 (Fig. 3A). While there were significant differences in the thickness and number of cell layers of the corneum between different body sites (see Supplementary Tables S1–S10; Supplementary Data are available online at
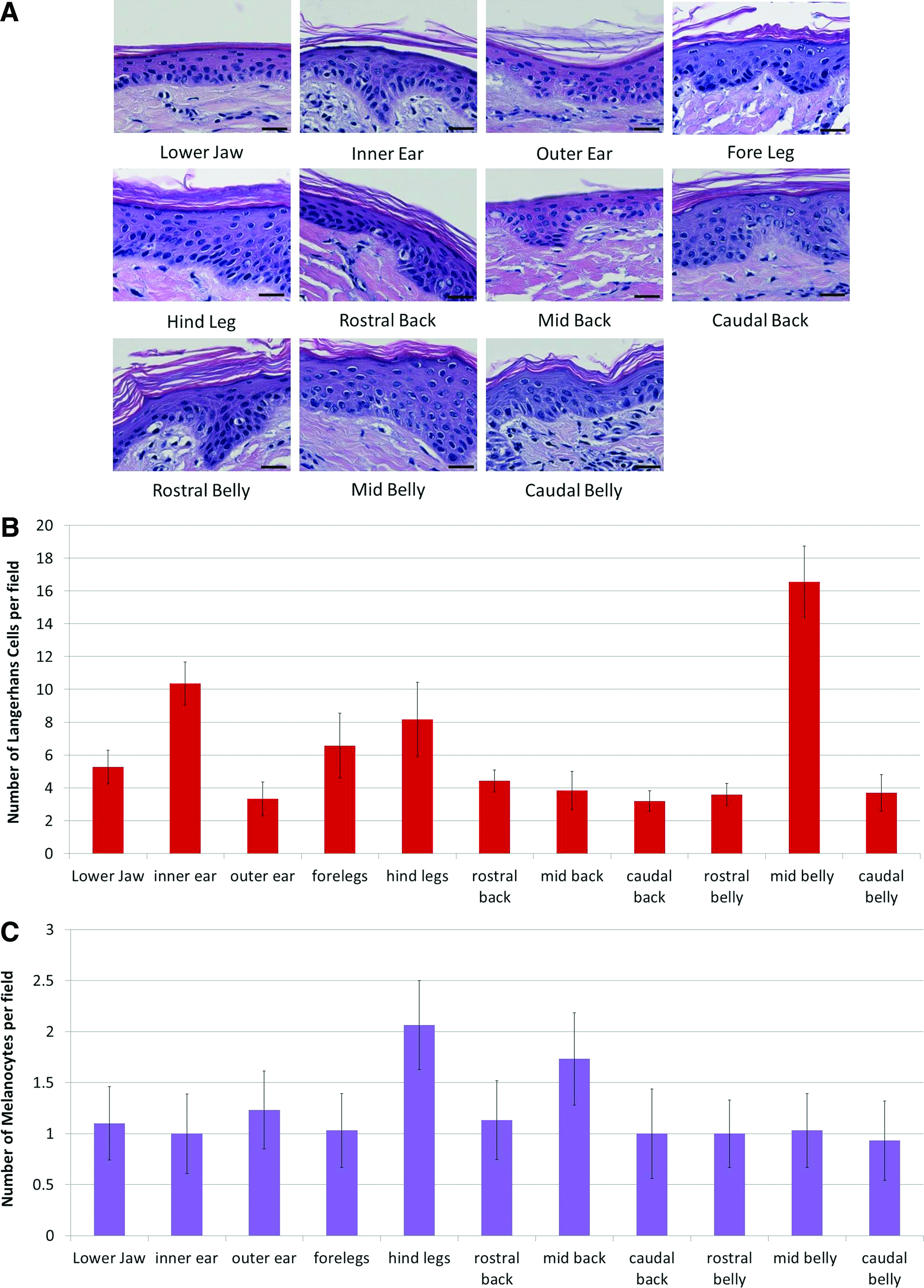
Summary of the histologic appearance of porcine epidermis from different body locations
Quantification of the number of Langerhans cells, the antigen-presenting cells of the epidermis, and melanocytes, the pigment-producing cells of the epidermis, showed differences in the cellular composition of the epidermis between body sites (Fig. 3B, C). The number of Langerhans cells was the highest in the mid belly with an average of 16.57±2.15 cells per field and the lowest in the outer ear and caudal back with 3.33±1.01 and 3.20±0.67 cells per field, respectively. Comparing related body sites showed differences in the number of Langerhans cells between the mid belly, rostral belly, and caudal belly and the inner versus outer ear. There were also differences between unrelated body sites, which can be found in Supplementary Data (Supplementary Tables S1–S10). Quantification of the number of melanocytes showed much less variability between skin locations, although differences were still apparent. In general, the number of melanocytes per field varied between 1 and 3 cells per field with the hind leg having the highest number of cells with an average of 2.07±0.43 cells per field; other body sites generally average one cell per field. Consequently, there were no statistical differences in melanocyte counts between different body sites or within related body sites except between the fore and hind legs. Complete statistical analysis can be found in the Supplementary Data (Supplementary Tables S1–S10).
The dermis showed the greatest variability in thickness between the different body sites (Fig. 2D). The dermis of the inner ear was the thinnest with an average thickness of 796.33 μm while the dermis in the caudal back was the thickest averaging 1687.67 μm, more than twice as thick as the dermis of the inner ear. Comparing related body sites showed that in general the thickness of the dermis increased rostro-caudally. While there was no difference in thickness between the inner and outer ear or between the rostral, mid, and caudal belly, the dermis increased in thickness between the rostral and caudal back and between the fore and hind legs (Fig. 2D). Results of statistical comparisons between all body sites can be found in Supplementary Data (Supplementary Tables S1–S10).
