
Abstract
The aim of this study was to evaluate how low-intensity pulsed ultrasound (LIPUS) modulates the effect of transforming growth factor-β3 (TGF-β3) on the differentiation of scaffold-free dedifferentiated bovine articular chondrocyte tissues toward a cartilage-like phenotype. Specifically, the effect of these stimuli on the expression of hypertrophic markers collagen type I, collagen type X, and cartilage-degrading collagenase gene expression for a scaffold-free model was analyzed. A bioreactor that applied LIPUS directly from the transducer through a silicone gel to a six-well plate containing the tissues allowed simple, sterile, and large-scale experiments. Tissues were subjected to LIPUS of 55 mW/cm2 in a 200 μs burst sine wave of 1 MHz over a 10-day period with or without TGF-β3 (10 ng/mL). Tissues exposed to TGF-β3 had significantly increased glycosaminoglycan and total collagen protein production along with upregulated cartilage-specific gene expression, resulting in tissues with a higher Young's Modulus. However, these tissues had also upregulated gene expression for hypertrophic markers collagen type I, collagen type X, MMP-1, MMP-13, MMP-2, and also an increase in the phosphorylation of p38. The expression of these matrix-degrading enzymes was remediated by hypertrophic development and differentiate dedifferentiated bovine articular chondrocytes towards a chondrogenic lineage allowing it to be a valuable tool in cartilage tissue engineering.
Introduction
T
Although traditional methods of tissue engineering typically use scaffolds, scaffold-free tissues are of increasing interest as there is immediate biocompatibility and less material is required to produce the tissues. The pellet culture system is most commonly used, but this system is limited to one culture per tube and a spherical shape. Other methods include the use of transwell membranes, 8 agarose gel, 9 alginate gel,10,11 rotational wall vessels, 12 and a suspension culture of aggregating chondrocytes.9,13 These methods have been shown to induce the chondrogenesis of MSCs and the redifferentiation of dedifferentiated chondrocytes.
Soluble factors, such as bone morphogenetic proteins (BMPs), insulin-like growth factors (IGFs), fibroblast growth factors (FGFs), and transforming growth factor-β (TGF-β), are used to manipulate the environmental signals sent to the cells. Although each of these growth factors is effective individually, research showed that there were no additive effects with the combination of TGF-β with BMP-2 and IGF-1. 14 In a study between TGF-β, IGF-I, and IL-4 on bovine articular chondrocytes seeded in polyglycolic acid scaffolds, TGF-β was shown to be the only growth factor to increase collagen protein production. 15 TGF-β is a peptide that can regulate cell growth and differentiation, tissue repair, and extracellular matrix (ECM) production; however, the effects of TGF-β are critically dependent on the experimental conditions and the stage of differentiation of the target cell. For instance, when TGF-β was used in human MSCs cultured in collagen type I hydrogels, cartilage-specific ECM was produced. 16 The application of TGF-β2 and -β3 resulted in a higher glycosaminoglycan (GAG) production in human MSCs cultured in the pellet form. 17 Conversely, collagenase-3 mRNA in nonlesional human chondrocytes was upregulated with the treatment of TGF-β. 18
Mechanical stimulation can be used to alter the signaling of cells; for instance, low-intensity pulsed ultrasound (LIPUS) can modulate the metabolism of cells19,20 and induce cartilage matrix formation.21,22 In a human mesenchymal stem cell (hMSC) pellet model exposed to LIPUS, the aggrecan protein content was enhanced and chondrogenesis occurred. 21 LIPUS treatment also delayed the expression of collagen type X of primary rat femur chondrocytes in aggregate culture 19 and increased aggrecan gene expression in a rat femur fracture model. 20
In this study, the modulation of the TGF-β3 treatment through LIPUS on scaffold-free dedifferentiated bovine articular chondrocyte tissues was examined. We demonstrated that LIPUS treatment can effectively downregulate the mRNA expression of cartilage degrading enzymes and prevent the phosphorylation of p38, a hypertrophic marker, initiated by TGF-β3 treatment in a scaffold-free model with a customizable shape. Through these results, we propose that LIPUS can be used to modulate the effect of TGF-β3 on the chondrogenic differentiation of dedifferentiated bovine articular chondrocytes and may be an effective tool for the engineering of cartilage tissues for the treatment of cartilage diseases.
Materials and Methods
Isolation and culture of bovine articular chondrocytes
Full-thickness articular cartilage was isolated from 4- to 6-week-old bovine calves within 24 h of slaughter. To prevent contamination, tissues around the joints were kept intact and refrigerated immediately after slaughter. Cartilage specimens were extracted using a sterile scalpel blade, washed twice with phosphate-buffered saline (PBS), minced, digested by 0.2% w/v type II collagenase (Worthington Biochemical Corp) in Ham's F12 medium (Life Technology) for 12 h at 37°C in a 50-mL polypropylene tube, and then filtered through a 70-μm cell strainer (Becton Dickinson Labware). The cells were suspended in PBS, centrifuged at 1500 g for 10 min, and then resuspended in Ham's F12 medium supplemented with 10% fetal bovine serum (Gibco BRL), 5 ng/mL FGF-2, (Peprotech), 100 U/mL penicillin, 100 μg/mL streptomycin (Gibco BRL), and 0.5 μg/mL fungizone. Cells were seeded onto 24.5×24.5-cm2 square dishes at a density of 1.6×105 cells/cm2 and cultured at 37°C, 5% CO2, and 95% humidity. After 5 days of culture, cells were detached using 0.5% trypsin/1.0 mM EDTA and replated at 1000 cells/cm2.
Scaffold-free tissue constructs
Bovine articular chondrocytes at passage 2 were utilized to ensure that the cells were properly dedifferentiated.
23
These cells were subcultured and washed twice with Dulbecco's Modified Eagle's Medium (DMEM)-high glucose and resuspended with a chondrogenic medium consisting of DMEM-high glucose supplemented with 100 nM dexamethasone (Sigma), 50 μg/mL sodium pyruvate, 50 μg/mL ascorbic acid 2-phosophate (WAKO Pure Chemicals), 40 μg/mL
Treatment with LIPUS and TGF-β3
The ultrasound emission apparatus was provided by Sigmax, Japan, and consisted of a power supply, function generator (model USE-MODL-004), and three transducers (effective area 9.6 cm2 per transducer) connected to a timer that automatically applied the LIPUS each day (Fig. 1). A mounting unit was designed to hold a six-well culture dish directly above the transducers, with the surface area of each transducer matching exactly the area of a single well in a six-well plate. A LIPUS signal in a square wave with 200 μs tone bursts at a frequency of 1 MHz was delivered to the tissues from the LIPUS transducers through a silicone gel coated with ultrasound gel (Aquasonic).
