
Abstract
Living tissue-engineered heart valves (TEHV) would be a major benefit for children who require a replacement with the capacity for growth and biological integration. A persistent challenge for TEHV is accessible human cell source(s) that can mimic native valve cell phenotypes and matrix remodeling characteristics that are essential for long-term function. Mesenchymal stem cells derived from bone marrow (BMMSC) or adipose tissue (ADMSC) are intriguing cell sources for TEHV, but they have not been compared with pediatric human aortic valve interstitial cells (pHAVIC) in relevant 3D environments. In this study, we compared the spontaneous and induced multipotency of ADMSC and BMMSC with that of pHAVIC using different induction media within three-dimensional (3D) bioactive hybrid hydrogels with material modulus comparable to that of aortic heart valve leaflets. pHAVIC possessed some multi-lineage differentiation capacity in response to induction media, but limited to the earliest stages and much less potent than either ADMSC or BMMSC. ADMSC expressed cell phenotype markers more similar to pHAVIC when conditioned in basic fibroblast growth factor (bFGF) containing HAVIC growth medium, while BMMSC generally expressed similar extracellular matrix remodeling characteristics to pHAVIC. Finally, we covalently attached bFGF to PEG monoacrylate linkers and further covalently immobilized in the 3D hybrid hydrogels. Immobilized bFGF upregulated vimentin expression and promoted the fibroblastic differentiation of pHAVIC, ADMSC, and BMMSC. These findings suggest that stem cells retain a heightened capacity for osteogenic differentiation in 3D culture, but can be shifted toward fibroblast differentiation through matrix tethering of bFGF. Such a strategy is likely important for utilizing stem cell sources in heart valve tissue engineering applications.
Introduction
H
One of the challenges for tissue-engineered heart valve (TEHV) is to find suitable cell sources. Early studies demonstrated that allogenic cell sources caused an acute inflammatory response even in the presence of immunosuppressant treatment. 6 Because valve cells are nonsacrificial, TEHV initially involved the isolation of autologous cells from accessible vascular sources, including carotid artery,7,8 peripheral arteries, 9 and saphenous veins. 10 The cells were mixed cellular populations with fibroblasts and smooth muscle cells, sometimes with endothelial cells. 11 Although the isolation process is technically simple and these cells show substantial tissue formation capacity in vitro, the use of vascular-derived cells requires the sacrifice of intact vascular donor tissues. In addition, the hybrid cells have different phenotypes compared with valve interstitial cells (VIC) and may lead to the stenosis of the valve conduits and result in final failure. More recently, postnatal multipotent stem cells such as mesenchymal stem cells (MSC) have been evaluated as cell sources for TEHV due to their self-renewal capacity and potential for differentiation into several lineages.12–14 Among various MSC sources, bone marrow-derived MSC (BMMSC) and adipose-derived MSC (ADMSC) are prime candidates for TEHV due to their greater availability and popularity.15–19 However, a few studies have compared phenotype and extracellular matrix (ECM) deposition characteristics between MSC and VIC, especially pediatric VIC. VIC are a heterogeneous population of quiescent fibroblasts, a small population (∼20% for neonatal, ∼6% for child and ∼2.5% for adult valves) of myofibroblasts and mesenchymal progenitor cells.20,21 Pediatric valves have less organized collagen fiber, lower elastin content, and higher glycosaminoglycans (GAG).20,22 Latif et al. demonstrated that both BMMSC and adult VIC expressed fibroblast surface antigen (FSA), α-smooth muscle actin (αSMA), vimentin, and CD44. 23 ECM components secreted by both cell types showed similar levels and patterns of staining, though expression of elastin was not detected. Colazzo et al. compared ADMSC with BMMSC and VIC for collagen and elastin deposition in both static and cyclic stretch condition. 19 All the aforementioned comparisons were based on qualitative assessments with cells cultured on two-dimensional (2D) substrates. It is well known that cells exhibit very different behaviors when cultured in three-dimensional (3D) environments, and these environments are better mimics of the physiological condition. 24
Cultured adult VIC exhibit multi-lineage differentiation potential, for example, positive for von Kossa, Alcian blue, and Oil red staining after conditioning in induction media. 25 During aortic valve disease progression, both pediatric and adult diseased aortic valves express markers of chondrogenic cells, while adult diseased aortic valves express more mature osteogenic markers. 26 However, it is unclear what potency pediatric VIC possess, and controlling stable MSC differentiation toward pediatric VIC is important for long-term success of TEHV for pediatric applications.
In this study, we compared the multi-lineage differentiation capacity of human ADMSC and BMMSC with pediatric human aortic VIC (pHAVIC). We evaluated their spontaneous differentiation in physiological condition and pathological response in different pathological conditions. We also investigated and compared the feasibility to induce MSC differentiation toward pHAVIC within a 3D physiological microenvironment. We identified several differences in baseline and induced valvular phenotype expression and ECM remodeling characteristics between pHAVIC and differentiated MSC. We further tested the hypothesis that basic fibroblast growth factor (bFGF) influenced MSC differentiation toward pHAVIC via media supplementation and direct tethering of bFGF to the 3D matrix.
Materials and Methods
Polymer modification and hydrogel preparation
Photocrosslinkable hyaluronic acid (HA, Novozymes, ∼1200 kDa) and gelatin (Gel, from bovine skin; Sigma) were synthesized as previously reported 27 through the reaction of methacrylic anhydride (Sigma) with 0.5% HA or 10% Gel in deionized water. The degree of methacrylation was approximately estimated to be 22.5% for methacrylated HA (Me-HA) and 61.1% for methacrylated Gel (Me-Gel) based on the 1 H NMR spectroscopy.
For hydrogel preparation, Me-HA (0.5%w/v) and Me-Gel (1%w/v) were dissolved in cell culture medium with cell suspension (2.5×106 cells/mL) and 0.05% w/v 2-hydroxy-1(4-(hydroxyethox)pheny)-2-methyl-1-propanone (Irgacure 2959; CIBA Chemicals). The gel precursor was transferred into silicon molds (ø8 mm, 1 mm thickness) and subsequently exposed to 365 nm UV light (EN-280L, Spectroline, 2.0 mW/cm2) for 5 min. The cell-hydrogel discs were washed with PBS and maintained at 37°C and 5% CO2.
Cell isolation and cell culture
pHAVIC were isolated from the aortic valve leaflets of the donor heart from a 12-year-old patient undergoing cardiac transplant for a myocardial contractility deficiency. These valve leaflets exhibited normal dynamics via clinical echo, absence of calcific deposits, and no thickened lesions that are characteristic of aortic valve disease that is not present in valves. 28 Tissue was procured with consent as approved by the Institutional Review Board of Weill-Cornell Medical College in New York City as previously described. 29 Cells were cultured in HAVIC growth medium (HGM) containing MCDB131 medium (Sigma), 10% fetal bovine serum (FBS; Invitrogen), 1% penicillin/streptomycin (P/S; Invitrogen), 0.25 μg/L recombinant human fibroblast growth factor basic (rhbFGF; PeproTech), and 5 μg/L recombinant human epidermal growth factor (rhEGF; Invitrogen).28,30,31 Cells were used at passages 4–8. Established primary human ADMSC and BMMSC were purchased from Lonza and cultured in stem cell growth medium (GM) containing DMEM/F12 medium (Invitrogen), 10% FBS, and 1% P/S.
Multi-lineage differentiation
The cell-hydrogel constructs were maintained in their respective growth medium (MCDB medium for pHAVIC and DMEM/F12 medium for ADMSC and BMMSC) for 7 days and were then induced into osteogenic, adipogenic, and chondrogenic differentiation for another 21 days. The osteogenic medium (OGM) consisted of GM, 100 nM dexamethasone (Sigma), 10 mM β-glycerophosphate (Sigma), and 50 μM ascorbic acid (Sigma). 32 For adipogenic differentiation, the constructs were exposed to a cycle of 2 days of adipogenic induction medium (AGM containing GM, 1 μM dexamethasone, 200 μM indomethacin, 10 μg/mL recombinant human insulin, and 0.5 mM 3-isobutyl-1-methyl-xanthine; Sigma) and then 1 day of adipogenic maintenance medium (GM, 10 μg/mL recombinant human insulin) as previously described. 33 For chondrogenic differentiation, the encapsulated cells were conditioned in chondrogenic differentiation medium (CGM) consisting of GM, 100 nM dexamethasone, 0.2 mM ascorbic acid, 1 mM sodium pyruvate (Sigma), 1x insulin-transferrin-selenite (ITS+Premix; BD Biosciences), and 10 ng/mL transforming growth factor β1 (Sigma). 34 For induction of MSC toward VIC phenotypes, the encapsulated ADMSC or BMMSC were conditioned in GM for 7 days and then cultured in HGM for another 21 days.
