
Abstract
Proliferation, integration, and neurite extension of PC12 cells, a widely used culture model for cholinergic neurons, were studied in nanocellulose scaffolds biosynthesized by Gluconacetobacter xylinus to allow a three-dimensional (3D) extension of neurites better mimicking neuronal networks in tissue. The interaction with control scaffolds was compared with cationized nanocellulose (trimethyl ammonium betahydroxy propyl [TMAHP] cellulose) to investigate the impact of surface charges on the cell interaction mechanisms. Furthermore, coatings with extracellular matrix proteins (collagen, fibronectin, and laminin) were investigated to determine the importance of integrin-mediated cell attachment. Cell proliferation was evaluated by a cellular proliferation assay, while cell integration and neurite propagation were studied by simultaneous label-free Coherent anti-Stokes Raman Scattering and second harmonic generation microscopy, providing 3D images of PC12 cells and arrangement of nanocellulose fibrils, respectively. Cell attachment and proliferation were enhanced by TMAHP modification, but not by protein coating. Protein coating instead promoted active interaction between the cells and the scaffold, hence lateral cell migration and integration. Irrespective of surface modification, deepest cell integration measured was one to two cell layers, whereas neurites have a capacity to integrate deeper than the cell bodies in the scaffold due to their fine dimensions and amoeba-like migration pattern. Neurites with lengths of >50 μm were observed, successfully connecting individual cells and cell clusters. In conclusion, TMAHP-modified nanocellulose scaffolds promote initial cellular scaffold adhesion, which combined with additional cell–scaffold treatments enables further formation of 3D neuronal networks.
Introduction
T
Hence, there is a strong interest in both academia and industry to identify assays for neuronal cell growth in 3D tissue-mimicking environments, including promising examples of laminin-coated agarose gels, 8 self-assembling beta-sheet oligopeptide scaffolds, 9 and collagen, laminin, and fibronectin gels.10,11 The general consensus of 3D in vitro cultivation studies, comparing different materials, is that neurons grown in collagen hydrogels show longer periods of survival and better neurite extension than other hydrogels.12–14 However, while collagen hydrogels represent a correct chemical microenvironment and architecture at all length scales, the raw material is extracted from animals, posing problems with batch-to-batch variations.
We have investigated a novel nanocellulose/ECM protein platform, which offers a competitive alternative to other biopolymer scaffolds as the nanocellulose scaffold material (i) can be produced on an industry scale, (ii) forms reproducible stable environments; cellulose nanofibrils are not degraded with time, and (iii) is modular at macro to molecular levels by electrospinning and bioprinting. It further allows the introduction of surface charges and coating with ECM proteins displaying specific bioactive motifs that regulate cell interaction.
We propose nanocellulose synthesized by Gluconacetobacter xylinus 15 as an alternative host material for 3D neuronal growth. It has shown promising results for several tissue-mimicking designs, 16 for example, as a scaffold for endothelial cells, 17 smooth muscle cells, 18 chondrocytes, 19 and osteoprogenitor cells. 20 The advantages of these nanocellulose scaffolds include the following: (i) a fibril structure similar to that of collagen scaffolds, (ii) a tissue-like microenvironment characterized by an exceptionally high water content without compromised mechanical strength,15,16,21 (iii) good biocompatibility, 22 (iv) a nanofibril network with a morphology that can be tailored by varying the microbial cultivation conditions, 21 (v) surface chemistry that can be readily optimized for cell adhesion by different surface modification protocols,17,23 and (vi) a robustness toward chemical and thermal treatment, such as sterilization processes.