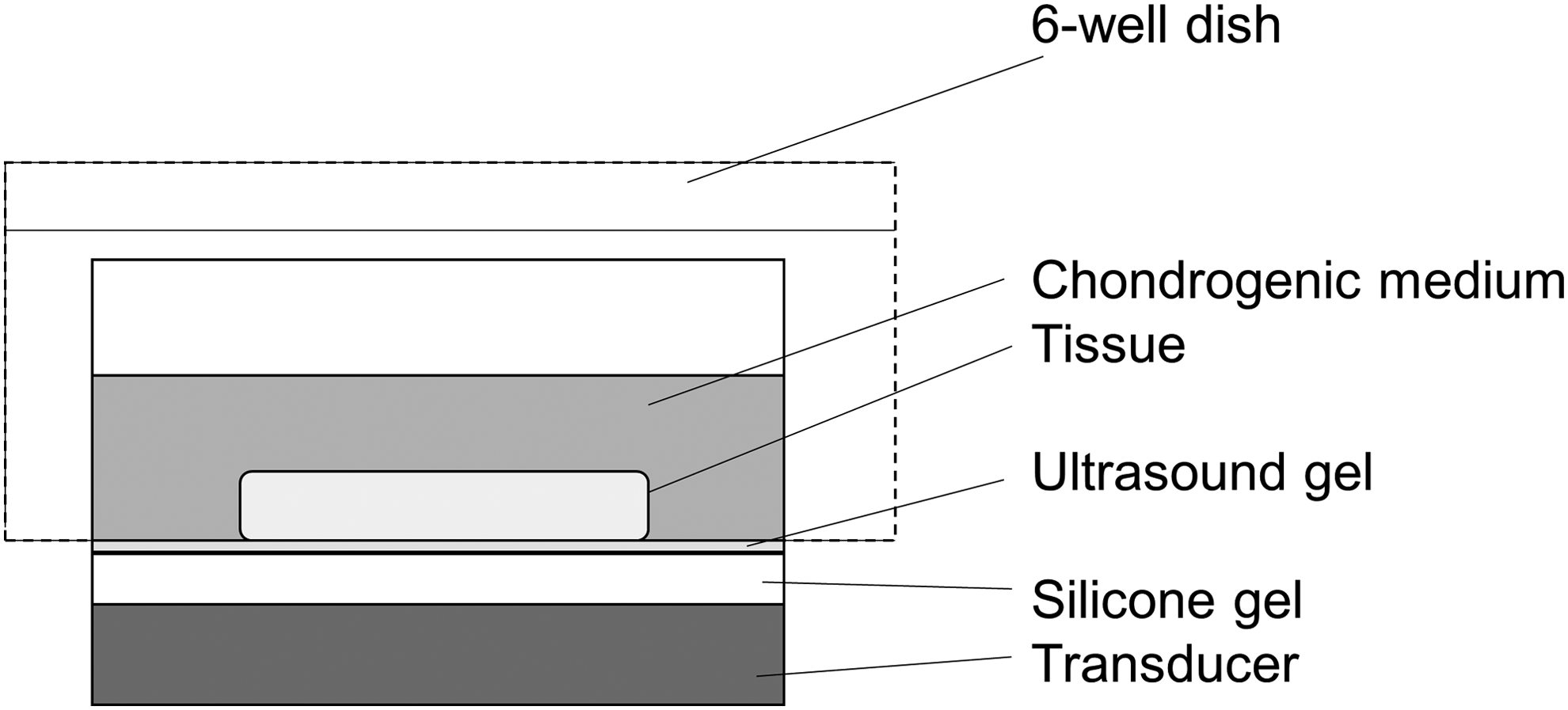
Apparatus for low-intensity pulsed ultrasound (LIPUS).
Preliminary experiments tested the effect of LIPUS intensities of 30, 55, and 120 mW/cm2 at 1 and 2 MHz on the differentiation toward a chondrogenic lineage of dedifferentiated chondrocytes at 20 min/day over a 10-day period. From the results, the optimal intensity of 55 mW/cm2 was determined and then utilized for experiments with TGF-β3. LIPUS treatment was applied for 20 min per day over a 10-day period. A 20 min/day of exposure was chosen based on experiments from Ebisawa et al. (2004), Mukai et al. (2006), and Nishikori et al. (2002). Tissues were divided into four groups: control (no LIPUS or TGF-β3 applied), LIPUS alone, TGF-β3 (10 ng/mL) alone, and LIPUS+TGF-β3. All tissues were cultured in 5 mL of chondrogenic medium per tissue per well. Each treatment had three samples and was repeated with three different cows.
Gene expression
Tissues were washed with PBS, frozen by liquid nitrogen, and then homogenized using a polytron into a powder form. The samples were then dissolved with TRIzol (Gibco) and left at room temperature for 5 min, and 0.2 mL of chloroform was added; the samples were then vigorously mixed for 10 s and allowed to stand for 2 min at room temperature. The samples were then centrifuged at 12,000 g for 15 min to separate the mixture into an aqueous phase that contained the RNA, which was separated into a new tube. Isopropanol (0.5 mL) was then added to the RNA, left at room temperature for 10 min, and then centrifuged for 10 min at 12,000 g and 4°C. The precipitates were converted to complementary DNA (cDNA) using superscript reverse transcriptase and analyzed by real-time polymerase chain reaction using SYBR green chemistry using the protocol recommended by the manufacturer (ROCHE). Bovine primer sequences were designed and ordered from Invitrogen (Table 1). The data were quantified by comparing them to a standard curve obtained from a set of serial dilutions (1:10, 1:100, and 1:1000) of a calibrator cDNA solution. Gene expression was normalized to that of the housekeeping gene GAPDH.
Mechanical testing
Tissues were subjected to a stress relaxation test using an Autograph control/measuring unit (Shimadzu Corp.). The equipment applied a constant rate of displacement with a 500 N load cell through a servo motor controlled by a computer. The thickness of the sample was set to the point at which the load cell detected a force of 10 mN. The specimen was then compressed at a speed of 10% specimen thickness, automatically stopped at a depth of 20% thickness, and held in that position for 10 min. The force and displacement over time were recorded by the Autograph, and the Young's Modulus was calculated from the linear portion of the equilibrium stress–strain curve.
Quantification of DNA content, GAG, and total collagen
Tissues were frozen, digested with papain (Sigma) at 60°C for 16 h, and then homogenized using a polytron machine. The sulfated GAG content was quantified using dimethyl methylene blue dye binding. DNA content of the papain digests was measured using 4, 6-diamidino-2-phenylindole (DAPI; Dojindo). Total collagen was measured using the Sircol Soluble Collagen Assay (Biocolor). All specimens were analyzed under a UV-VIS spectrophotometer (Parker Elmer) set at wavelengths of 620, 350, and 280 nm for GAG, DNA, and total collagen, respectively.