Despite these changes in dermal thickness, the histologic appearance of the dermis varied little between the different body sites. In general, the dermis was composed of dense interlacing bands of fibrous tissue (Fig. 2A). These fibers appeared to give a cross-hatch appearance to the dermis with some fibers arranged longitudinally and others transversely. The dermis was relatively acellular in comparison to the epidermis, with individual cells dispersed throughout the dermis. Occasional clusters of cells were visible, corresponding to hair follicles, blood vessels, or apocrine glands. Blood vessels were most evident in the upper dermis, while apocrine glands were typically observed at the junction between the dermis and subcutaneous fat.
Analysis of hair follicle density
Porcine skin is a preferred in vitro model for the study of percutaneous absorption.11,55 Hair follicle density is an important modulator of absorption 56 and, is, therefore, an important consideration when designing percutaneous absorption experiments. In addition, the presence of hair follicles and the stem cell population associated with them may influence the rate and extent of wound healing.47–49 Hair follicles in the skin samples were sparsely arranged as single follicles rather than in clusters of two or more follicles. The density of hair follicles varied greatly across the different body sites (Fig. 4). The highest density of hair follicles was in the skin of the outer ear with 42±11 follicles per cm2. The lowest density was in the belly skin with the caudal belly having only 4±2 follicles per cm2. There was no significant difference in the follicle density between the different belly skin sites, or between the different sites on the back. However, there was consistently more than double the hair follicle density on the back versus the belly (Fig. 4). No difference was seen between the fore and hind legs. The greatest difference was seen between the inner and outer ear. Full details of the significant differences observed between all sampling sites can be found in the Supplementary Data (Supplementary Tables S1–S10).