Cell viability
The viability and circularity of encapsulated cells was determined using Live/Dead assay (Invitrogen) after 28 days of culture as previously described, 35 and fluorescence images were obtained using a confocal laser scanning microscope (CLSM, LSM 710; Carl Zeiss).
Alkaline phosphatase staining and activity
Alkaline phosphatase (ALP) staining and activity quantification was performed after a 28 day culture by using the ALP leukocyte kit (Sigma) according to the manufacturer's protocol. The cells within the hydrogels were lysed by adding 500 μL lysis buffer containing 0.1% (v/v) Triton X-100, 1 mM MgCl2, and 20 mM Tris. A 50 μL lysate was mixed with 200 μL ALP substrate solution containing p-nitrophenyl phosphate (pNPP) (Sigma) at 37°C for 10 min. The reaction was stopped by the addition of 50 μL of 3 N NaOH and then the production of p-nitrophenol in the presence of ALPase was measured by monitoring the absorbance of the solution at a wavelength of 405 nm using a microplate reader (Bio-Tek Instruments). The total protein content was determined using BCA assay kit (Pierce) with bovine serum albumin (BSA) as a standard, and the ALP activity was expressed as μmol of p-nitrophenol formation per minute per milligram of total proteins (μmol/min/mg protein).
Alizarin red S staining and quantification of calcium deposition
Calcium deposition was examined by using Alizarin red S (Sigma) staining. The cell-laden hydrogels were fixed with 4% paraformaldehyde and stained with Alizarin red S dye (pH=4.0–4.3). The hydrogels were washed several times, and the stained samples were imaged on Zeiss Discovery v20 stereomicroscope (Spectra Services, Inc.). Bound Alizarin red S dye was then released from the gels using 10% acetic acid, followed by neutralization with 10% ammonium hydroxide. 36 The absorbance of the dye solution was measured at 405 nm.
Oil red O staining and quantification of lipid accumulation
For Oil red O staining, the constructs were fixed in 4% paraformaldehyde and incubated in Oil red O solution (0.5% in propylene glycol; Sigma) for 10 min at room temperature. The stained hydrogels were imaged on Zeiss Discovery v20 stereo microscope after rinsing in 60% isopropyl alcohol and repeated washings with distilled water. Quantification of lipid accumulation was achieved by extracting Oil red O from stained cells with isopropanol and measuring the OD of the extract at 510 nm.
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA was extracted from cell-laden hydrogels using QIA-Shredder and RNeasy mini-kits (QIAgen) according to the manufacturer's instructions. Thirty nanograms of total RNA was synthesized into first-strand cDNA in a 20 μL reaction using iScript cDNA synthesis kit (BioRad Laboratories). Real-time polymerase chain reaction (PCR) analysis was performed in a CFX Connect Real-Time PCR detection system (Bio-Rad) using SsoAdvanced SYBR Green Supermix (Bio-Rad). All primers used in this study are listed in Supplementary Table S1 (Supplementary Data are available online at
ECM components analysis
Dimethylmethylene blue assay was performed to measure the sulfated GAG production in the hydrogels after a 7, 14, and 28 day culture. 28 The constructs (n=4–5 per condition) were digested with 300 μg/mL papain in 50 mM phosphate buffer (pH 6.5), containing 5 mM cysteine and 5 mM EDTA for 16 h at 60°C. GAG concentration was calculated by calibrating against a standard curve obtained with shark chondroitin sulfate (Sigma). To assess the biosynthetic activity of the cells, results of GAG were expressed as the ratio of GAG amount to DNA amount, which was assessed via the PicoGreen double-strand DNA assay (Invitrogen). The total collagen content was determined using the hydroxyproline assay in which hydroxyproline makes up 12.5% of collagen.37,38 The samples were hydrolyzed by 4.8 N HCl at 100°C for 3 h, the dried hydrolyzate was treated with 0.056 M chloramine T reagent, and the oxidation progressed for 25 min at room temperature. The Ehrlich's aldehyde reagent was reacted with samples at 65°C for 20 min to generate chromospheres. The amount of hydroxyproline was measured with the microplate reader at 550 nm (n=5). The total elastin content in the cell-laden hydrogels (n=4–6) was measured using the colorimetric Fastin elastin assay kit (Biocolor Ltd.), following the manufacturer's instructions. Results for collagen and elastin contents were normalized using the wet weight of the constructs and expressed in units of μg/g.
Immunohistochemistry staining
For immunohistochemical staining, cell-laden hydrogels were fixed in 4% paraformaldehyde, permeabilized in 0.2% Trion X-100, and blocked with 1% BSA overnight at 4°C. Hydrogels were treated with primary antibodies to vimentin (1:100, mouse monoclonal anti-vimentin; Invitrogen) and monoclonal anti-αSMA-Cy3 antibody (1:200; Sigma), or Sox9 (1:100; Abcam) overnight at 4°C. Secondary fluorescent antibodies were incubated for 2 h, and nuclear counterstaining (via Draq 5, 1: 1000; Biostatus) was performed for 30 min at room temperature. To visualize actin cytoskeletal filaments, the hydrogels were stained by Alexa Fluor 488 conjugated phalloidin (1:40; Invitrogen). The stained samples were imaged with Zeiss 710 CLSM.
Synthesis and characterization of the PEG-FGF conjugate
bFGF was conjugated to PEG by adding acrylate-PEG-succinimidyl carboxymethyl (ACRYL-PEG-SCM, 3400 Da; Laysan Bio, Inc.) to bFGF in a 200:1 molar in 50 mM sodium bicarbonate (pH 8.5) under sterile conditions for 4 days at 4°C. The PEG-FGF solution was then lyophilized under sterile condition and stored at −80°C. Western blot was used to confirm the conjugation of PEG-FGF.
Western blot
For western blotting, the cell-hydrogel constructs were washed twice with ice-cold PBS, flash frozen in liquid nitrogen, and lysed in hot laemmli buffer. The lysates were sonicated and microcentrifuged, and the protein concentrations were determined by Pierce 660 nm Protein Assay. The growth factor solution or lysates were loaded into a 4–15% gradient precast gel (Bio-Rad) and transferred to nitrocellulose membrane (Thermo Scientific). The membranes were blocked in Odyssey Blocking Buffer (Li-Cor) and incubated in monoclonal mouse anti-recombinant human bFGF (PeproTech) (1:2000), monoclonal mouse anti-vimentin antibody (Sigma) (1:2000), and mouse anti-human GAPDH (Invitrogen) (1:10,000), respectively. Blots were imaged using the Odyssey Infrared system (Li-Cor) after incubation in secondary antibodies. Semi-quatitative analysis of western blot was conducted by comparing the band intensity of vimentin to GAPDH using ImageJ as previously described.29,39
Statistical analysis
All quantitative data are expressed as the mean±standard deviation (SD). Statistical analysis was performed using ANOVA with Scheffé post hoc tests for pairwise comparisns. A value of p<0.05 was considered statistically significant.
Results
Cell viability and morphology within 3D hybrid hydrogels
Me-HA and Me-Gel were first synthesized by methacrylation of hydroxyl groups and primary amine groups, respectively, using methacrylic anhydride. Figure 1 presents cell viability and cell circularity of encapsulated pHAVIC, ADMSC, and BMMSC within the hybrid hydrogels after a 14 and 28 day culture in their GM. Generally, the cell viability was greater than 85% for all the cell types throughout 28 day culture (Fig. 1A, B). ADMSC had significantly higher cell viability compared with BMMSC and pHAVIC (Fig. 1B), supporting that they were more resistant to UV exposure during the encapsulation and photocrosslinking process. pHAVIC and ADMSC spread more than BMMSC over time in 3D hydrogel culture, and with lower circularity (Fig. 1A, C). HGM did not affect cell viability of ADMSC and BMMSC compared with GM but increased cell spreading for both ADMSC and BMMSC at day 14 and enhanced cell spreading for BMMSC at day 28. pHAVIC had the greatest spreading (lowest circularity) morphology after a 28 day culture.