Histological analysis and immunostaining
Tissues were washed with PBS and then fixed in 4 mL of 10% formalin (Wako) per tissue. Next, the tissues were dehydrated through a graded series of ethanol, embedded in paraffin, sectioned at 5 μm thickness, and stained with toluidine blue and safranin-O to detect the anionic sulfated proteoglycans (PGs). Collagen type I (Fine Chemicals) and collagen type II (Fine Chemicals) were detected using immunohistochemical analysis, and phosphorylated p38 (Santa Cruz Biotechnology), JNK (Santa Cruz Biotechnology), and ERK2 (Santa Cruz Biotechnology) expression was detected using immunofluorescence. Briefly, the paraffin was removed through rinses with xylene (Wako) and ethanol (Wako), and then washed with PBS. Then, samples were soaked for 15 min in 0.005% proteinase (Sigma) in Tris-HCl and then washed with PBS. Hydrogen peroxide (3%) (Wako) in methanol (Wako) was used to quench the action of the proteinase and was then blocked with 1.3% normal goat serum (Rockland Immunochemicals, Inc.) in Tris-HCl (pH 7.6) for 45 min. Samples were then stained with the primary antibody (Fine Chemical) overnight, washed with PBS, and then the secondary antibody was added. Peroxide-conjugated streptavidin (DAKO) in 1% bovine serum albumin was added at room temperature for 45 min and then washed with PBS. DAB (Dojindo) in 0.05 M Tris-HCl and 30% hydrogen peroxide were added for 30 s, and the sections were finally stained with hematoxylin (Sigma) or DAPI before mounting.
Statistical analysis
All results were obtained from three independent experiments and presented as mean±standard error of the mean. Statistical analysis of data was done by performing an ANOVA followed by a series of Student's t-tests with a Bonferroni correction of n=7 for the preliminary ultrasound intensity experiments and n=4 for the experiments utilizing LIPUS and TGF-β3. A p-value of 0.05 or less was assumed statistically significant.
Results
The effect of LIPUS and TGF-β3 on the differentiation of scaffold-free dedifferentiated bovine articular chondrocytes was studied. Tissues were evaluated through histology and immunostaining, PG and total collagen protein production, cartilage-specific and hypertrophic-inducing gene expression, and mechanical properties.
Increased PG and total collagen protein production
The differences in the cartilage matrix assembly in the tissues were assessed by quantifying the GAG and total collagen content per unit of DNA relative to the control tissues. Preliminary experiments to determine the optimal frequency and intensity of LIPUS indicate that 55 mW/cm2 at 1 MHz resulted in the greatest increase in relative GAG/DNA (Fig. 2A) and total collagen/DNA (Fig. 2B), which was subsequently used for experiments with TGF-β3.
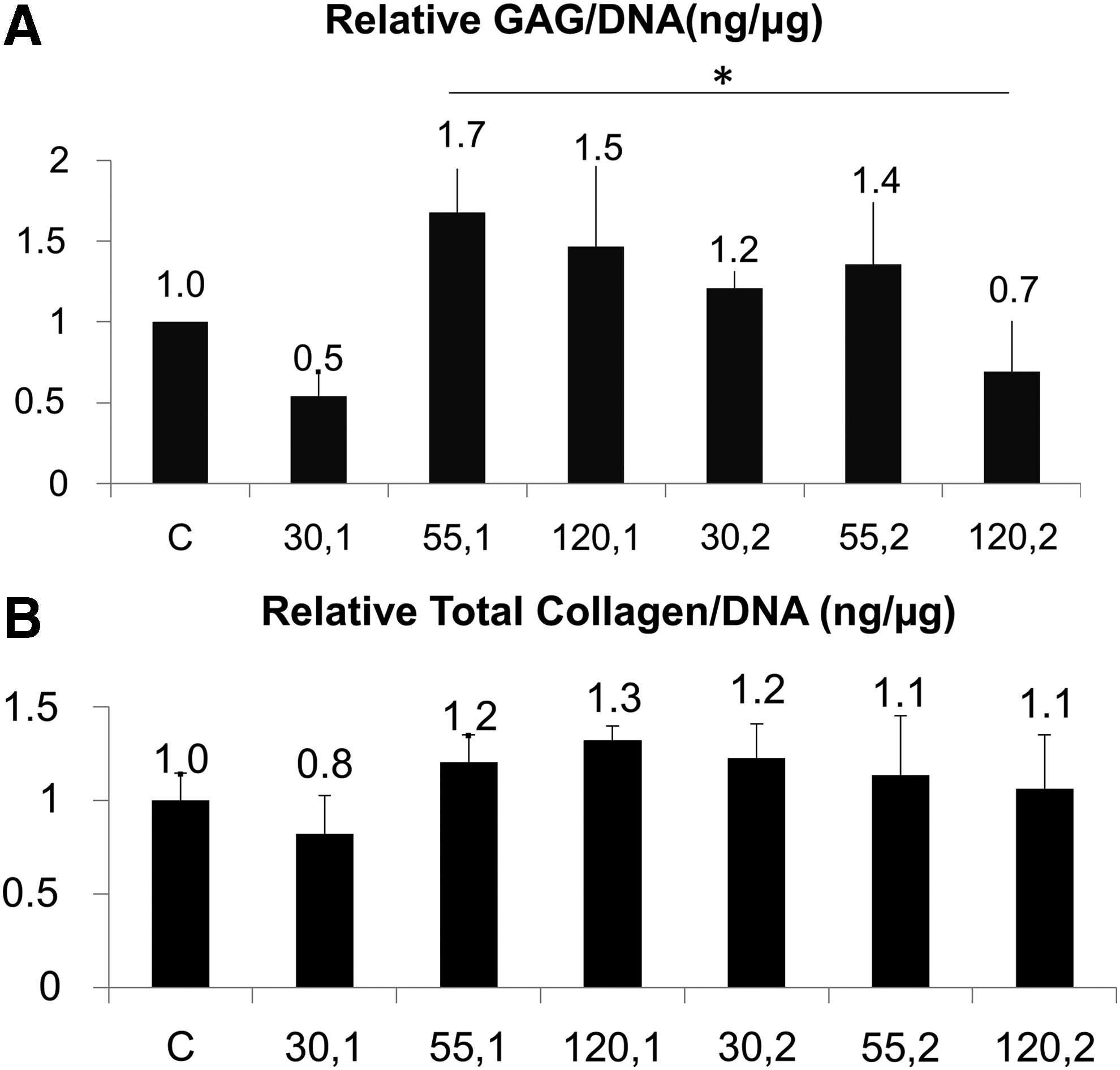
Relative amount of
Results show that the effect of mechanical and chemical stimulation significantly increased GAG/DNA by 2.15±0.12, 3.33±0.12, 3.39±0.05 times the control case for LIPUS, TGF-β3, and LIPUS+TGF-β3, respectively (Fig. 3A). The addition of LIPUS+TGF-β3 resulted in only a small increase of GAG/DNA over TGF-β3-treated tissues, but had a smaller margin of error. LIPUS+TGF-β3 resulted in the greatest total collagen ratio relative to control at 6.2±1.3 (Fig. 3B). TGF-β3 also had a greater increase in total collagen over LIPUS alone and the control case at 4.3±0.74; however, this increase could also be due to the extra type I collagen protein produced, as indicated by the immunostaining for collagen type I. The application of LIPUS on tissues effectively promoted proliferation of the chondrocytes by a factor of 2.51±0.63 and 2.63±0.47 for LIPUS and LIPUS+TGF-β3, respectively. DNA quantification indicates that DNA was increased by over twofold when chemical and mechanical stimulation was applied, indicating the positive effect of these stimuli on proliferation (Fig. 3C).