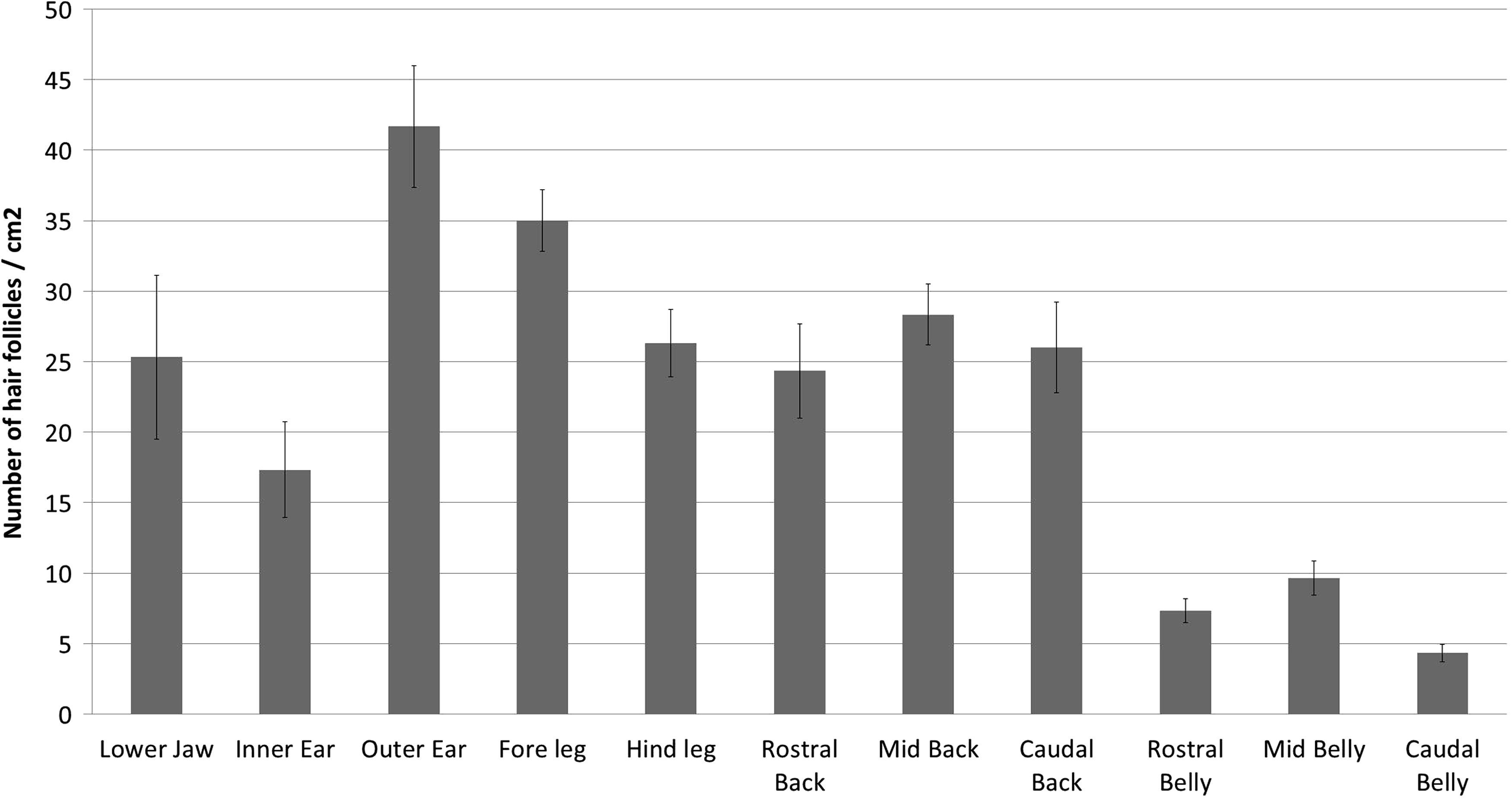
The average number (±SEM) of hair follicles per cm2 for each body site tested.
Analysis of type I and type III collagen content
Collagen in porcine skin has a similar composition to human collagen 57 and elicits a minimal immune response, making it ideal for production of ADMs. The mechanical properties of the skin depend largely on the organization of the collagen fibers within the skin, while elasticity and extensibility are determined in part by the proportions of type I and type III collagen present. Given the use of porcine skin as an experimental model and for production of ADMs, an understanding of the arrangement and proportion of these fibers is important.