Live/Dead assay for encapsulated pediatric human aortic valve interstitial cells (pHAVIC), adipose-derived mesenchymal stem cells (ADMSC), and bone marrow-derived mesenchymal stem cells (BMMSC) within hybrid hydrogels conditioned in HAVIC growth medium (HGM) or stem cell growth medium (GM) for 14 and 28 days.
Multi-lineage differentiation capacity of pHAVIC, ADMSC, and BMMSC
To examine and compare the multi-lineage differentiation potential of pHAVIC, ADMSC, and BMMSC, we cultured the cell-laden hydrogels for 7 days in GM, followed by treatment with osteogenic, adipogenic, and chondrogenic differentiation medium for a period of 3 weeks. Controls were maintained in HGM (pHAVIC, ADMSC, and BMMSC) or GM (ADMSC and BMMSC).
Encapsulated pHAVIC were negative to ALP staining as expected (Fig. 2A), and they had the lowest ALP activity (Fig. 2B). Unexpectedly, both ADMSC and BMMSC conditioned in GM and HGM were stained positively for ALP (Fig. 2A). We next tested whether the 3D hydrogel culture environment itself induced ALP expression. We therefore compared cell response with HGM or GM media in 2D culture for 7 days. We found that ADMSC and BMMSC expressed ALP in both baseline media conditions (Supplementary Fig. S1). Osteogenic supplementation induced osteogenic differentiation of pHAVIC, ADMSC, and BMMSC with a significant increase in ALP expression and activity (Fig. 2A, B). ADMSC and BMMSC showed significantly higher ALP activity compared with pHAVIC in OGM, with BMMSC having the highest ALP activity.
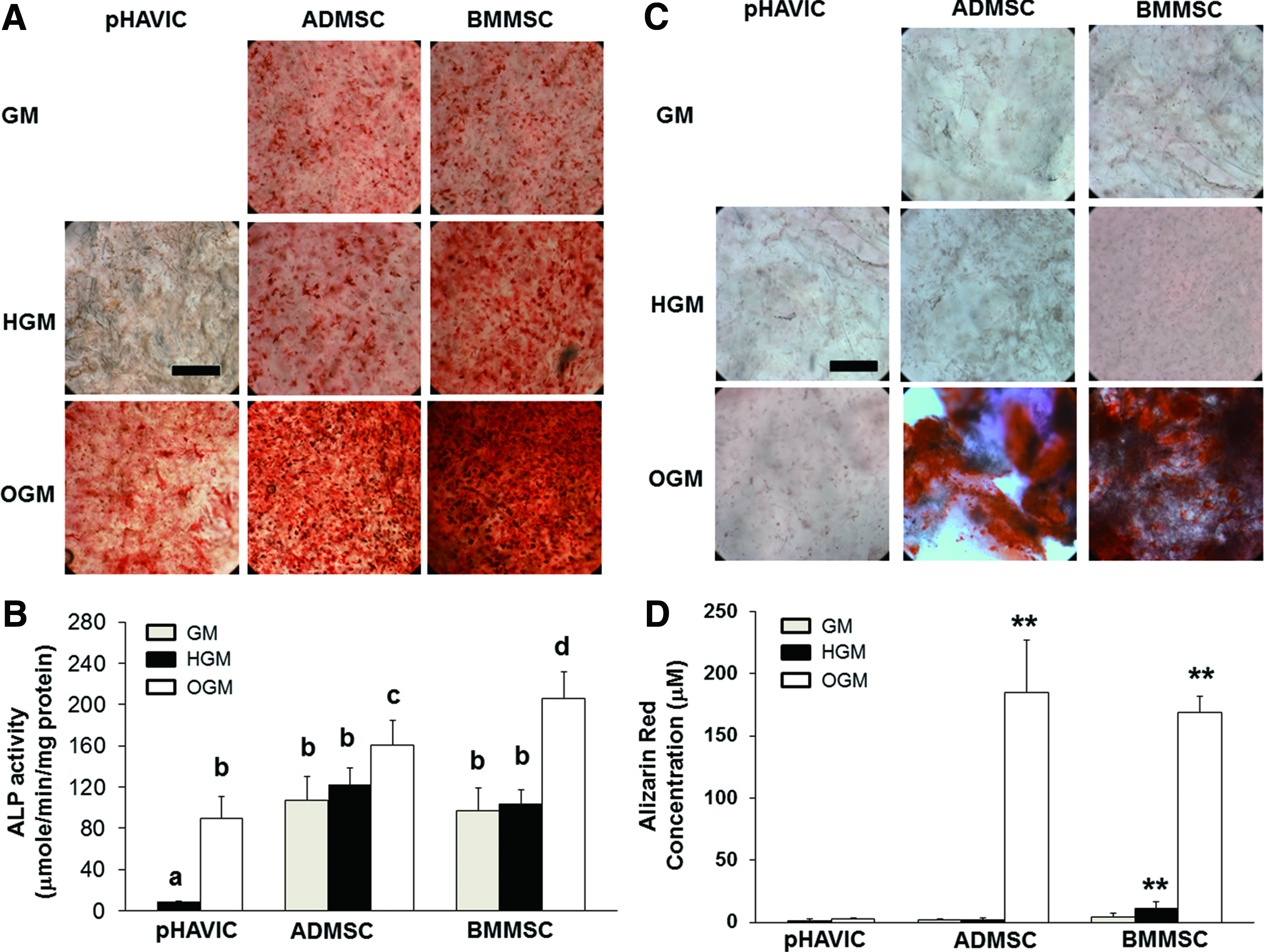
Osteogenic differentiation of pHAVIC, ADMSC, and BMMSC conditioned in osteogenic medium (OGM), GM, or HGM for 28 days.
pHAVIC and ADMSC were negative to Alizarin red S staining in GM and HGM (Fig. 2C, D). Although BMMSC showed much higher absorbance of Alizarin red S in HGM than GM, no obvious calcium deposition was observed for BMMSC in HGM. In OGM, both ADMSC and BMMSC had extensive calcium deposition, whereas pHAVIC did not show calcium deposition.
All three cell types were negative to Oil red O staining regardless of base culture medium applied, indicating no spontaneous adipogenic differentiation tendency (Fig. 3A). In AGM, pHAVIC, ADMSC, and BMMSC showed significantly more lipid accumulation (Fig. 3A, B). ADMSC also showed significantly higher lipid accumulation compared with pHAVIC.