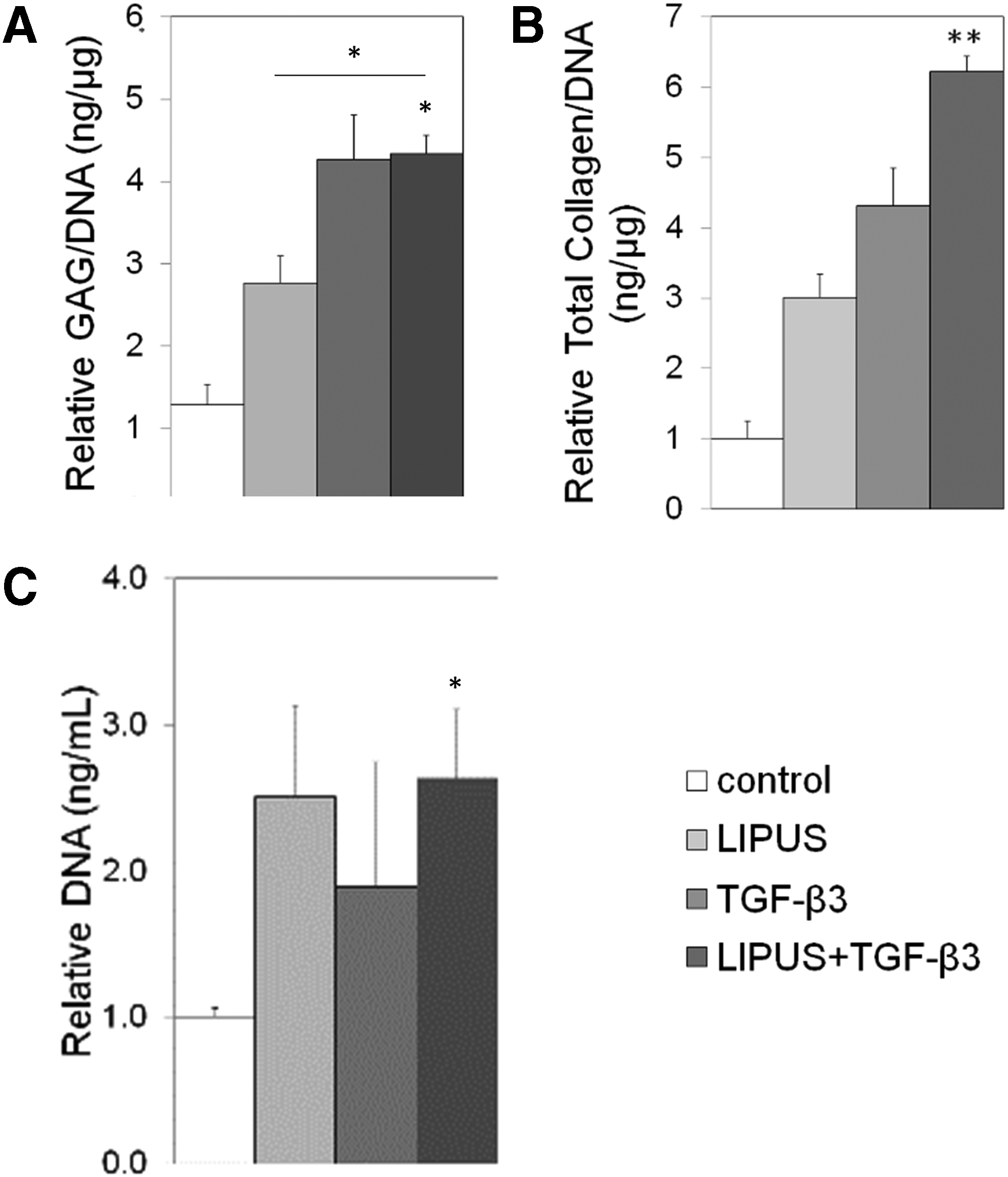
Relative amount of
Increased cartilage matrix production with LIPUS and TGF-β3 treatment
Staining for toluidine blue, safranin-O, and collagen type I and II was done to determine the relative amount and distribution of cartilage-specific matrix within the tissues. Histological staining showed that control and TGF-β3-treated tissues had large sections of necrosis at the center of the tissue. However, the addition of LIPUS to the tissues slightly remediated this and also promoted cartilage matrix development (Fig. 4D, H, L, P). TGF-β3-treated tissues had a stronger positive stain for PGs in the center of the tissue with a less intense stain at the edge with a clear border (Fig. 4C). LIPUS+TGF-β3-treated tissues showed a similar effect, but were more homogeneous (Fig. 4D). Collagen II was more positively expressed on the bottom of samples subjected to LIPUS and LIPUS+TGF-β3 (Fig 4N, P). Typically, more cells were concentrated in the center of the samples for tissues subjected to TGF-β3 and LIPUS+TGF-β3 (Fig. 4G, H). However, although the collagen II was most strongly stained in the case of LIPUS+TGF-β3-treated tissues, there was also a stronger stain for collagen I (Fig. 4P). A minimum of three samples were used to prepare paraffin blocks, and all control and TGF-β3 samples showed blank areas in the center of the scaffold. Testing for apoptosis using TUNEL staining was carried out on the samples, but no areas of apoptosis (data not shown) were detected, suggesting that these areas were most likely necrotic.