The collagen composition of the dermis in the different skin samples was assessed by semi-quantitative assessment of Herovici's polychrome staining. A representative example of Herovici-stained skin is shown in Figure 5. While it was not possible to discern the papillary and reticular layers of the dermis by H&E or Masson's trichrome, staining with Herovici's polychrome clearly identified two distinct layers in the dermis based on their collagen content. Immediately below the epidermis, the dermis was predominately composed of type III collagen with little type I collagen visible. Below this layer, type I collagen was predominately present in the dermis as large-diameter fibers (Fig. 5A). Using image analysis and color thresholding, 52 the dermis was isolated and the collagen composition was quantified. Type III collagen content averaged 10.10%±0.78% of the dermal extracellular matrix in all samples (Fig. 5B). There were no significant differences in type III collagen content between related body sites but differences were seen between the lower jaw versus caudal belly, fore leg and hind leg, and between the hind leg versus inner ear, outer ear, rostral back, and mid belly. Greater variation was seen in in the type I collagen content, which ranged from 23.46% in the rostral belly to 29.05% in the mid back (Fig. 5B). Expressing these values as a ratio showed that porcine skin typically has a collagen I:III ratio of ∼2.5:1. While significant differences in type I collagen content were seen between multiple body sites (see Supplementary Data Supplementary Data [Supplementary Tables S1–S10]), comparing related body sites show no difference in type I collagen content between inner and outer ear, fore and hind leg, or between belly skin samples. A difference was seen between rostral back and mid back skin.
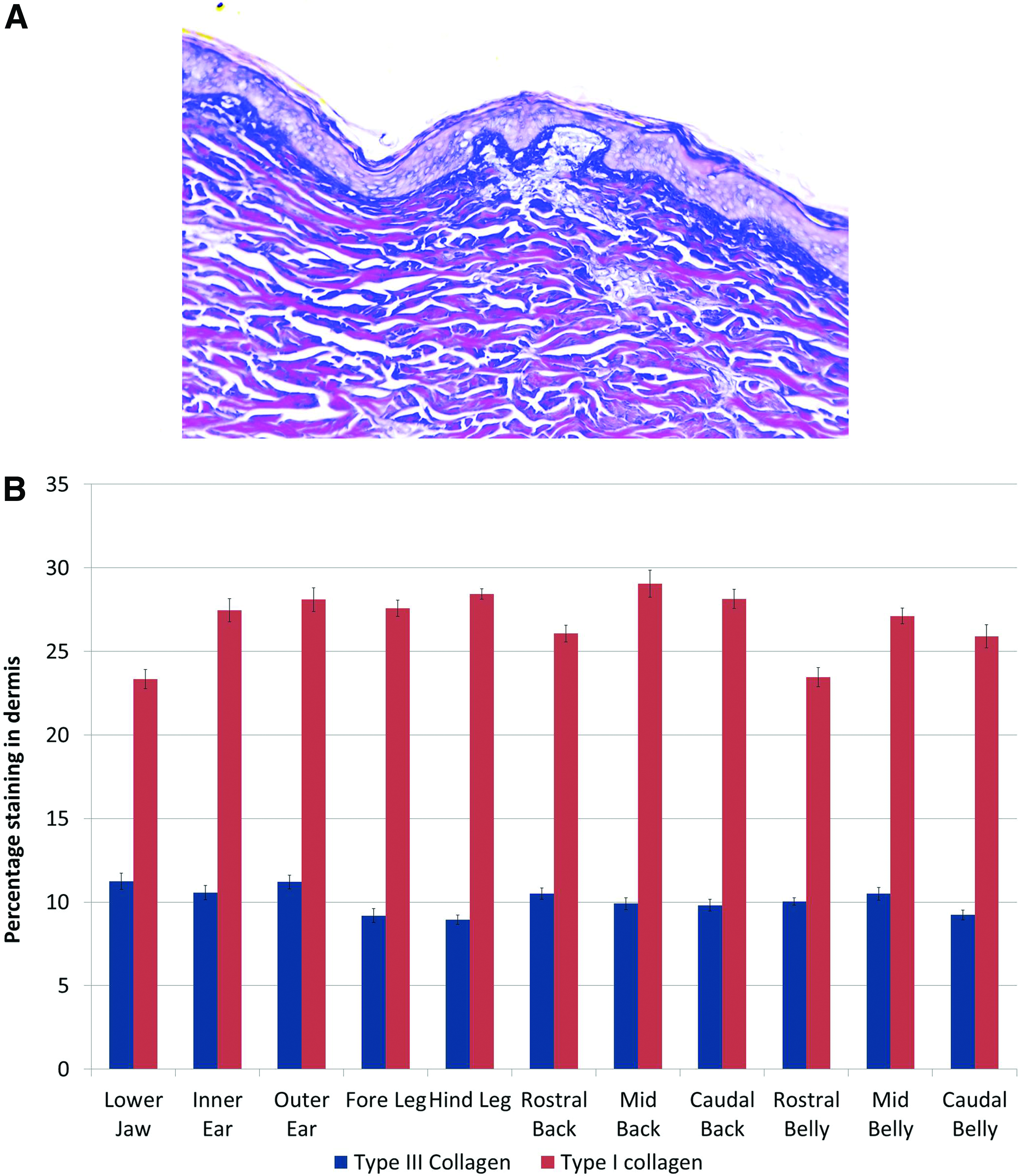
Quantification of Herovici's polychrome staining for type I and type III collagen. Representative staining
Analysis of elastin content
Elastin provides recoil to the skin, enabling it to recover when pinched or compressed. The elastin fibers provide an important signaling network and may play a role in wound healing and repair, particularly mediating the extent of scar tissue formation.58,59 Porcine tissue is known to have a high elastin content, 60 and it has been suggested that the elastin content in ADMs may be important for improved elastin regeneration after burn injury.
Elastin content in the different skin samples was assessed by two methods. First, skin samples were stained with Verhoeff's vanGieson to visualize the elastic fibers (Fig. 6A). This staining was then quantified by image analysis (Fig. 6B). Second, biopsy samples from the skin sites were digested in oxalic acid and the elastin was isolated and quantified using a biochemical assay (Fig. 6C). Comparison of the semi-quantitative image analysis and quantitative elastin assay showed a similar pattern of elastin content, although