In this work, we investigate the potential of nanocellulose as a scaffold material for neuronal cells using the PC12 cell line, derived from a rat adrenal gland pheochromocytoma. 24 The capability of PC12 cells to grow in 3D scaffolds of agarose, collagen, fibrin, fibronectin, and laminin has been evaluated previously,11,12 making it a good model system for further comparison. While PC12 cells are different from primary neurons in many aspects, they are nevertheless a well-established model widely used to study several mechanisms present in neurons such as neurosecretion (dopamine), exocytosis mechanisms, cell differentiation, and neurite outgrowth after neuronal growth factor (NGF) stimulation. 25 Accordingly, PC12 cells have been used to decipher transcription mechanisms involved in differentiation, also present in neurons. 26
While no surface modifications have been needed for the attachment and growth of smooth muscle cells, chondrocytes, and osteoprogenitor cells on cellulose scaffolds,18–20 studies comparing the growth of neurons on different hydrogels indicate that these cells are more sensitive to the molecular composition and arrangement of ligands.12–14 Thus, in addition to untreated nanocellulose for PC12 cell growth, we have investigated substrates with different chemical surface modifications.
The 3D cell and neurite arrangement in the different scaffolds, the character of the cell adhesion, and the local impact of the cells on the scaffold structure have been assessed by simultaneous Coherent anti-Stokes Raman Scattering (CARS) and second harmonic generation (SHG) microscopy. CARS microscopy has been used to visualize the cell body and neurites of the PC12 cells and has also given good specific contrast for other cell types in nanocellulose scaffolds.20,27,28 The nanocellulose fibrils in the support network are simultaneously visualized by filtering out the scattered second harmonic (SH) of the probe light.27,29 Together with conventional assay measurements of the overall cell proliferation, this provides detailed insights in neuronal cell growth in a 3D tissue matrix.
Materials and Methods
Synthesis of bacterial nanocellulose scaffolds
Cultures of the strain G. xylinus subsp. sucrofermentans BPR2001, trade number: 700178, purchased from the American Type Culture Collection were maintained on complex media as described by Matsuoka et al. 30 Nanocellulose scaffolds were grown with 0.2 mL of bacterial preculture (containing ∼2 million bacteria as determined by OD600nm) added to the 2 mL medium in the wells of a cell cultivation plate that was incubated in a bacteriological cabinet at 30°C for 3 days. The cellulose pellicle scaffolds formed were then washed in two successive 0.1 M NaOH baths at 60°C for 1 day each, followed by two washes in distilled water at 60°C for 1 day each. Finally, all cellulose scaffolds were autoclaved in water before surface modification.
Nanocellulose scaffold surface modification
In addition to unmodified nanocellulose scaffolds, different surface modification protocols were applied to investigate to what extent the physicochemical and molecular properties of the scaffold influence cell adhesion. The ionic charge of the substrate was modified by chemical treatment, and/or the fibrils were coated with ECM-relevant proteins. For the chemical treatment, we essentially followed a protocol published by Watanabe et al. 23 where electrical charges were introduced to the surface of the nanocellulose fibrils forming trimethyl ammonium betahydroxy propyl (TMAHP) cellulose. Briefly, wet cellulose scaffolds (∼5 g) were treated in 0.5 M NaOH at 70°C for 1 h, then 0.3 g of glycidyl trimethyl ammonium chloride (39.6 mM) (Sigma-Aldrich) was added and the sample was incubated at 70°C for 5 h. Finally, the scaffolds were neutralized with a 6 M HCl solution followed by an extra washing step with a 0.1 M NaCl 0.1 M Na (acetate-acetic acid) pH 7.4 buffer solution to stabilize the pH of the modified scaffold.
The TMAHP-cellulose scaffolds as well as unmodified scaffolds were washed twice with 1 mL of cell culture medium and incubated in the cell culture medium overnight at 37°C. A set of scaffolds was further coated with proteins, bovine collagen I, natural mouse laminin, and bovine plasma fibronectin (Invitrogen). A protein concentration of 50 μg/mL was achieved by dilution with 0.02 M acetic acid for collagen or phosphate-buffered saline solution for laminin and fibronectin. The samples were incubated in protein solution for 24 h at 37°C, after which the solution was removed. The scaffolds were then rinsed three times with the warm cell culture medium.