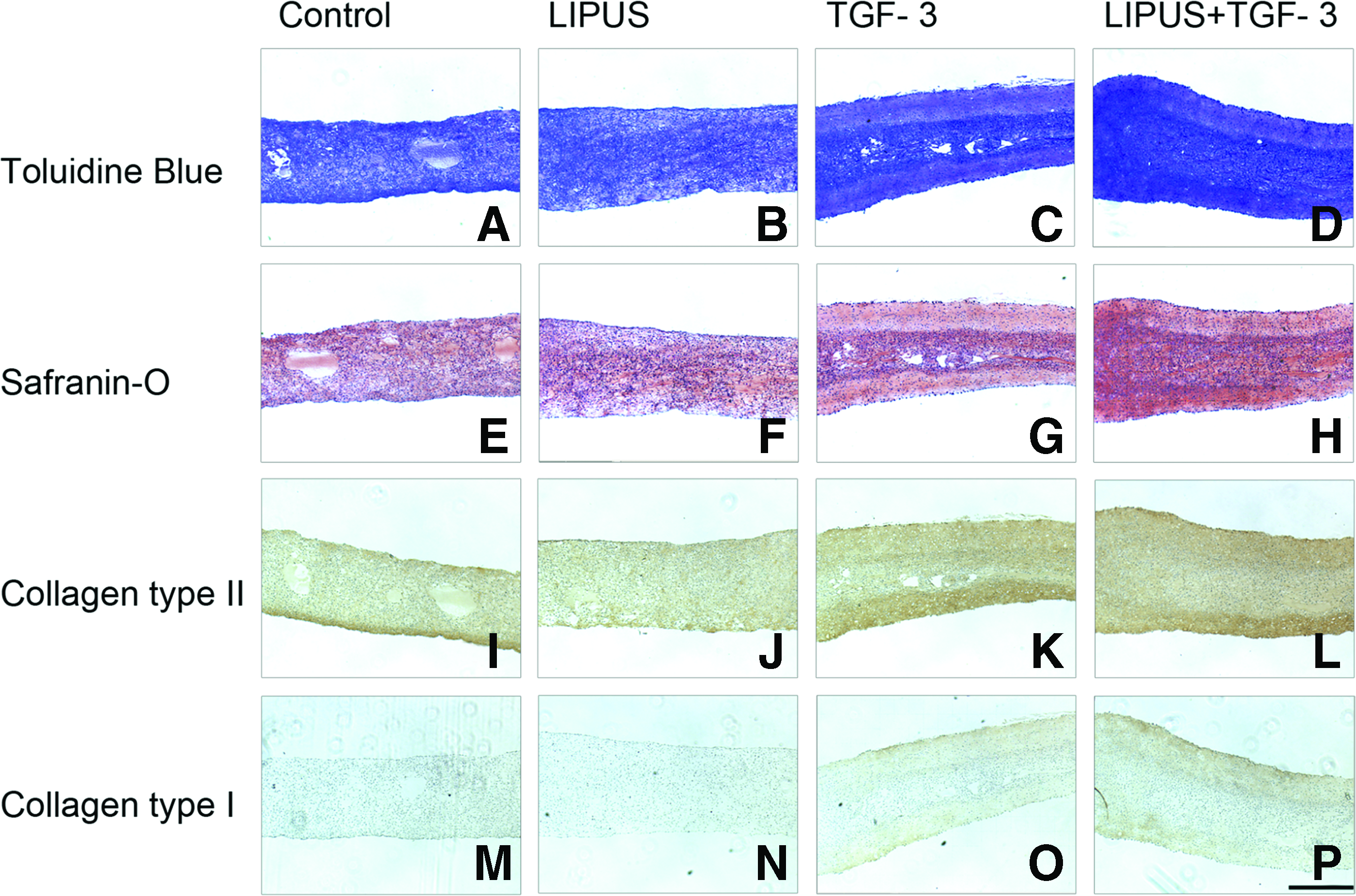
Comparison of matrix synthesis and localization in constructs exposed to LIPUS (55 mW/cm2) with or without TGF-β3. Toluidine blue stain for proteoglycan
Regulation of cartilage-specific gene expression, collagen type I, and collagen type X
The relative differences in gene expression between control and treated tissues for cartilage-specific genes (collagen type II and aggrecan) were determined using real-time polymerase chain reaction analysis, along with collagen type I and X, markers that indicate hypertrophy and OA development. Collagen type II and aggrecan upregulation was the highest for LIPUS+TGF-β3 constructs. The downregulation of collagen type I expression was greatest for LIPUS alone at 0.28±0.04 times the control value. Collagen type X gene expression was not significantly different between the four groups (Fig. 5).

Gene expression relative to the control case of tissues subjected to 55 mW/cm2 LIPUS for 20 min/day over 10 days with or without TGF-β3. All values at n=3 mean±STD, *p<0.05, **p<0.01.
Modulation of cartilage degrading proteinase gene expression
Collagen type II cleaving proteinases, MMP-13 and MMP-2, and aggrecan cleaving proteinases, MMP-3 and ADAMTS5, were measured to check for degradative enzyme production within the tissues. LIPUS significantly downregulated MMP-13 and MMP-2 to 0.27±0.08 and 0.02±0.01 relative to control, respectively; whereas treatment with TGF-β3 increased MMP-13 and MMP-2 gene expression by a factor of 1.4±0.38 and 2.8±1.5, respectively. LIPUS, TGF-β3, and TGF-β3+LIPUS stimulation downregulated MMP-3 and ADAMTS5 gene expression to 0.20±0.16, 0.03±0.02, 0.01±0.007 and 3.6e−3±1.9e−3, 3.9e−2±1.9e−3, 3.9e−3±3.5e−4, respectively (Fig. 5).
Increased mechanical properties and tissue growth
The Young's Modulus and tissue properties were analyzed to determine the effects of mechanical and chemical stimulation on the resulting mechanical properties of the tissue. The addition of TGF-β3 and LIPUS+TGF-β3 to the scaffold-free tissues resulted in a significant increase in thickness over the control case with the greatest increase in TGF-β3 alone (Fig. 6A).