some differences in quantification were seen. The greatest difference between the two assays was in the ear skin, which had the lowest percentage elastin by image analysis but the highest elastin content when quantified biochemically.
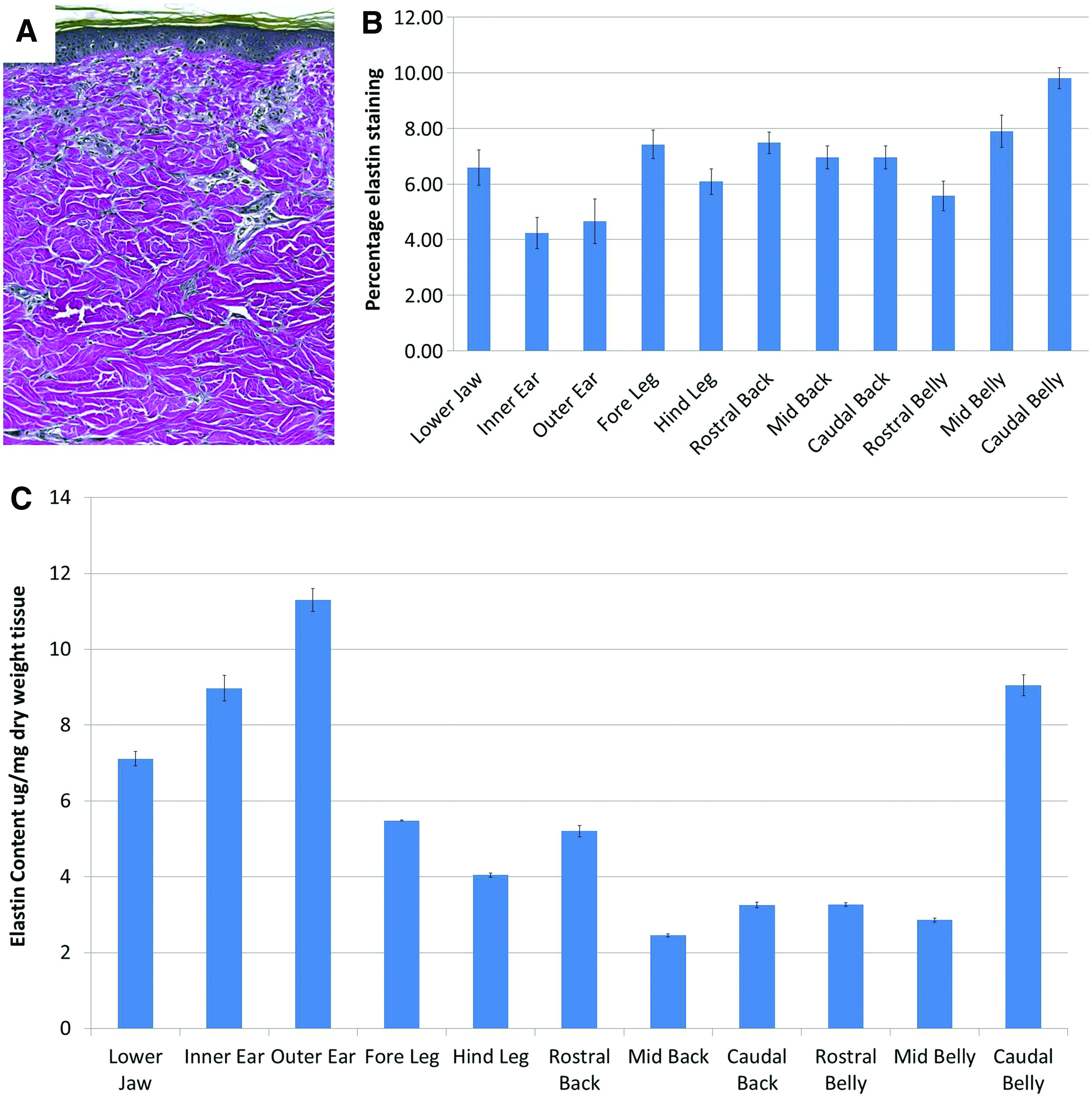
Quantification of elastin staining within porcine skin samples. Skin samples were stained with Verhoeff's vanGieson stain
In general, the elastin content of the skin samples was low, ranging between 4.24% and 9.8% of the dermis by image analysis or 2.45 and 11.30 μg/mg by elastin assay. Staining with Verhoeff's vanGieson stain showed that the elastin fibers were found in the highest density in the upper dermis, appearing as long thin-diameter fibers (Fig. 6A). Image analysis showed that elastin content was the highest in the caudal belly with elastin content decreasing rostrally. There was no difference in elastin content in skin samples from the back or between the fore and hind legs or inner and outer ear.
Quantitative assessment using the elastin assay appeared more sensitive and detected greater differences between the body sites. Elastin content was highest in the outer ear compared with all other body sites. The caudal belly skin also contained high levels of elastin and was higher than rostral or mid belly skin, which themselves were not different from each other. Differences in elastin content were also seen between the fore and hind legs and the rostral, mid, and caudal back.