PC12 cell seeding
Rat pheochromocytoma PC12 cells were purchased from the American Type Culture Collection and maintained as described previously. 31 In brief, cells were grown in collagen IV-coated culture flasks (BD Biosciences) in a 7% CO2 atmosphere at 37°C and subcultured approximately every 7–9 days or when confluence was obtained. Cells were extracted for scaffold seeding between 2 and 4 days after subculturing. Each scaffold was seeded with 200,000 cells in the medium and incubated at 37°C. Cells were grown on the nanocellulose scaffolds for 7 days, and the medium was changed every second day. The cell growth medium was composed of RPMI-1640, with 10% equine serum (PAA, Fisher Scientific), 5% fetal bovine serum (PAA, Fisher Scientific), and 0.4% penicillin–streptomycin solution (Sigma-Aldrich). For a subset of samples, the medium was during day 1 replaced by the differentiation medium composed of 3% equine serum, 1.5% fetal bovine serum, 0.4% penicillin–streptomycin solution, and mouse 2.5S NGF (Invitrogen) at a concentration of 50 ng/mL. For these samples, PC12 cells differentiated within 2–4 days.
Cellular proliferation assay
A CellTiter 96® AQueous Non-Radioactive Cell Proliferation Assay (Promega) was used to determine the viability and proliferation of the PC12 cells on nanocellulose scaffolds. We followed the protocol supplied by the manufacturer, but increased the reagent incubation time from 4 to 16 h, as the amount of soluble formazan formed was found insufficient. The PC12 cells seem to have a relatively slow metabolism, at least within the first hours of growth, for which the standard curve is determined. Cell viability on the scaffolds after 1 and 7 days of culture was analyzed by statistical methods using the SPSS software. The variance of the data was analyzed by means of an analysis of variance (ANOVA) test, and the significance levels between six groups were evaluated by a post hoc Tukey's test.
CARS and SHG microscopy
The overall experimental procedure and the nonlinear microscope are schematically illustrated in Figure 1. The main components of the microscopy setup are a pulsed Nd:Vanadate laser (HighQ Lasers GmbH), an optical parametric oscillator (Levante Emerald OPO APE GmbH), and an inverted microscope (Eclipse TE-2000-E Nikon). The Nd:Vanadate laser emits pulses with 7 ps duration at a repetition rate of 76 MHz on the fundamental wavelength 1064 nm as well as the SH at 532 nm. The 532 nm beam is used to pump the OPO, which generates a tunable laser beam in the wavelength range 690–990 nm. The OPO beam, tuned to 817 nm, and the 1064 nm Nd:Vanadate beam are spatially overlapped and guided into the microscope. A delay line in the OPO beam path is used to obtain temporal overlap at the sample position between pulses from the two beams. A Nikon Plan Fluor 40× oil immersion objective (N.A. 1.3) focuses the beams, which are scanned over the sample surface using a Nikon Eclipse C1 beam scanner.

Three-dimensional imaging is achieved by scanning the beams over different horizontal planes, accessed by vertical translation of the objective. The typical lateral and axial spatial resolutions are 0.4 and 1 μm, respectively, corresponding to a probe volume of 0.1 μm3. The combined laser beams (817 and 1064 nm) induced a CARS process probing the CH2 vibration at a frequency of 2845 cm−1, resulting in a signal emitted at 663 nm. In addition, the laser beams also induced SHG in the noncentrosymmetric cellulose material, and the SH of the 817 nm beam was detected at 409 nm. Typical excitation powers at the sample were 35 mW per laser beam. The back-scattered CARS and SHG signals were collected by the objective, separated by a dichroic beamsplitter, and focused on single photon counting photomultiplier tubes (PMTs; PMC 100-1 and PMC 100-20 Hamamatsu). Band pass filters were used to suppress background contributions.
For microscopy, nanocellulose scaffolds seeded with cells were mounted between two 0.17-mm-thick #1.5 glass cover slides and the sample was oriented with the cell layer toward the objective, see Figure 1. Stacks of CARS and SHG images were collected simultaneously with an acquisition time of 60 s per plane and field of view ranging from 20 × 20 to 150 × 150 μm2. The ImageJ software was used for data analysis as well as rendering of overlay images and volume views, showing the 3D arrangement of cells and cellulose. Cell integration depths and neurite lengths were evaluated for 68 CARS/SHG stacks and 21 CARS stacks, respectively, by manual inspection in ImageJ.