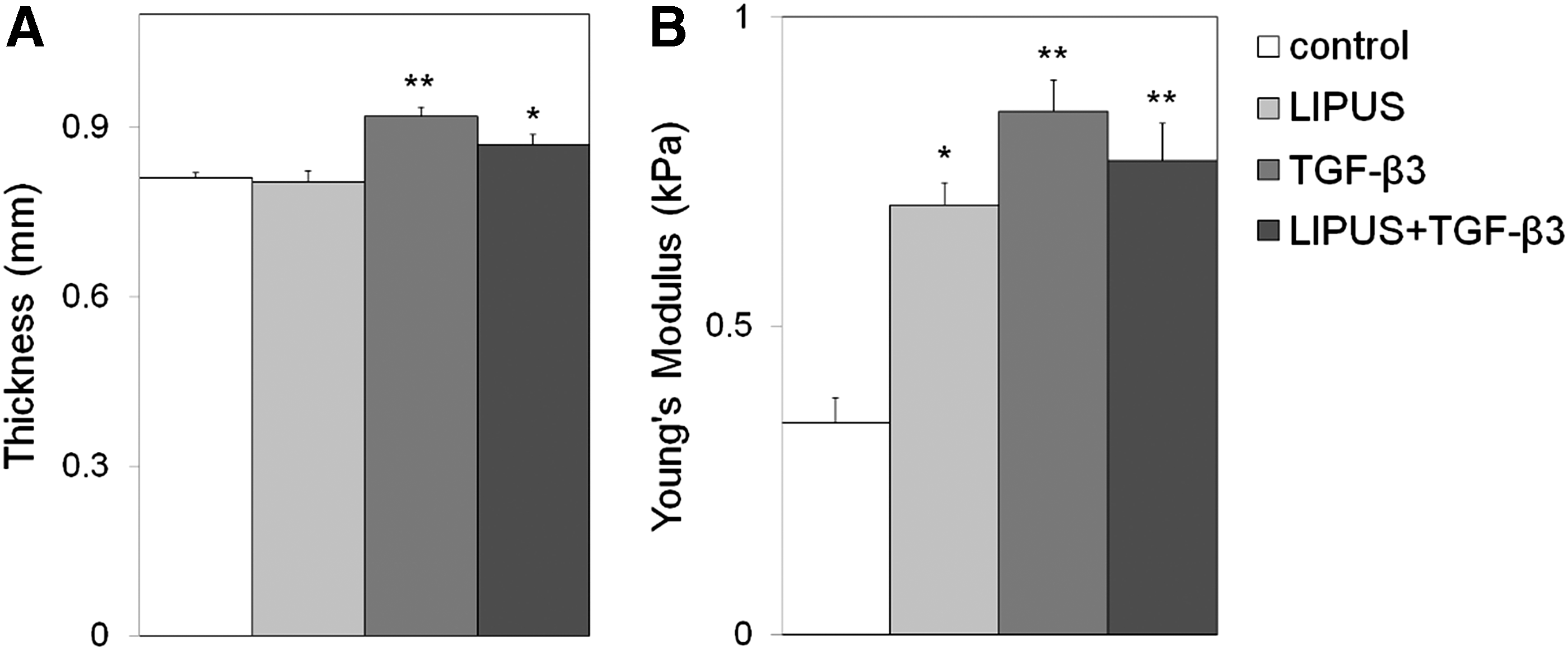
Results from unconfined compression test of tissues subjected to 55 mW/cm2 LIPUS for 20 min/day over 10 days with or without TGF-β3.
The Young's Modulus calculated from the compressive stress–relaxation curve was determined to be 343±40, 695±36, 847±51, 767±61 Pa for control, LIPUS, TGF-β3, LIPUS+TGF-β3, respectively (Fig. 6B), which corresponds to the increased PG and total collagen protein production measured in the samples. However, the Young's Modulus for the native cartilage is 380±120 kPa, which is an order of magnitude greater than our results, indicating that the mechanical properties of our tissues are still mechanically inferior to the native tissue.
Increased phosphorylated p38 signaling with TGF-β3 treatment
Immunofluorescent staining of phosphorylated p38 indicated an increase in the nuclear expression of p38 with TGF-β3 treatment and the absence of staining when LIPUS is added to TGF-β3 (Fig. 7). Total ERK2 was also increased with TGF-β3 treatment, although no nuclear-specific translocation was observed (Fig. 8). No changes in staining for JNK were shown in all groups (Fig. 9). No noticeable differences were noted for pSMAD2/3 and pSMAD1/5/8 staining (data not shown), indicating that only p38 signaling was activated with the application of chemical stimulation.
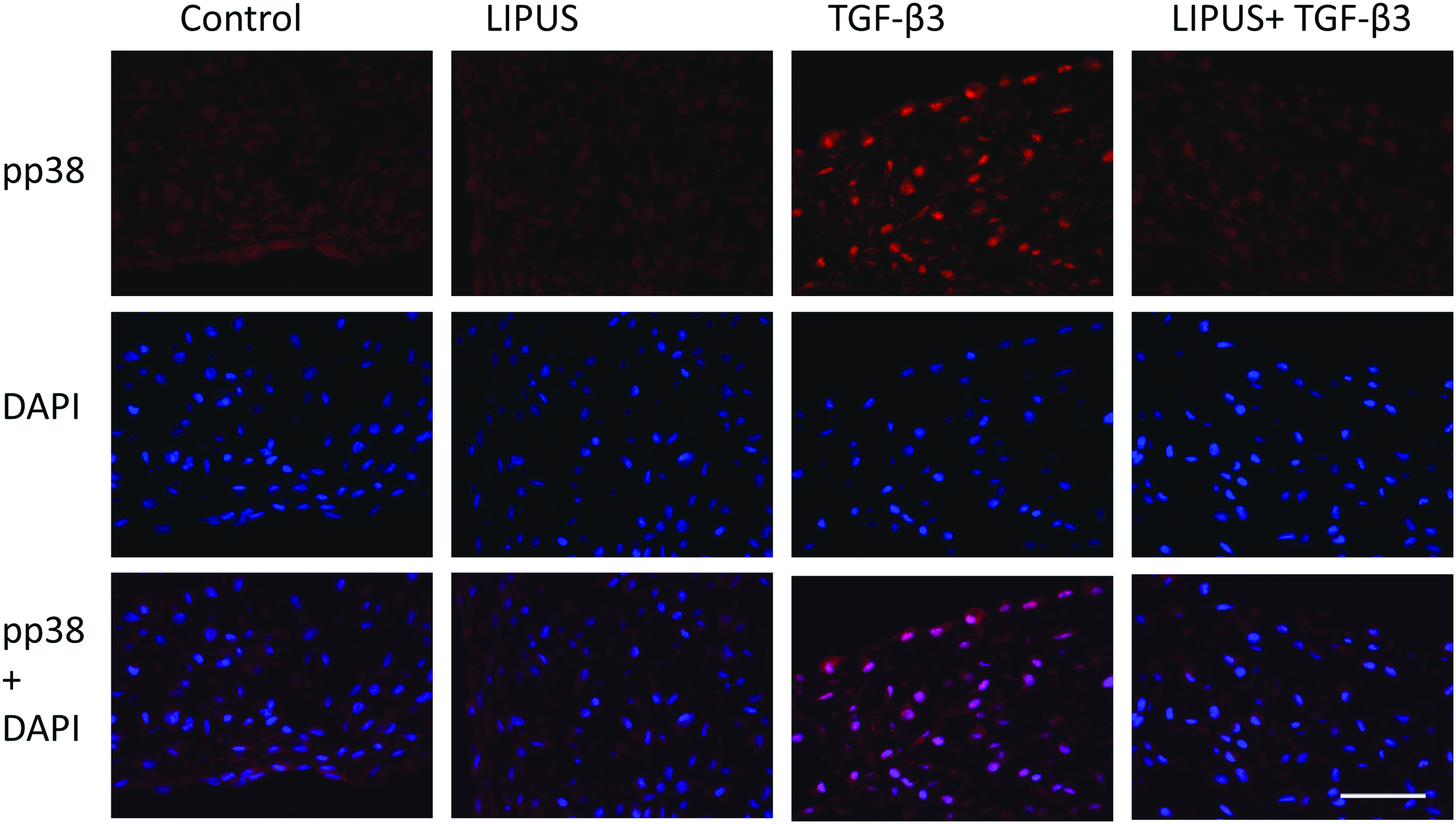
Immunofluorescent staining of phosphorylated p38, DAPI, and the overlay of both. Scale bar=100 μm. Color images available online at
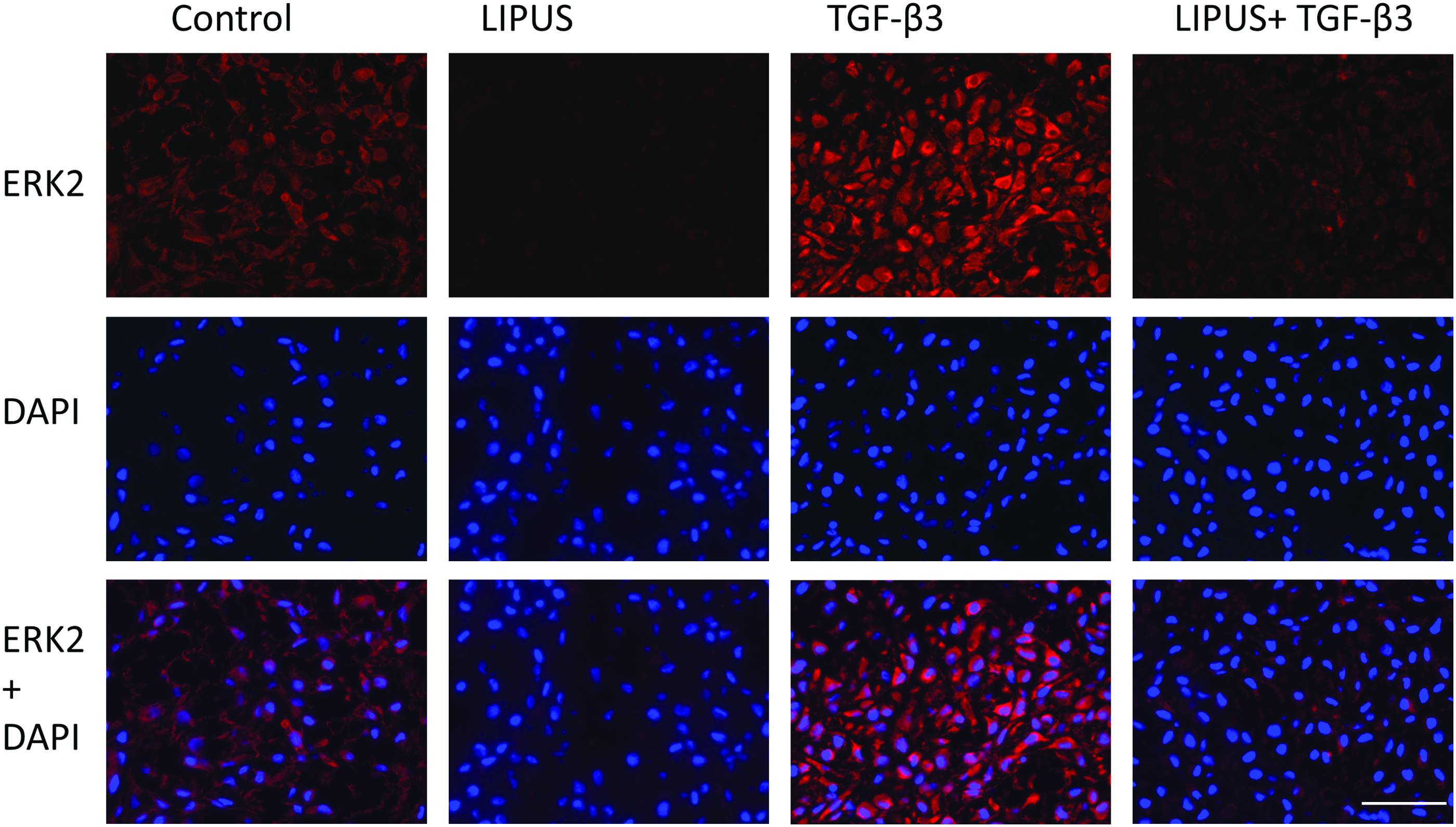
Immunofluorescent staining of ERK2, DAPI, and the overlay of both. Scale bar=100 μm. Color images available online at

Immunofluorescent staining of JNK, DAPI, and the overlay of both. Scale bar=100 μm. Color images available online at
Discussion
In this study, we present how LIPUS inhibited TGF-β3-induced hypertrophic gene expression and signaling, and further enhanced the differentiation of dedifferentiated chondrocytes. In our system, TGF-β3 upregulated cartilage-specific gene expression and increased GAG and collagen protein production; however, there was also the upregulation of hypertrophic markers of collagen type X and MMP13, and the phosphorylation of p38, indicating that TGF-β3 could not suppress terminal differentiation. Conversely, the addition of LIPUS to TGF-β3-treated tissues significantly downregulated some hypertrophic gene expression and cellular signaling and enhanced collagen type 2 protein production, suggesting that LIPUS can modulate the effects of TGF-β3 toward a chondrogenic lineage in dedifferentiated articular bovine chondrocytes.