Discussion
The histology of porcine skin from 11 different anatomic sites was studied. While other studies have investigated the characteristics of porcine skin, these studies have focused on specific aspects such as the enzymatic properties and collagen fiber alignment or have been limited to analysis of skin from a specific location.9,11,34,61–65 This study shows that significant regional differences in skin histology exist between various body sites and suggests that consideration regarding anatomic location should be made when selecting porcine skin as an analogue for human skin either for in vivo studies or in the development of biologic scaffolds derived from decellularized dermis.
The pig has been utilized in numerous studies involving wound healing, the study of burns and scalds, and scar tissue formation.12,14,50,66,67 In addition, porcine skin, and porcine dermis in particular, have been used in the production of biologic scaffolds and used as surgical meshes for hernia repair and in the treatment of burn injuries, although their success has been mixed.16–18,37,41,43,46,68 The use of porcine skin as an experimental model or as a source of biologic scaffolds has resulted from investigations into the properties of skin of domestic animals which showed that on all the animals studied, the pig offered the most appropriate model. 34 However, it has been shown that regional differences exist in the composition and morphology of human skin from different body sites, and it is, therefore, likely that similar variations exist in porcine skin. Determining the appropriateness of porcine skin as an analogue for human skin should logically factor in these regional differences, and any experimental model should match the morphology and composition of the likely human application site. As such, a comparison or the results of this study to the properties of human skin is warranted.
The average hair follicle density in the porcine skin samples ranged from ∼4 hairs/cm2 in the caudal belly to 42 hairs/cm2 in the outer ear. In contrast, human skin, with the exception of the head, typically ranges between 14 and 32 hairs/cm2. 28 While this is similar to the hair follicle density found in the porcine skin, the greater variability in the pig skin samples suggests that notable site-to-site variation may exist. The average thickness of the keratinized layer of porcine skin in this study was ∼18.88 μm ranging between 15.41 and 24.37 μm and consisted of between 5 and 17 cell layers. In comparison, while the keratinized layer of human skin typically ranges between 6 and 19 μm depending on the body site and shows a strong similarity to the porcine skin,1,29,31 the human corneum has been shown to contain between 6 and 30 cell layers depending on the region1,31 with the skin of the genitals, face, and neck having the least number of layers, while areas such as the palms and heels may have between 40 and 90 layers. 31 Comparing the viable epidermis thickness shows not only some similarities to human skin but also notable differences. Reported thicknesses of human epidermis vary by body site with reported thicknesses of 70 and 82 μm in the shoulder and buttock, respectively, 29 and generally between 50 and 120 μm. 34 Comparing similar body sites with those used in this study, human adult (23–53 years) epidermal thickness in the neck, abdomen, back, anterior arm, and anterior leg are reported as 130, 179.3, 97, 113, and 143.9 μm, respectively. 25 In comparison, the same measurements in the porcine skin samples were 51.7, 64.9, 45.8, 86.1, and 91.7 mm for the lower jaw, mid belly, mid back, fore, and hind legs respectively, considerably thinner than the comparative human location. The epidermal thickness in pigs has been reported by others to be between 30 and 140 μm.34,69 Langerhans cells are important immune mediator cells of the epidermis. In general, they are thought to comprise 3% of the epidermal cells, but the results of this study show distinct variations in the distribution of these cells by body location. 70 The highest concentration of Langerhans cells was found in the mid belly skin and inner ear, and these differences in distribution may indicate the relative pathogenic insult that each region is exposed to.3,4,71 Melanocytes provide pigmentation to the skin, preventing UV light damage to the hypodermis. The American Yorksire breed of pigs used in this study are a large white pig strain with little skin pigmentation; as a result, the numbers of melanocytes within the pig skin was low with only 1–3 melanocytes per field. In humans, melanocytes make up between 5% and 10% of the basal epithelial cells with around 1000–2000 melanocytes per square millimeter of skin.24,72,73.