The benefits of these microscopy techniques for visualization of 3D growth of neuronal networks in thick scaffolds are multifold: (i) the near-infrared probe light provides large optical probe depth; (ii) no artificial fluorophores or labels are needed, which tend to distribute inhomogeneously in thicker samples, thus, not reaching deeper located cells; (iii) true 3D imaging as the signal generation is confined to the high-intensity region of the laser beam focal volume; and (iv) the arrangement and mutual interaction between cells and matrix components can be visualized under native soft wet conditions as no sample preparation or physical sectioning that might modify the 3D morphology of the scaffold and/or the fragile connections formed between the cells and the scaffold components is required.
Examples of a bright-field image and corresponding CARS and SHG images of a pair of PC12 cells growing on a collagen-coated TMAHP-cellulose scaffold with NGF added to the medium are shown in Figure 1b–e. The cell bodies, as seen in both the bright-field (Fig. 1b) and the CARS (Fig. 1c) images, show a classical rounded shape, confirming that PC12 cells conserve their morphology on the cellulose scaffolds. In addition, the outgrowth of a neurite can be distinguished. Thus, despite their fine dimensions and the limited content of lipids, neurites can clearly be visualized by CARS microscopy without any labeling. The lipid content in the cell membranes and the active filaments of the neurites seem to provide sufficient image contrast. This is a simple but important observation as it brings forward CARS microscopy as an attractive instrument within the neurosciences, offering the unique capabilities of label-free 3D imaging of neurite outgrowth. Figure 1d further shows the corresponding SHG image of the distribution of cellulose fibers in the plane interfacing the cells, illustrating the tight interaction between cells and cellulose fibers. Finally, the volume rendering of CARS and SHG overlay images in Figure 1e reveals the integration depth of the cells and the 3D orientation of the neurite.
Results and Discussion
Cellular proliferation assay
The general capacity of the PC12 cells to proliferate on nanocellulose scaffolds, unmodified or treated with different surface modifications, was assessed with a proliferation assay.
The results are presented in Figure 2 as the average number of cells in each group with error bars showing the 95% confidence interval. ANOVA between groups showed none of the averages to be equal, and statistical significances, also indicated in Figure 2, for observed differences between samples were evaluated by the Tukey's post hoc test. After 1 day, data show that PC12 cells were able to better attach on TMAHP-treated scaffolds compared to unmodified bacterial nanocellulose (BNC) scaffolds (p < 0.001). The protein coating had no significant effect at this stage. TMAPH-treated samples showed on an average, a three times higher number of cells compared to nontreated samples 1 day after cell seeding (Fig. 2a). During sample preparations and microscopy measurements, it was observed that cells grown on untreated nanocellulose relatively easily detached from the sample surface, which is in agreement with the low cell adhesion reported for cellulose. 23 In contrast, cells on TMAHP-treated cellulose were not as easily detached, indicating an improved adhesion to the matrix when surface charges are introduced. Rather than direct effects of the ionic charges, a more likely explanation to the enhanced cell adhesion is the unspecific protein binding from the medium, previously reported for TMAHP-treated cellulose membranes. 23 TMAHP-treated samples with protein coatings of collagen, fibronectin, or laminin qualitatively showed similar cell adhesion.

PC12 cell proliferation expressed as number of cells monitored by assay measurements on cellulose scaffolds following 1
Following 7 days of growth, all categories showed a 5- to 10-fold increase in cell proliferation relative to day 1 (Fig. 2). Although the enhancement due to the TMAHP treatment was less apparent at this stage, samples with surface charges showed significantly higher numbers of cells, p < 0.001 for the TMAHP-treated and TMAHP-treated/collagen-coated/NGF scaffolds. The number of cells is typically a factor 1.2 higher for the TMAHP-treated compared with the nontreated samples (Fig. 2b). Differences between BNC alone and BNC with collagen and NGF were significant (p < 0.05), but not between BNC collagen and TMAHP collagen. As expected, the unmodified scaffolds with NGF added showed a somewhat lower number of cells (p < 0.05) compared with the undifferentiated scaffolds (without NGF) where cell proliferation has continued.