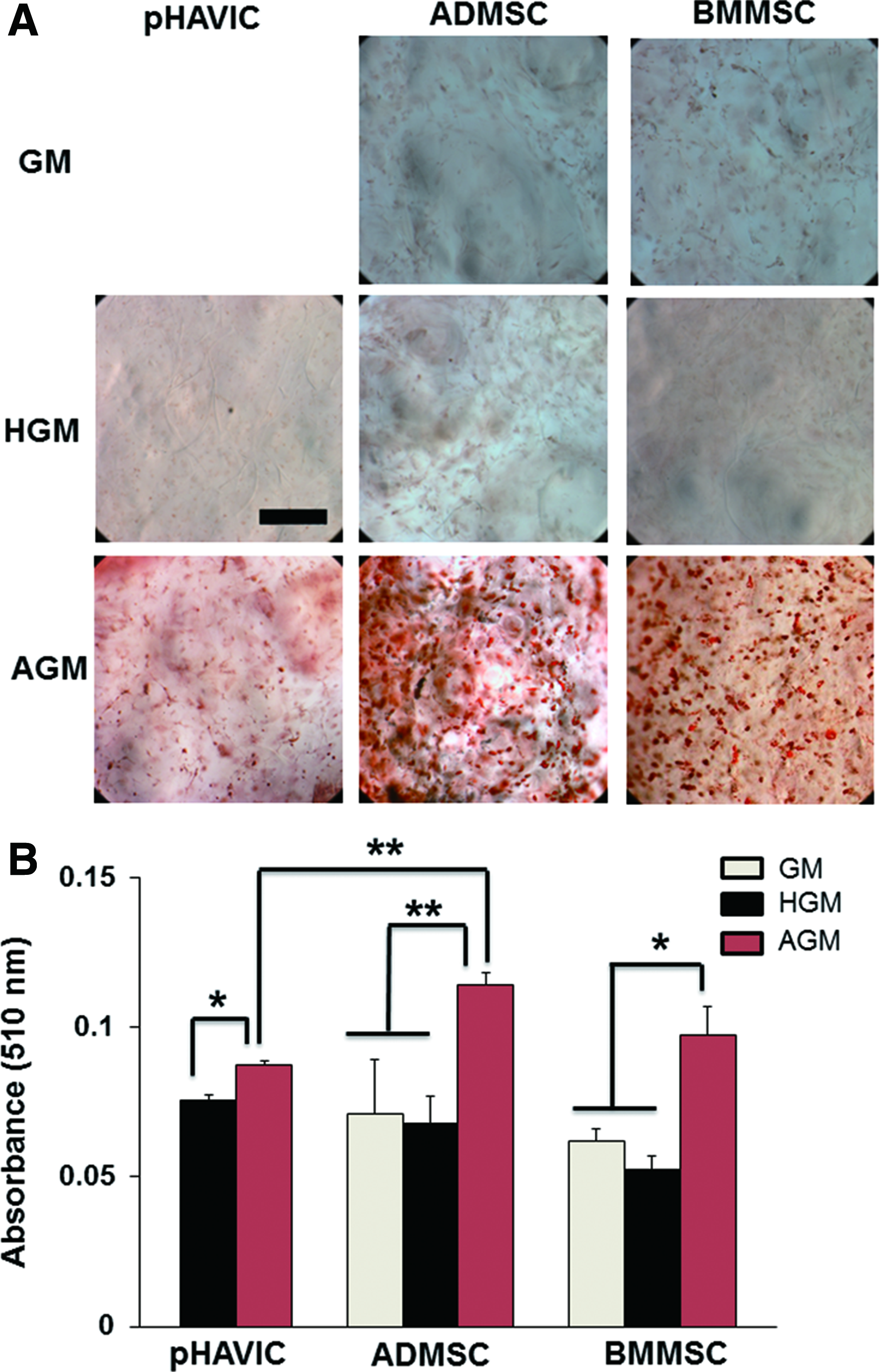
Adipogenic differentiation of pHAVIC, ADMSC, and BMMSC conditioned in adipogenic medium (AGM), GM, or HGM for 28 days.
We quantitatively analyzed Sox9 and aggrecan expression to assess chondrogenic differentiation tendency. Sox9 is a transcription factor, is an important early marker for cartilage development, and is also necessary for valve cell proliferation and differentiation. Sox9 was expressed by all three cell types (Fig. 4A). Sox9 was transcriptionally inactive (non-nuclear expressed) in GM culture, but became extensively nuclear expressed in CGM. As expected, Sox9 expression by pHAVIC was comparable to ADMSC cultured in CGM and was greater than BMMSC in CGM (Fig. 4B). The HGM condition upregulated Sox9 expression for both types of MSC compared with GM condition. The expression of both Sox9 and aggrecan, a later stage ECM component of chondrogenesis, was upregulated for ADMSC and BMMSC in CGM compared with GM. However, pHAVIC expressed very limited aggrecan regardless of culture media (Fig. 4B).
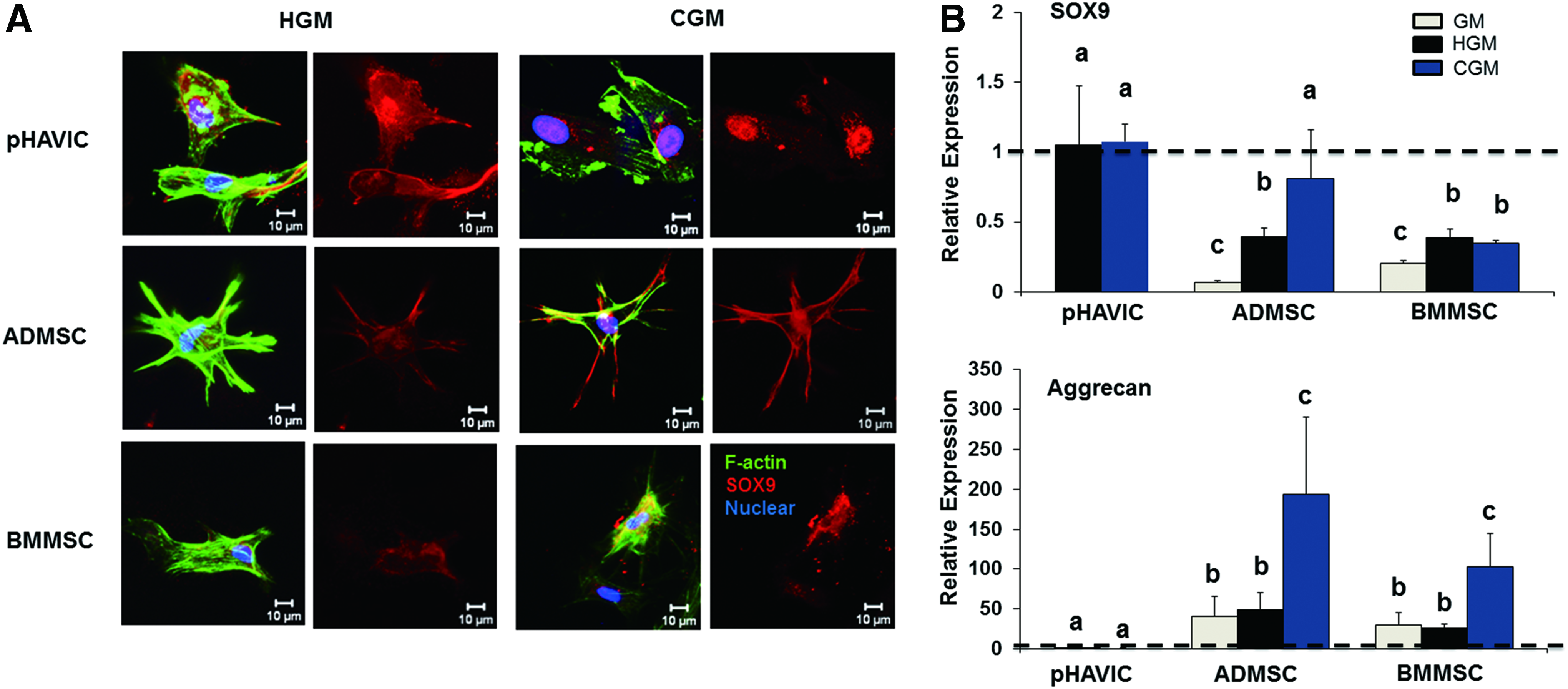
Chondrogenic differentiation of pHAVIC, ADMSC, and BMMSC conditioned in chondrogenic medium (CGM), GM, or HGM for 28 days.
Expression of VIC markers in ADMMSC, BMMSC, and HAVIC
Then, we quantitatively compared gene expression of vimentin, αSMA, periostin, and fibroblast surface protein (FSP)1 between pHAVIC and MSC in different culture media. In injured or diseased valves, VIC transition from quiescent fibroblasts in normal valves into pathogenic myofibroblasts was accompanied with a significant decrease in expression of vimentin and an increase in αSMA.40,41 Periostin is required for normal collagen architecture during heart valve development and lack of periostin resulted in valve leaflet thickening and matrix dysregulation in adult mice.42–44 FSP1 is a very selective and effective biomarker for cardiac fibroblasts, 45 and most HAVIC express FSP1. 46
ADMSC in GM had significantly lower expression of vimentin and αSMA compared with pHAVIC (Fig. 5A, B). HGM culture, however, increased expression of vimentin, αSMA, and periostin in ADMSC to the same level of HAVIC (Fig. 5A–C). BMMSC expressed a significantly lower level of vimentin in all the conditions and a significantly lower level of αSMA in GM and HGM (Fig. 5A, B). ADMSC expressed FSP1 to a greater degree than pHAVIC and BMMSC (Fig. 5D), indicating they are inherently closer to fibroblasts. Treatment of MSC with CGM caused a significant upregulation of vimentin, αSMA, and periostin expression for ADMSC and HAVIC, and αSMA and periostin expression for BMMSC.