Similar regional differences also exist in dermal thickness. In humans, the dermis typically ranges from around 1.9 mm in the back to ∼1 mm in other body regions.25,30 The porcine skin samples in this study had a dermal thickness ranging from ∼1.7 mm in the caudal back to 0.7 mm in the outer ear, showing great similarity to the regional patterning in humans, although they remained thinner than the equivalent human site. One possible explanation for the differences in epidermal and dermal thickness may be related to age of the sample population. All the skin samples used in this study came from pigs between 3 and 4 months of age, the typical age for use for in vivo studies. Previous studies have shown that, in terms of organ maturity, pigs at 4 months of age are analogous to humans in their mid-teens.74,75 Therefore, the skin samples in this study may not have reached full maturity and would likely be thinner than adult skin. In humans, epidermal and dermal thickness has been shown to vary significantly as a consequence of age in multiple body sites.25,29 In addition, this study investigated only one strain of pig, American Yorkshire crossbreed, and the regional variations in skin morphology in this strain may differ considerably from other pig breeds commonly used for in vivo studies.
Together, the results on porcine skin thickness and hair follicle density provide important information for studies such as percutaneous absorption where regional differences in skin morphology may influence absorption rates. Previous studies have shown the importance of hair follicle density on the penetration of topically applied substances in both humans and pigs, while regional differences in skin morphology have also been shown to influence percutaneous absorption. The results of this study support these previous studies demonstrating that while on average porcine skin is remarkably similar to human skin, notable differences can exist on a site-by-site comparison. Differences in the thickness of the dermis and epidermis within related body sites in the pig may also influence the production of biologic scaffolds derived from porcine dermis. Typically, these scaffolds are prepared from the skin on the back between the shoulders and the hips, with the epidermal and subcutaneous fat layer mechanically delaminated to a uniform thickness. 51 In this study, the dermis increased in thickness between the rostral and caudal back samples while the epidermis showed a variable thickness, being thicker in the caudal back and thinnest in the mid back. This variability in thickness may influence the preparation of biologic scaffolds, as the mechanical delamination process may result in tissues from different depths in the dermis being retained. As a result, there may be differences in mechanical properties, degradation rate, and composition of the final scaffolds as a result of composition differences in the starting material.
Both collagen and elastin content of the dermis showed differences in distribution within the dermis as well as variations between different body sites. Most notable was the presence of a predominately type III collagen layer in the upper dermis beneath the epidermis. While the porcine skin samples had no distinct papillary or reticular layering to the dermis, the presence of this collagen layer indicates that differences in the composition of the dermis exist that are similar to those found in humans. The dense, predominantly type I collagen, deep dermis in the porcine skin samples showed many similarities to the architecture described by others with large fiber bundles arranged in a complex meshwork,62,64 although the ratio of type I:III collagen in the porcine dermis was slightly lower than has been reported for human skin. 52 Elastin content showed the greatest variation between skin samples. Previous studies have shown that porcine skin has a distinct elastin network that is interwoven with the collagen fibers in the dermis. This study showed that the elastin component comprised between ∼4% and 10% of the dermis when quantified by image analysis. Interestingly, Hult and Goltz showed that human dermis contained ∼4.16–9.4% elastin when quantified by enzymatic digestion, 35 suggesting that the composition of porcine skin closely resembles that of humans, although others have report much higher concentrations. 60
In conclusion, the pig arguably presents the most suitable experimental model for use in research studies on human skin. The results of this study show that while porcine skin shares many similarities with human skin, in the American Yorkshire crossbreed, distinct regional differences in composition and morphology exist. However, many body sites on the pig share similarities to human skin with the skin in the rostral and mid back regions being closest in thickness, hair follicle density, and composition to human skin. The results on skin thickness and hair follicle density also provide important information for studies such as percutaneous absorption. Moreover, the variability in back skin thickness may influence the preparation of biologic scaffolds where variable dermal thickness may alter the mechanical properties, degradation rate, and composition of the final product. To avoid misinterpretation of experimental results that utilize porcine skin, care should be taken to appreciate these regional differences.
Footnotes
Acknowledgment
The authors would like to thank McGowan histology lab for preparation of the histologic slides.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.