From this it can be concluded that PC12 cells are able to grow on unmodified nanocellulose, although the TMAHP modification clearly enhances, in particular, the initial attachment to the scaffold and expedites the process. TMAHP modification of the nanocellulose also gives a more reproducible result as higher variations in cell number were seen in the three BNC scaffold groups. Interestingly, the protein coating did in itself not contribute to any additional improvement of the cell proliferation compared with the uncoated categories. At day 1, protein-coated samples without TMAHP treatment showed cell attachment less than or equal to that of an untreated sample (Fig. 2a). The reason for this could be inadequate protein adsorption due to the inherently low surface charge of unmodified nanocellulose fibrils. In contrast, a slight improvement, however, not statistically significant, is indicated for protein-coated TMAHP-treated cellulose at day 1 (Fig. 2a).
For the protein-coated scaffolds without TMAHP treatment, the addition of NGF to the culturing medium seemed to reduce the proliferation rate (p < 0.05), taken from the data 7 days after cell seeding (Fig. 2b). Accordingly, cell differentiation with neurite outgrowth and reduced proliferation has been reported for PC12 cells following NGF addition. 24 However, also samples with NGF added show an increased number of cells compared with day 1, implying continued proliferation and that not all cells differentiated into a neuron-like phenotype. Indeed, after 7 days of culture, the highest numbers of cells were measured on the TMAHP and TMAHP-NGF scaffolds, both reaching ∼25,000 cells.
CARS and SHG microscopy of cell adhesion and integration
While the proliferation assay gives important information on general trends of the cell establishment on the different scaffold categories, no insights in the character or degree of cell–scaffold interaction, cell migration, or neurite extension can be retrieved. This was instead investigated by simultaneous CARS and SHG microscopy, and data for arrangements of PC12 cells on nanocellulose scaffolds are shown in Figure 3. In general, the cells establish large confluent regions of single-cell layers immersed into the scaffold. In Figure 3a–c, the interaction of PC12 cells with an unmodified nanocellulose scaffold is exemplified. Peripheral cells of the cluster establish adhesion points to nanocellulose fibril bundles, and even actively rearrange them (arrows in Fig. 3a), resulting in a boundary cellulose region surrounding the cells with higher density than the bulk scaffold (see SHG microscopy image in Fig. 3b). The cell layer is superficial and immersed merely ∼5 μm into the scaffold as shown in the 3D CARS/SHG overlay volume view in Figure 3c.

CARS and SHG images of PC12 cells on nanocellulose scaffolds.
Image data for a cluster of cells grown on TMAHP-modified nanocellulose are shown in Figure 3d–f. Nanocellulose fibrils apparently involved in adhesion points appear more distinct (see arrow heads), located also in between cells, and bundle up in larger collections as pointed out by arrows in the 3D CARS/SHG overlay volume view in Figure 3f. Compared with the cells on the unmodified scaffold (Fig. 3a–c), the cells on the TMAHP-modified scaffold show a deeper integration into the scaffold; ∼10 μm (Fig. 3f).
While the protein coating did not show any significant impact on cell proliferation taken from MTS assays, CARS/SHG microscopy images provide additional information. The arrangement of cells on a protein-coated scaffold as depicted in Figure 3g–i (fibronectin) indicates a higher degree of lateral cell mobility, resulting in a larger spread of smaller cell clusters. The cells seem to push their way through the uppermost scaffold layer (∼10 μm), leaving compact regions of nanocellulose along their path, as pointed out by arrows in the SHG volume view in Figure 3h. Their directional migration pattern, traced from the scaffold structure, is clearly illustrated and highlighted by arrows in Figure 3i, representing a cross section along the dashed line in Figure 3g observed from a view point indicated by the star symbol. While cells leading the migration (left arrow in Fig. 3i) show few connections to nanocellulose fibrils, a dense and continuous collection of cellulose nanofibrils connecting to the cells at the tailing edge and below the cell cluster can be observed, as pointed out by the three rightmost arrows in Figure 3i. These images imply that protein coating promotes a more active cell migration into and remodeling of the architecture of nanocellulose scaffolds.