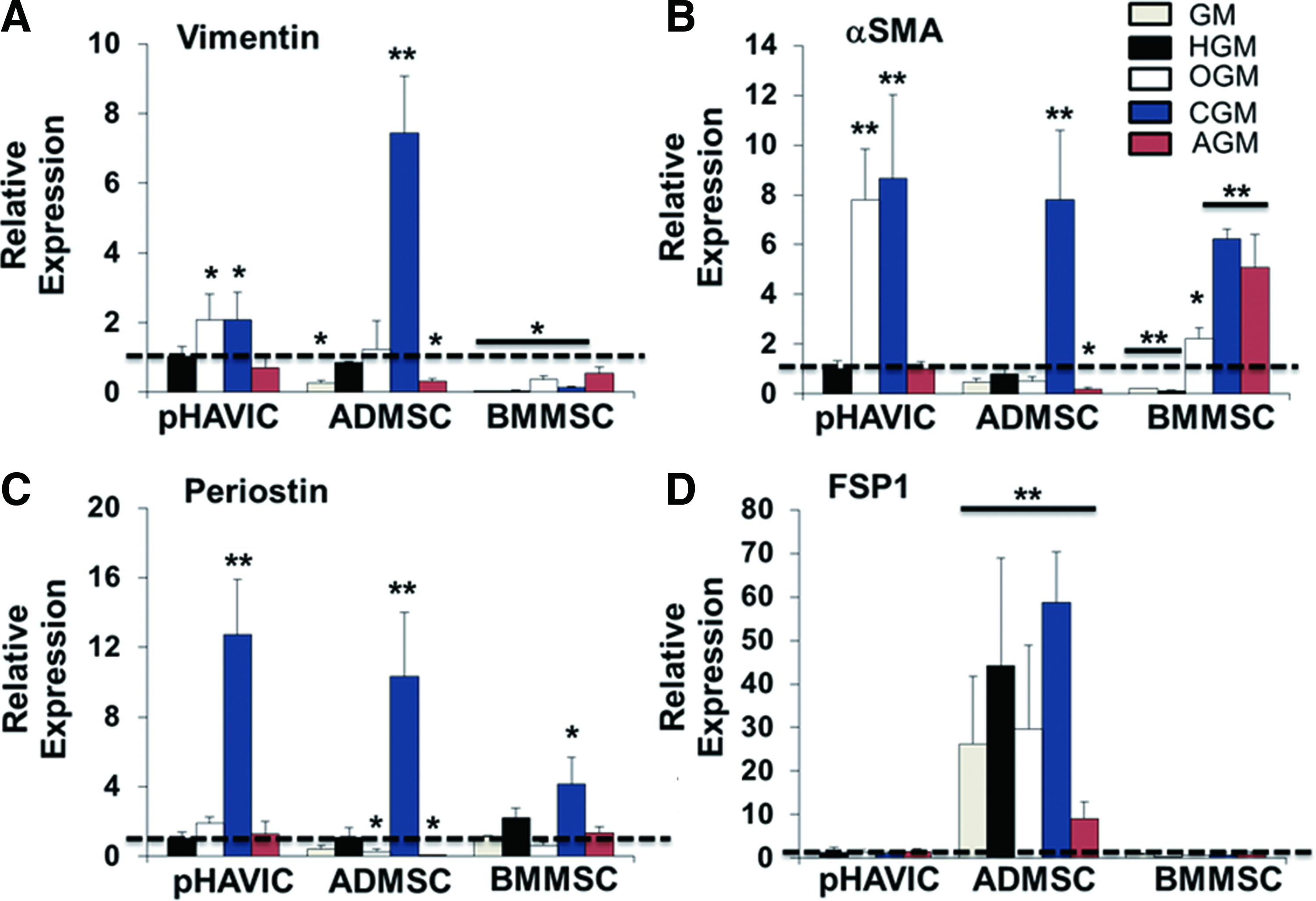
Real-time PCR analysis of
Immunohistochemical staining showed that HAVIC, ADMSC, and BMMSC expressed both vimentin and αSMA (Fig. 6). BMMSC expressed less vimentin and relatively more αSMA in both GM and HGM, showing yellow color in the merged images. This demonstrated that BMMSC had a more myofibroblastic phenotype.
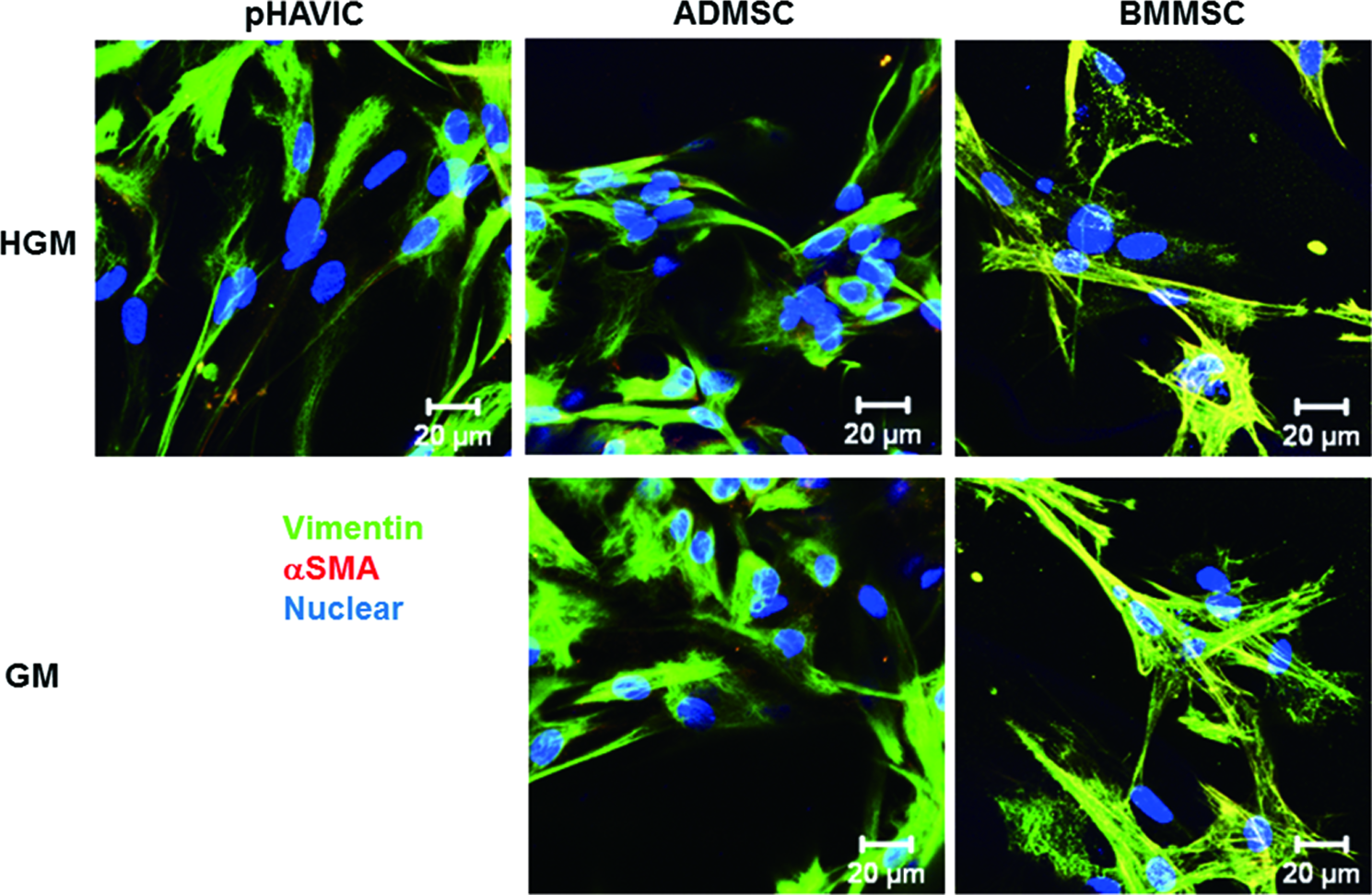
Immunohistochemical staining for αSMA (red); vimentin (green); and nuclei (blue) within encapsulated pHAVIC, ADMSC, and BMMSC in GM and HGM after 28-day culture. Color images available online at
Expression of biomarkers that distinguish BMMSC from fibroblasts
CD106 (vascular cell adhesion molecule 1), integrin alpha 11 (ITGA11), and matrix metalloproteinase (MMP) 1 expression were previously shown to differentiate between BMMSC and fibroblasts. 47 CD106 and ITGA11 were highly expressed in BMMSC in both GM and HGM than pHAVIC, whereas MMP1, the myofibroblast marker, was highly expressed in pHAVIC and much less in BMMSC (Fig. 7A, B). These results were consistent with previous reports. 47 However, ADMSC expressed the same level of CD106 compared with pHAVIC (Fig. 7A). This indicates that CD106 can be used as a biomarker to distinguish BMMSC from fibroblasts but cannot discriminate between ADMSC and pHAVIC. ADMSC in GM had significantly greater expression of ITGA11, but lesser MMP1 compared with pHAVIC. Conditioning in HGM, however, downregulated ITGA11 and upregulated the expression of MMP1 to similar levels of pHAVIC (Fig. 7B, C). Together, these results suggest that HGM promoted ADMSC differentiation toward pHAVIC.