TGF-β, which is considered an anabolic growth factor for cartilage, has been shown to regulate chondrocyte differentiation and growth 24 and can accelerate fracture healing by promoting chondrogenesis and chondrocyte proliferation in an in vitro scaffold-free culture.8,21,25 However, the effects of TGF-β on hypertrophic development are critically dependent on the culture conditions and the stage of differentiation of the target cell. With immature murine chondrocytes, it has been described that TGF-β induces proliferation and inhibits hypertrophy. 26 Oppositely, TGF-β induces the upregulation of collagen type X gene expression in bone marrow hMSCs. 27 In another study on long-term pellet culture of hMSCs subjected to TGF-β1, β3-supplemented chondrogenic medium, or hypertrophic medium for 2 or 4 weeks, all samples had a positive collagen type II immunostaining with collagen I detected in the periphery, consistent with our results, along with positive collagen type X in all samples. 28 In nondynamic culture of samples with TGF-β3, the concentration of the growth factor that cells are exposed to changes depending on the location of the cell within the scaffold. We predict that this difference in concentration resulted in the clear border for cartilage protein development. The same group also found no noticeable differences in the cell morphology between the hypertrophic and control group after 28 days. TGF-β may potentially only promote hypertrophic development with cells that have reached a mature state. Other variables such as growth factor concentration have also been shown to be important. The commonly used concentration of 10 ng/mL of TGF-β supplementation was implemented in our study, but differences in MMP-13 expression were shown to be dependent on the concentration of TGF-β applied to chondrocytes found in nonlesional areas of the cartilage. 18 Cartilage defects in rabbit knee joints filled with 4% HA gels mixed with 250 ng of TGF-β3 had no beneficial effect on cartilage regeneration 29 ; whereas overexposure to the growth factor through injections resulted in the formation of an OA-like tissue in murine knee joints. 30
Although a concentration of 10 ng/mL of TGF-β to the pellet culture of hMSCs induced chondrogenesis, 21 the surface area to volume ratio is lower in the pellet versus our disc-shaped tissue; therefore, it is possible that the exposure of exogenous TGF-β in our study was too high, resulting in the upregulation of hypertrophic gene expression.