Cell integration into the scaffolds was analyzed from 47 and 21 z-stacks measured on samples with and without TMHAP modification, respectively. For unmodified samples, 14 z-stacks were acquired on native nanocellulose and 7 on protein-coated scaffolds. For the native material, the majority of the z-stacks showed cells with no (5) or low (6) apparent integration into the scaffold, corresponding to depths ≤5 μm as exemplified in Figure 3c. Only four of the unmodified samples showed cell integration ≥10 μm, of which three were protein-coated scaffolds. Out of 10 z-stacks measured on samples with TMAHP treatment only, 2 indicate no cell integration whereas the remaining 8 showed medium integration depths of 4–15 μm. For the 37 z-stacks measured on samples with TMAHP modification and protein coating, 18 showed an integration depth of ≥10 μm; only 4 showed no cell integration and another 4 showed low cell integration.
The larger share of data showing no or low cell integration for unmodified samples is consistent with the observations of low cell adhesion 23 and lower proliferation (cf. Fig. 2) observed for unmodified cellulose. Average integration depths for native and TMAHP-modified nanocellulose are 4 and 6 μm, respectively. While TMAHP treatment clearly improved cell adhesion and proliferation, an effect on cell integration is less evident. A clearer indication was obtained for protein-coated samples with average integration depths of 9 μm irrespective of TMAHP-treatment, suggesting that protein coating promotes cell integration. Even though protein coating indicates some improvement, cell integration in general is rather low for the scaffolds. A possible explanation being that the nanocellulose fibrils cannot be protolytically degraded to allow for further cell integration into the matrix.
CARS and SHG microscopy of cell differentiation
The establishment of a neuronal network requires that cell proliferation as well as cell differentiation, including the development of neurites, is promoted in the scaffold. To evaluate to what extent a nanocellulose scaffold supports natural development of neurites, NGF was added to the culture medium of a subset of samples growing on native as well as TMAHP-modified and collagen-coated scaffolds after 1 day in culture. The response of a small ensemble of cells after 8 days of growth in a TMAHP-modified and collagen-coated scaffold is shown in Figure 4. Neurites extend from all four cells, although at different depths, as can be seen in the series of CARS and SHG microscopy overlay images in Figure 4a, spanning the depths 2.4–7.3 μm. While neurite 1 propagates immediately below the surface of the scaffold, neurite 2 has nestled its way 4–5 μm into the scaffold matrix. Neurites 3–5 submerge even deeper into the scaffold and their outlines are visible down to 7.5 μm. This is clearly illustrated in Figure 4b, where the different planes are color coded from magenta (depth 3 μm) to red (depth 10 μm). The superficial cells and neurite 1 appear here in magenta, neurite 2 in cyan/green, neurites 3 and 4 in yellow, and neurite 5 in yellow/red. A CARS volume view of the cells and the neurites demonstrates this 3D arrangement further in Figure 4c. In this study, it can be noted that all neurites extend below the level of the cell soma, gradually migrating into the scaffold. In the series of CARS/SHG microscopy images in Figure 4a, it is also interesting to note the close interaction between the neurites and the nanocellulose fibrils. This exemplifies that rather than remodeling the scaffold structure, the neurites seem to propagate through the fibrillar scaffold in an amoebic manner, a migration scheme observed in 3D tissue matrixes in contrast to two-dimensional surfaces.