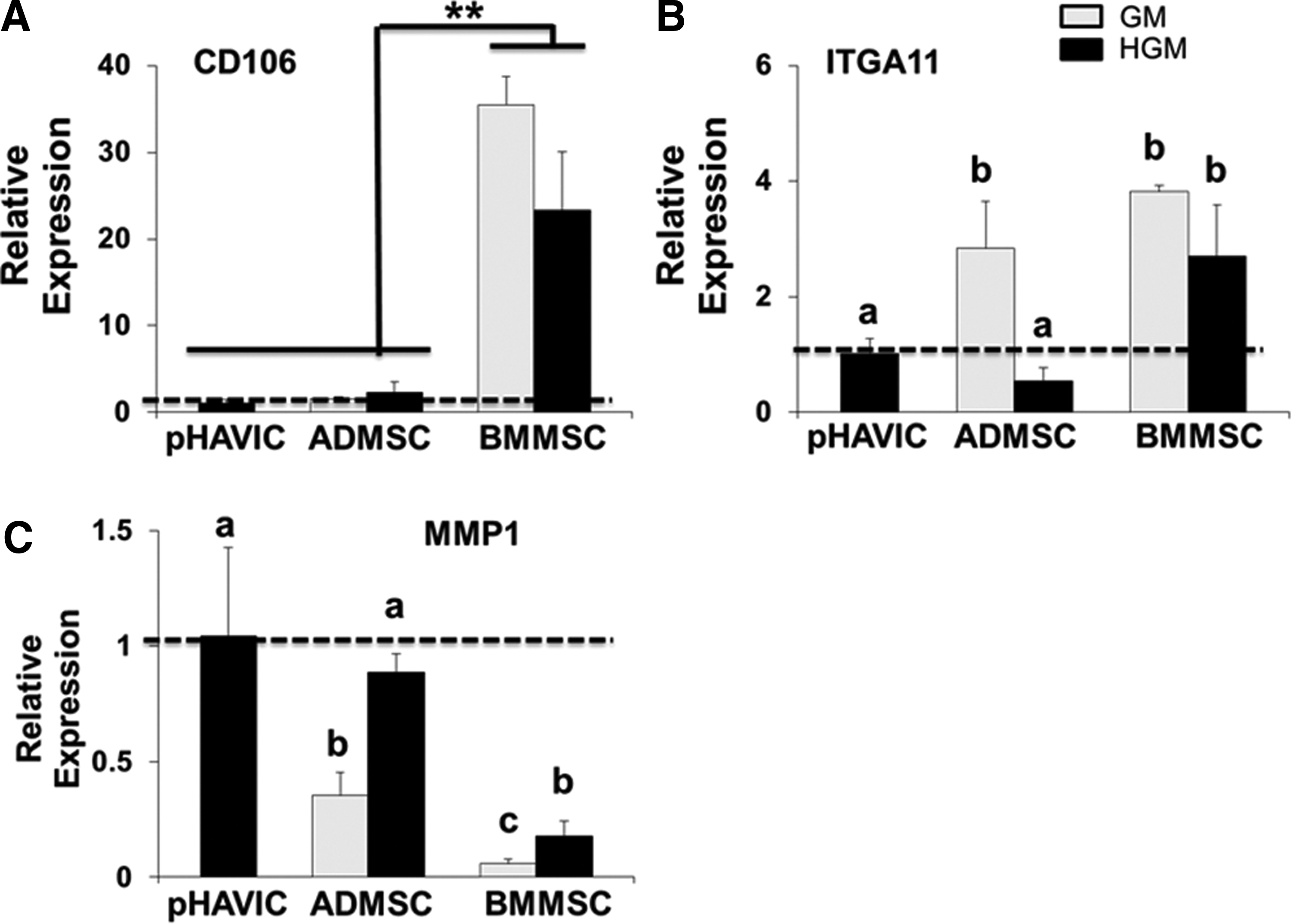
Comparative expression of biomarkers that distinguish MSC from fibroblasts. Real-time PCR analysis of
Dynamic remodeling of ECM by pHAVIC, ADMSC, and BMMSC
Generally, total GAG, collagen, and elastin content varied with cell type and culture time, indicating that ECM is dynamically remodeled, and different ECM components have different deposition rates in each cell type. GAG content increased significantly during middle time periods (day 7 to 14, Fig. 8A), but leveled off at later time points (day 14 to 28). ADMSC had the lowest GAG content over time. Collagen content did not change significantly over time for pHAVIC and ADMSC (Fig. 8B). Collagen content in BMMSC populated gels increased significantly between day 7 and 14 before returning to day 7 levels by day 28. ADMSC had the highest collagen production among the three cell types. The elastin content for ADMSC group greatly increased from day 7 to 14 and was then maintained on day 28. pHAVIC and BMMSC, on the other hand, maintained elastin content during the initial time intervals but markedly increased elastin content by day 28 (Fig. 8C). Of note, ADMSC populated hydrogels had the highest elastin content after the 28 day culture period. Histologically, the pHAVIC-laden hydrogels exhibited more intense positive Safranin O within the lacunae, confirming that more GAGs were expressed and accumulated by pHAVIC in comparison to ADMSC. In contrast, hydrogels with ADMSC showed more intense collagen staining (blue, Masson's trichrome) staining and more elastin fibers (Verhoeff's stain) compared with those with HAVIC and BMMSC (Supplementary Fig. S2). Together, these results support the fact that BMMSC-populated hydrogels have a similar level of GAG, collagen, and elastin contents and ECM remodeling pattern as pHAVIC.