The signaling pathway activated by TGF-β largely influences the cellular response. Signaling through SMAD2/3 is necessary for the regulation of chondrocyte differentiation and inhibition of terminal differentiation. 31 However, TGF-β signaling is dynamic and can shift from SMAD3-mediated hypertrophic repression to a dominant SMAD1/5/8 or p38-mediated promotion of chondrocyte terminal differentiation.32–34 In SMAD3-deficient mouse chondrocytes and ATDC5 chondroprogenitor cells, TGF-β-treatment repressed the expression of MMP-13 mRNA and protein within 8 h, but was later induced at 26 and 48 h through p38 activation. 32 Although there were no apparent changes in the fluorescent staining of phosphorylated SMAD2/3 between all samples (data not shown), the application of LIPUS effectively prevented the phosphorylation of p38 that was initiated by the application of exogenous TGF-β3 (Fig. 7).
To remediate the hypertrophic effects of TGF-β3 in our study, we implemented mechanical stimulation through LIPUS. The positive effects of LIPUS on TGF-β3 treatment through the cartilage-specific gene and protein production have been extensively studied using pellet or aggregate models.19,21,27 The application of LIPUS to rabbit legs has also been shown to delay the degradation of collagen type II and PG during the early stages of OA. 34 Furthermore, low-intensity ultrasound treatment on rabbit MSCs in alginate or polyglycolic acid scaffolds led to a downregulation in hypertrophic gene expression in vitro.35,36 In our study, we also showed that LIPUS effectively downregulated the TGF-β3-induced cartilage-degrading gene expression, but these effects of LIPUS were only found in conjunction with the application of TGF-β3, which is similar to the results by Ebisawa et al. using MSCs in pellet form. Conversely, a study with rabbit MSCs in alginate subjected to continuous ultrasound for 15 min/day showed the same effect with low-intensity ultrasound alone. 21 These discrepancies may be due to experimental variables such as cell type and culture conditions. The mechanisms in which LIPUS modulates cell function and protein synthesis have yet to be determined, but some proposed mechanisms are acoustic streaming, caused by the compression of microbubbles or cavitation, which changes the cell membrane permeability and subsequently modifies intracellular signals for gene expression through changes in ion or protein transport, 20 and mechanical pressure at the cell surface, which could activate stretch-activated cation channels. 35 LIPUS could inhibit cartilage matrix turnover by augmenting TIMP expression, but this was only observed with the cotreatment of TGF-β3 in alginate cultures of MSCs. 36 As ultrasonic energy passes through a tissue, it is absorbed at a rate proportional to the density of the tissue, which results in the generation of heat. 37 Detrimental effects, such as cell death, due to ultrasound on tissues in vitro have been generally observed at temperatures of 39°C–43°C. 38 Enzymes such as MMP-1 and collagenase are also extremely sensitive to changes in temperature. 39 In surgical and therapeutic ultrasound (1–300 W/cm2), a large amount of heat is generated when applied to tissues; however, LIPUS (<100 mW/cm2) is regarded as nonthermogenic and nondestructive. 37 The actual temperature within the tissues themselves as the LIPUS passes through them may have changed due to the difference in the density, but there was no change in the temperature of the medium noted before and after LIPUS treatment, indicating that potentially the same occurred in the tissues.
It is commonly thought that TGF-β3 and LIPUS have different signaling pathways: SMADs and mechanotransduction mediated by integrins, respectively. 36 However, mechanical stimulation such as dynamic compression has been shown to modulate TGF-β3 signaling by potentially increasing the phosphorylation of the SMAD2/3 proteins and/or their translocation to the cell nucleus, 39 suggesting that there may be an interconnection between pathways activated by chemical and mechanical stimuli. In addition, LIPUS treatment on myogenic C2C12 cells could divert their differentiation into an osteoblast and chondroblast lineage through the activation of ERK1/2 and/or p38 MAPK. 40 Potentially, in our system, LIPUS may have shifted the balance from TGF-β-activation of p38 to SMAD3, to enhance the anabolic effects and reduce the catabolic effects of the growth factor supplementation. In addition, the added TGF-β3 may have better penetrated the entire scaffold-free tissue when LIPUS was applied due to the acoustic streaming phenomena, leading to better perfusion of nutrients and wastes, thus altering the cell signaling pathways and also preventing central necrosis, as seen in both control and TGF-β3 tissue cultures. The influence of mechanical loading also depends on the TGF-β concentration in the culture medium, with a much stronger effect on chondrogenic gene expression observed at lower concentrations, 41 and therefore, additional optimization of TGF-β3 concentration may enhance chondrogenesis or prevent hypertrophy all together.
For clinical studies, the most relevant cell source would be adult human chondrocytes; however, these cells require repeated expansion on a 2D surface to be sufficiently expanded. Although the cells in this study were used at passage 2, previous research indicates that human chondrocytes can effectively be redifferentiated through alginate beads up to passage 4 42 ; therefore, in clinical studies, our system may potentially be applicable to human chondrocytes at higher passage numbers. Previous studies have also indicated that pretreatment of implants with LIPUS resulted in less blood invasion and osteogenic phenotypes and more cartilage ECM development, compared to the constructs not treated with LIPUS. 36
The present study has shown that LIPUS is a simple and cost-friendly way to inhibit TGF-β3-induced hypertrophic development and differentiate dedifferentiated chondrocytes toward a chondrogenic lineage in a scaffold-free system with versatile shape control. By inhibiting hypertrophy in the tissue before implantation, the tissue could potentially better develop into articular cartilage instead of fibrocartilage, as commonly seen in damaged cartilage. Therefore, the LIPUS system in combination with TGF-β3 treatment can be an effective tool in clinical studies for cartilage tissue engineering.
Footnotes
Acknowledgments
This work was supported by the program of Next Generation on World-Leading Researchers (NEXT), Japan Society for the Promotion of Science (JSPS) and, in part, by the Global COE Program (GCOE), Medical System Innovation on Multidisciplinary Integration, MEXT, Japan. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the article.
Disclosure Statement
No competing financial interests exist.