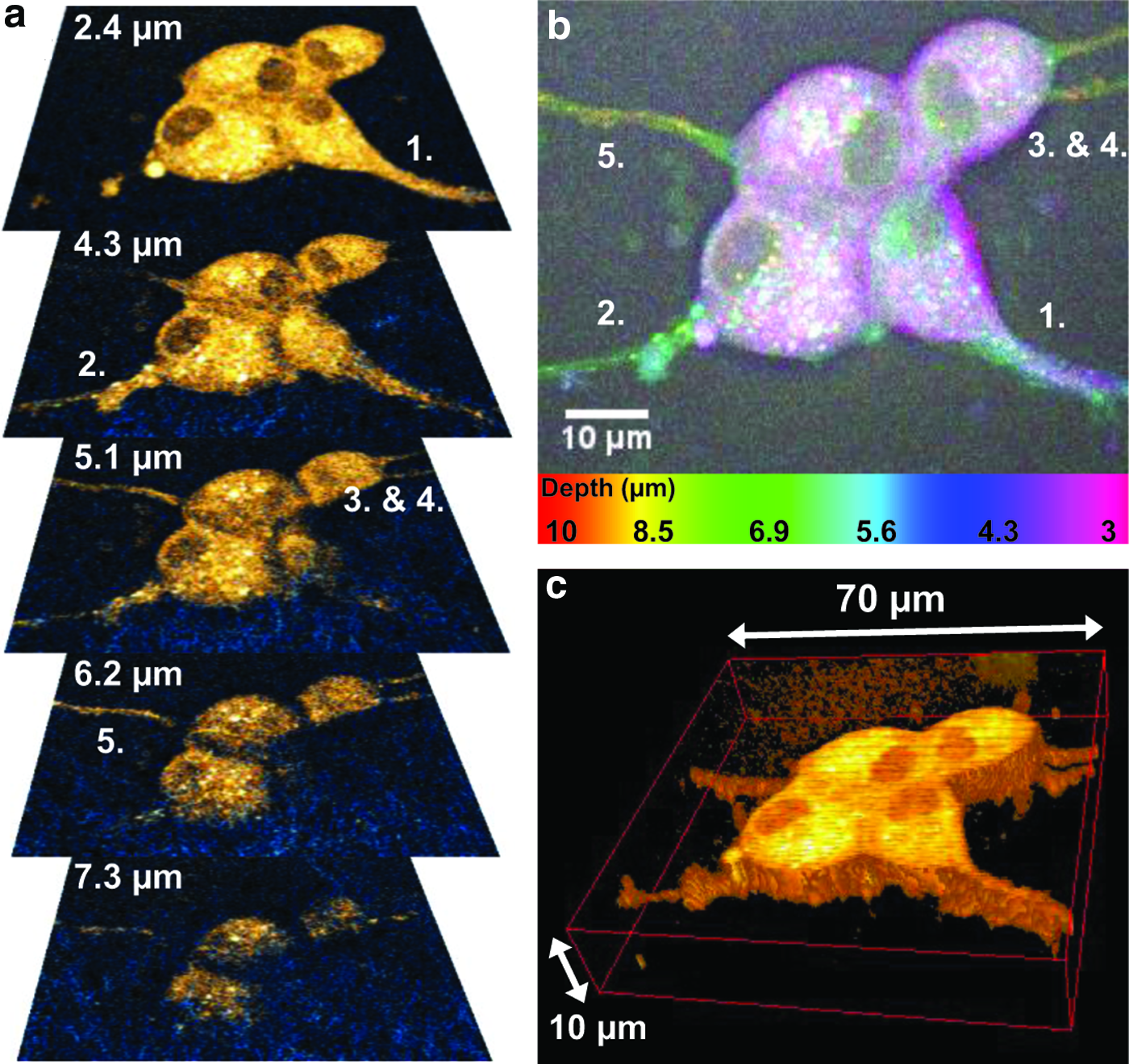
An ensemble of PC12 cells after 8 days of growth on a collagen-coated TMAHP-treated scaffold with NGF added.
Figure 5 illustrates how neurites have migrated through the scaffold (collagen-coated TMAHP cellulose) and successfully connected to other ensembles of PC12 cells, forming a simple neuronal network. Different planes of CARS and SHG microscopy overlay images illustrate that the neurites propagate at different depths in the scaffold. At a depth of 2 μm, neurite 1 can be distinguished, linking the two most superficial cell clusters. At depth 3.3 μm, a twin set of neurites (2 and 3) extending from the central cell can be identified and at 4.3 μm, a >35 μm long (neurite 4) and a branched collection (neurite 5) of neurites can be observed. Neurites 6 and 7 of the deepest lying cells can accordingly be found deeper into the scaffold at depths 7–8 μm. This 3D arrangement of neurites is further illustrated in the depth of color coded image in Figure 5b, where the different neurites appear in colors corresponding to their depths; neurite 1 and associated cells in magenta, neurites 2–5 in blue/green, and neurites 6–7 in yellow/red. The full 3D arrangement of the network is outlined in the CARS volume view in Figure 5c. A zoom-in depth projection CARS/SHG overlay image of the central cell acting as the hub in the network is shown in Figure 5d. Neurites 2–7 can here all clearly be distinguished as well as the arrangement of surrounding nanocellulose fibrils, in particular those associated with neurite 5 (arrow in Fig. 5d). A close-up of the 3D arrangement of the neurites of the central cell is shown in the volume view in Figure 5e. The corresponding areas of the zoom-in images are indicated by dashed squares in the overview images in Figure 5b and c.