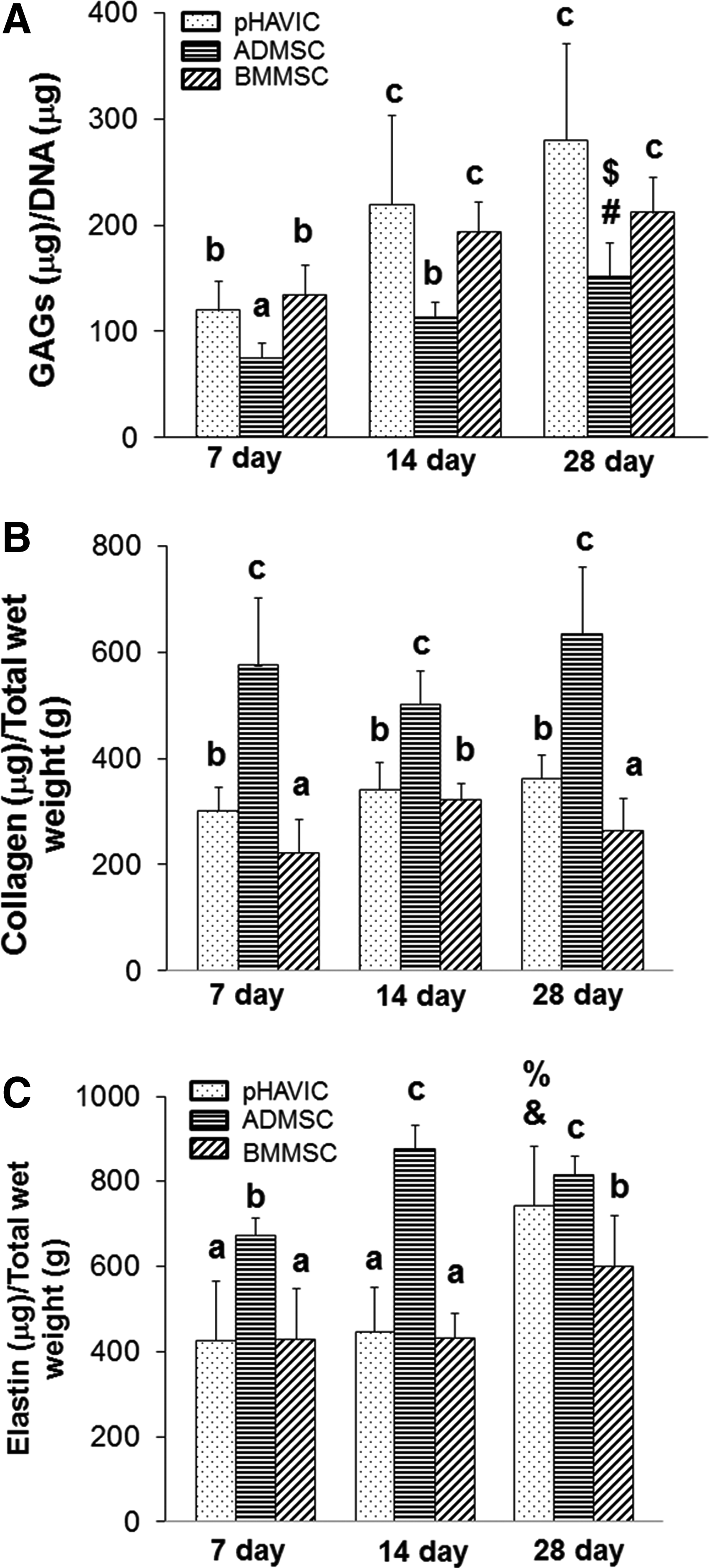
Quantitative analysis of GAGs
Effects of bFGF conjugation in hydrogel matrices on MSC differentiation
As previously demonstrated, soluble bFGF containing HGM promoted differentiation of ADMSC and HAVIC toward fibroblastic phenotype. We thus conjugated bFGF to PEG and covalently immobilized within our hybrid hydrogels network. Covalent bonding of growth factors to hydrogel matrix provides a more prolonged release profile, and the conjugated growth factors have more slowly degraded and internalized features.48,49 Conjugation of bFGF to ACRYL-PEG-SCM was confirmed via Western blot (Fig. 9A). Native bFGF appears in two bands, at around 34 and 17 kDa, indicating both dimer and monomer species, respectively. The band for PEG-conjugated bFGF samples was present at higher molecular weights, indicating successful PEGylation of the growth factor.50,51 After 28 days in culture, immobilized bFGF significantly upregulated vimentin expression in HAVIC, ADMSC, and BMMSC for around three-, two-, and eight-fold compared with the hydrogels without the growth factor (Fig. 9B). Hydrogels modified with PEG-FGF also greatly promoted αSMA expression in HAVIC, but not in ADMSC and BMMSC. Similarly, HAVIC and BMMSC encapsulated within hydrogels with PEG-FGF showed more periostin expression, and conjugated PEG-FGF also upregulated MMP1 expression in BMMC. Western blot results confirmed the increased expression of vimentin in hydrogels with FGF-conjugated samples (Fig. 9C), indicating that tethered bFGF promoted fibroblast phenotype of these three cell types.
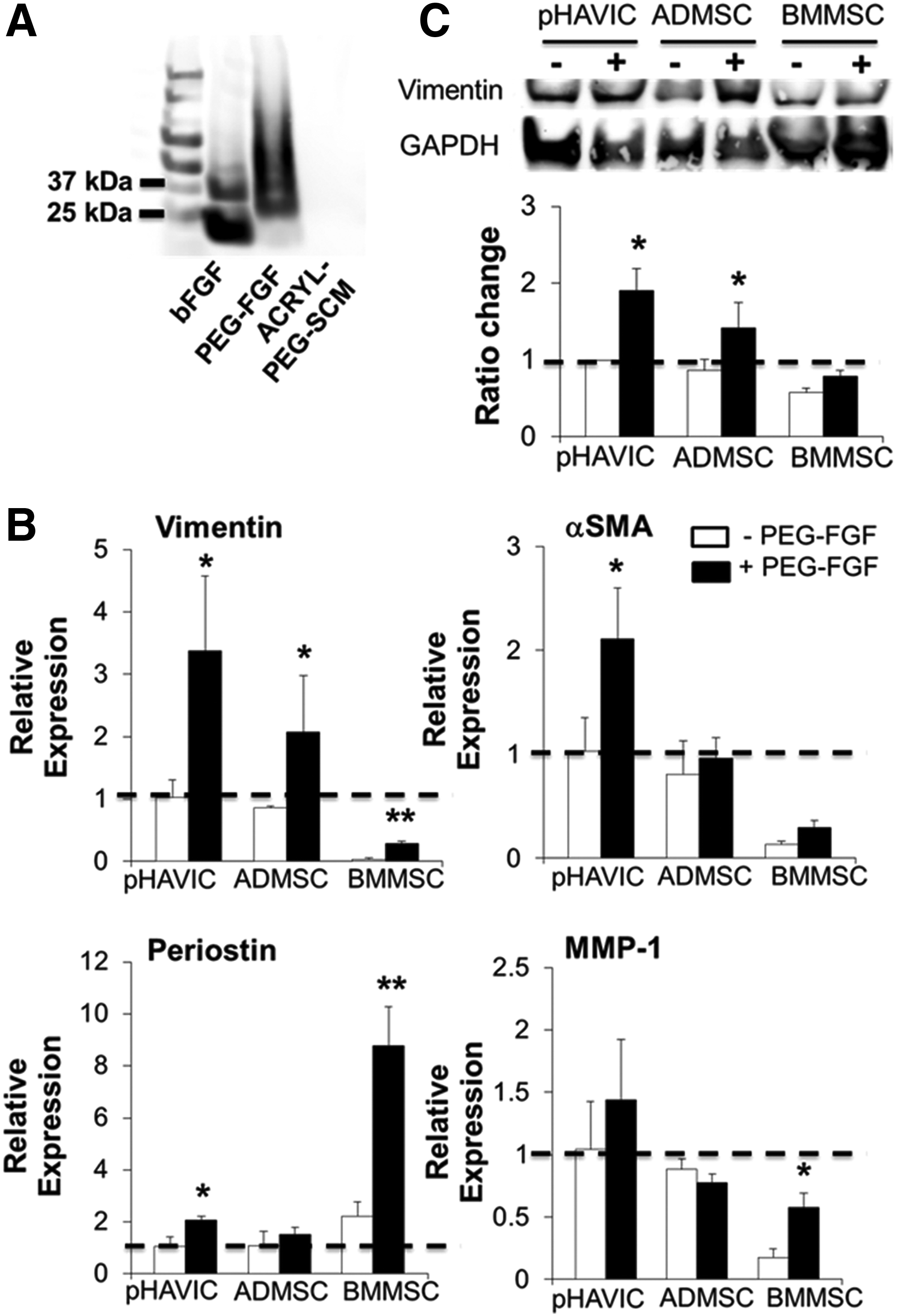
Conjugation of bFGF within engineered hydrogels facilitates the differentiation of MSC toward fibroblastic phenotype.
Discussion
Identification of a proper cell type with phenotype and function similar to that of native VIC is essential for successful development of TEHV with potential for long-term function, similar structure, and phenotypic characteristics to those of the native tissue. This study compares the multi-lineage differentiation capacity, VIC phenotype, and ECM deposition characteristics of pHAVIC, ADMSC, and BMMSC encapsulated within 3D hybrid hydrogels consisting of Me-HA and Me-Gel. Hydrogel stiffness (4.27±0.18 kPa) was tuned to match that of the ventricularis and fibrosa layers of aortic heart valve leaflet (0.25 to 5 kPa). 52
Primary VIC exhibit an elongated myofibroblast-like morphology in confluent 2D culture and rarely formed calcified nodules spontaneously in normal growth media.53,54 The 3D hybrid hydrogels in this study maintained fibroblastic-like phenotypes of pHAVIC without spontaneous osteogenic and adipogenic differentiation and with limited activation in HGM. However, both human ADMSC and BMMSC spontaneously expressed early marker (ALP) of osteogenic differentiation within the 3D hybrid hydrogels. Spontaneous osteogenic differentiation of MSC was reported in both 2D and 3D culture due to aging, preculture on stiff substrates, and increased cytoskeletal tension and/or actinomyosin contraction in some 3D environments.55–58 This indicated that there is potential risk for using ADMSC and BMMSC as cell sources for heart valve tissue engineering. Therefore, any 3D microenvironment for heart valves should be well designed and controlled to limit spontaneous osteogenic differentiation tendency of resident cells.
This study confirms that pHAVIC have multi-lineage differentiation capacity. However, the ALP activity and lipid accumulation in HAVIC were lower than those for ADMSC and BMMSC, indicating that pHAVIC had lower osteogenic and adipogenic potential. In contrast from some studies that demonstrated calcification of adult VIC cultured in 2D environment,25,59 within collagen hydrogels 31 or under cyclic strain,60,61 our results showed that pHAVIC cultured within 3D hybrid hydrogels lacked calcium deposition, suggesting limited osteogenic potential. It may be that the HA component in the hybrid hydrogel suppresses nodule formation, 62 but such suppression was insufficient for the stem cell sources. pHAVIC highly expressed Sox9, an early marker of chondrogenic differentiation, but lacked expression of aggrecan, the late marker of chondrogenic differentiation. This again indicates that the chondrogenic differentiation potential of pHAVIC is limited.
We show that the encapsulated ADMSC express comparable levels of vimentin, αSMA, and periostin as compared with pHAVIC in the HGM, which contained bFGF and EGF. bFGF is reported to promote VIC repair through the activation of Akt1 signaling pathway and transforming growth factor-β/Smad signaling pathway.63–65 bFGF plus EGF can trigger Wnt signaling pathway to induce epithelial-mesenchymal transition, which is an important process in the embryonic development of the heart valve.66,67 The combination of bFGF and EGF can also maintain fibroblastic phenotype by inhibiting TGF-β-induced activation 68 and preserving multipotential properties of ovine BMMSC. 69 In addition, Song et al. and Biver et al. found that FGF2 could inhibit bone morphogenic protein-induced osteogenic differentiation of MSC.70,71 The HGM was effective to induce ADMSC toward pHAVIC phenotype. However, the expression of vimentin and αSMA in BMMSC was significantly lower than pHAVIC and ADMSC even in the HGM, indicating that the induced ADMSC have a phenotype more typical of pHAVIC. Sohier et al. showed that HAVIC consistently expressed CD44 and FSA over culture, but calponin and myosin were not detected.23,46,72 This group also demonstrated that ADMSC expressed a similar level of CD44 and FSA to HAVIC on nanofibrillar scaffolds. 72 Tedder and his coworker dynamically conditioned decellularized porcine pericardium and spongiosa scaffolds seeded with BMMSC in a bioreactor and found that BMMSC were positive for vimentin, which was similar to HAVIC.73,74 This indicates that biomechanical stimuli are likely also beneficial to promote MSC differentiation toward valve phenotypes.
Since external addition of soluble bFGF facilitated differentiation of ADMSC and HAVIC toward fibroblastic phenotype, we further incorporated bFGF within our hybrid hydrogels by conjugating bFGF to PEG and covalently immobilizing in the hydrogel network. Soluble growth factors will diffuse and degrade very rapidly in vivo, compromising their ability for specific and local induction. Immobilization of growth factors would enable prolonged functional signaling and controlled release profile. 48 PEGylated growth factors were proved to maintain their biological activity after chemical coupling and to promote cell–cell contact formation.49,51,75 The conjugated bFGF not only further promoted the fibroblastic differentiation of HAVIC and ADMSC but also significantly upregulated vimentin expression in BMMSC, which was not achieved by soluble bFGF. This indicated the importance to regulate both internal microenvironment and external environment to drive MSC differentiation toward the target fibroblastic phenotype.
We also comparatively analyzed several gene expression profiles in pHAVIC and MSC, since HAVIC in normal adult and pediatric valves show predominantly quiescent fibroblast phenotype. Although both MSC and fibroblasts have been well studied, differences between these two cell types are not fully understood. Fibroblasts were reported to share mesenchymal phenotypes and multi-lineage differentiation potential with ADMSC. 76 In addition, Halfon et al. demonstrated that CD106, ITGA11, and MMP1 had robust differences of expression in BMMSC and fibroblasts and could be used for identifying BMMSC. 47 We confirm that these markers can discriminate between BMMSC and normal HAVIC. However, ADMSC expressed a significantly lower level of CD106 when compared with BMMSC, but similar to pHAVIC. Kern et al. also compared BMMSC and ADMSC and demonstrated that the expression of all tested markers was similar in both cell types except CD106. 33 This indicates that CD106 is probably not a reliable marker to distinguish ADMSC from pHAVIC. After conditioning in HGM, both BMMSC and ADMSC had downregulation of ITGA11 and upregulation of MMP1, indicating the differentiation toward fibroblast phenotypes.
Another important function of VIC is to synthesize and remodel the ECM to maintain sufficient strength and durability for withstanding substantial mechanical stress and strain.4,77 The aortic valve leaflet is known to exhibit a complex tri-layer striation with ECM components, mainly GAGs, collagen, and elastin. 77 We thus quantitatively evaluated and compared the secretion and remodeling of these ECM components. We observed that all these three cell types were able to dynamically produce and remodel their own matrix within the hydrogels. Histological staining and biochemical assay results showed that ADMSC-laden hybrid hydrogels had lower GAGs and higher collagen and elastin contents compared with those with BMMSC and pHAVIC. Compared with ADMSC, BMMSC generally showed more similarity to GAG, collagen, and elastin contents and ECM remodeling pattern to pHAVIC within 3D hybrid hydrogels. Although valvular ECM have the stress-bearing capacity and can transfer the load and absorb shocks during the valve cycle, 78 they also influence VIC responses and contribute to calcific valve disease development. 79 In diseased valves, dense fibrosis is usually first observed, with significant collagen and GAGs accumulation and thickening of the valve leaflets.80,81 The decrease of elastin fiber content and increase of disorganized and fragmented elastin have been shown to induce ALP activity and further calcification. 82
Conclusions
This study compares the differentiation of ADMSC and BMMSC toward pHAVIC within 3D bioactive hybrid hydrogels. We have confirmed that pHAVIC have multi-lineage differentiation capacity, but that capacity is limited to the early stages of differentiation and is much less than that of ADMSC and BMMSC. When conditioning in HGM, ADMSC exhibits similar biomarkers expression to pHAVIC within 3D microenvironment, while BMMSC generally expressed similar GAG, collagen, and elastin content and ECM remodeling patterns in comparison to pHAVIC. Conjugated bFGF within the hydrogels upregulated vimentin expression in BMMSC and promoted the fibroblastic differentiation of pHAVIC and ADMSC. These results clarify design considerations for using ADMSC or BMMSC as potential cell sources for the development, maturation, and long-term fidelity of engineered heart valve tissues.
Footnotes
Acknowledgments
This work was funded by the American Heart Association Postdoctoral Fellowship (13POST17220071), Morgan Family Foundation, The Hartwell Foundation, the National Science Foundation (CBET-0955172), NSF Graduate Research Fellowship, Foundation LeDucq MITRAL Transnational Network, and the National Institutes of Health (HL110328, HL118672). The authors thank Dr. Jonathan Chen in the Department of Cardiothoracic Surgery at Weill Cornell Medical College [now at Seattle Children's Hospital] for providing human aortic valves. They would also like to thank Cornell University Biotechnology Resource Center (BRC), which is supported by National Institutes of Health (NIH 1S10RR025502-01) for the assistance with CLSM imaging.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.