Multiple clusters of PC12 cells, grown for 8 days on a collagen-coated TMAHP-treated scaffold with NGF added, are connected by neurites expanding in different planes as observed in a series of CARS images
Lengths of neurite extensions were analyzed from CARS microscopy images measured on samples for which NGF was added during cultivation. For collagen-coated TMAHP-treated scaffolds, 35 neurites were distributed over 22 cells with between 1 and 4 neurites each (cf. Fig. 5). The average neurite length was 20 μm with a standard deviation of 11 μm and the values ranged from 7 to 54 μm (cf. Fig. 5). Smaller data sets for scaffolds with TMAHP treatment only, collagen-coating only, and TMAHP-treatment with laminin-coating showed average neurite lengths of 20, 26, and 34 μm, respectively. The value obtained for laminin coating is higher than the others but is based on measurements of too few neurites to confirm a significant difference. For comparison, PC12 neurite growth on collagen gels has been investigated by Baldwin et al. 11 and neurite lengths in the range 50–80 μm were observed after 4 days of growth. The results obtained for nanocellulose therefore seem promising for this material as a scaffold for neuronal cells; however, neurite growth depends on cultivation conditions 11 as well as mechanical properties of the scaffold material.32,33 Thus, additional investigations are required to clarify possible differences in neurite growth between different preparations of the nanocellulose and in comparison with other scaffold materials.
The importance of reliable methods for neurite visualization and quantification during growth in 3D systems has been pointed out by Arien-Zakay et al. 10 who developed PC12 cells expressing green fluorescent protein (GFP) to allow for neurite monitoring by fluorescence microscopy. 10 The results presented in Figures 4 and 5 clearly show that CARS microscopy is able to provide this for standard PC12 cells or other neuronal cell models, without such special preparations making it a valuable method to support neuronal tissue engineering. Refined image analysis methods, such as those presented by Arien-Zakay et al. 10 for automatic evaluation of neurite extension and branching, could be used to extract data for larger sets of CARS images.
Conclusion
TMAHP modification of nanocellulose fibrils promotes attachment and proliferation of PC12 cells on nanocellulose scaffolds, potentially due to a more efficient nonspecific protein adsorption from the medium. Protein coating per se does not improve cell proliferation significantly, but promotes more active interaction between the cells and the scaffold components, as well as cell migration and integration. Cell integration appears to take place by active remodeling of the scaffold to a depth of one to two cell layers. Deeper integration was not observed irrespective of scaffold preparation procedure, most likely due to the fact that nanocellulose fibrils cannot be protolytically degraded, which is the natural route for 3D cell integration in tissues. To promote deeper cell integration, we propose to integrate protolytically degradable components in the scaffolds or use macroporous nanocellulose. From 3D imaging of neurites, we conclude that these have a capacity to integrate deeper than the cell bodies within the scaffold due to their fine dimensions and an amoeba-like migration pattern. Further developments in chemical modification and scaffold architecture open up for improved nanocellulose designs more suitable for neuronal cell growth. In addition, the combined approach of nanocellulose scaffolds and label-free high-resolution CARS/SHG microscopy provides a novel platform for neurophysiological investigations and opens up for functional imaging of neuronal networks in future time.
Footnotes
Acknowledgments
The financial support of the Swedish Research Council and the Wallenberg Foundation is gratefully acknowledged. Gatenholm and Enejder express their gratitude to Chalmers Area of Advance in Material Science.
Disclosure Statement
No competing financial interest exists.