
Abstract
The dental follicle is a source of dental follicle stem cells (DFCs), which have the potential to differentiate into the periodontal lineage. DFCs therefore are of value in dental tissue engineering. The purpose of this study was to evaluate the effect of growth factor type and concentration on DFC differentiation into periodontal specific lineages. DFCs were isolated from the human dental follicle and characterized for the expression of mesenchymal markers. The cells were positive for CD-73, CD-44, and CD-90; and negative for CD-33, CD-34, and CD-45. The expression of CD-29 and CD-31 was almost negligible. The cells also expressed periodontal ligament and cementum markers such as periodontal ligament-associated protein-1 (PLAP-1), fibroblast growth factor-2 (FGF-2), and cementum protein-1 (CEMP-1), however, the expression of osteoblast markers was absent. Further, the DFCs were cultured in three different induction medium to analyze the osteoblastic, fibroblastic, and cementoblastic differentiation. Runt-related transcription factor 2 (RUNX-2), alkaline phosphatase (ALP) activity, alizarin staining, calcium quantification, collagen type-1 (Col-1), and osteopontin (OPN) expression confirmed the osteoblastic differentiation of DFCs. DFCs cultured in recombinant human FGF-2 (rhFGF-2) containing medium showed enhanced PLAP-1, FGF-2, and COL-1 expression with increasing concentration of rhFGF-2 which thereby confirmed periodontal ligament fibroblastic differentiation. Similarly, DFCs cultured in recombinant human cementum protein-1 (rhCEMP-1) containing medium showed enhanced bone sialoprotein-2 (BSP-2), CEMP-1, and COL-1 expression with respect to rhCEMP-1 which confirmed cementoblastic differentiation. The expression of osteoblast, fibroblast, and cementoblast-related genes of DFCs cultured in induction medium was enhanced in comparison to DFCs cultured in noninduction medium. Thus, growth factor-dependent differentiation of DFCs into periodontal specific lineages was proved by quantitative analysis.
Introduction
P
In the current scenario, diverse periodontal regenerative strategies are being exploited for a successful outcome. These include application of biomaterials in the form of resorbable or nonresorbable barrier membranes for guided tissue regeneration,2,3 growth factors,4–6 cell-based therapy,7,8 grafts include autografts from intraoral and extraoral sites, 9 allografts such as freeze-dried and demineralized matrices,9,10 xenografts9,10 and alloplastic graft materials11,12 such as hydroxyapatite, bioglass, tricalcium phosphate, or a combination of two or more factors.10,13
Cell-based therapy, an obligatory element for successful tissue engineering, is gaining widespread interest in the field of periodontal regeneration. 7 Till date, different cell types such as mesenchymal stem cells of bone marrow, umbilical cord, or adipose-derived origin and dental stem cells such as periodontal ligament cells, dental follicle cells, stem cells from apical papilla, and stem cells from exfoliated deciduous tooth have been utilized to regenerate periodontal tissues.7,14 Dental stem cells are formed by ectomesenchymal tissue, derived from the neural crest during embryonic development unlike the bone marrow and umbilical cord stem cells, which are derived from mesoderm. 15 Periodontium and tooth, form a part of craniofacial structures that are derived from the ectomesenchyme. Owing to these advantages, dental stem cells may serve as a better choice for the repair of neural crest-derived tissues like periodontium. 16 Among the available sources of dental stem cells, dental follicle stem cells (DFCs) captured our interest owing to its location, ease of harvesting, and the ability to differentiate into multiple cell types (multipotency). 7
The dental follicle or sac is a loose, organized, mesenchymal tissue of the developing tooth germ. Internally, it lies in direct contact with the enamel organ whereas it is separated externally from the alveolar bone by a loose, vascular connective tissue, that is, dental papilla. It regulates bone resorption by furnishing osteoclasts or releasing necessary cytokines and growth factors followed by osteogenesis leading to new bone formation. Hence, dental follicle plays a vital role in tooth eruption.17,18 Stem cells and progenitor or precursor cells residing in the dental follicle were reported to differentiate into three cell types namely osteoblast, periodontal ligament fibroblast (PDLF), and cementoblast, which result in the formation of the three distinct periodontal tissues—the cementum, periodontal ligament, and the alveolar bone.7,19–21 This study was aimed at understanding the effect of growth factor concentration on the differentiation of DFCs.
Materials and Methods
Isolation and culture of DFCs
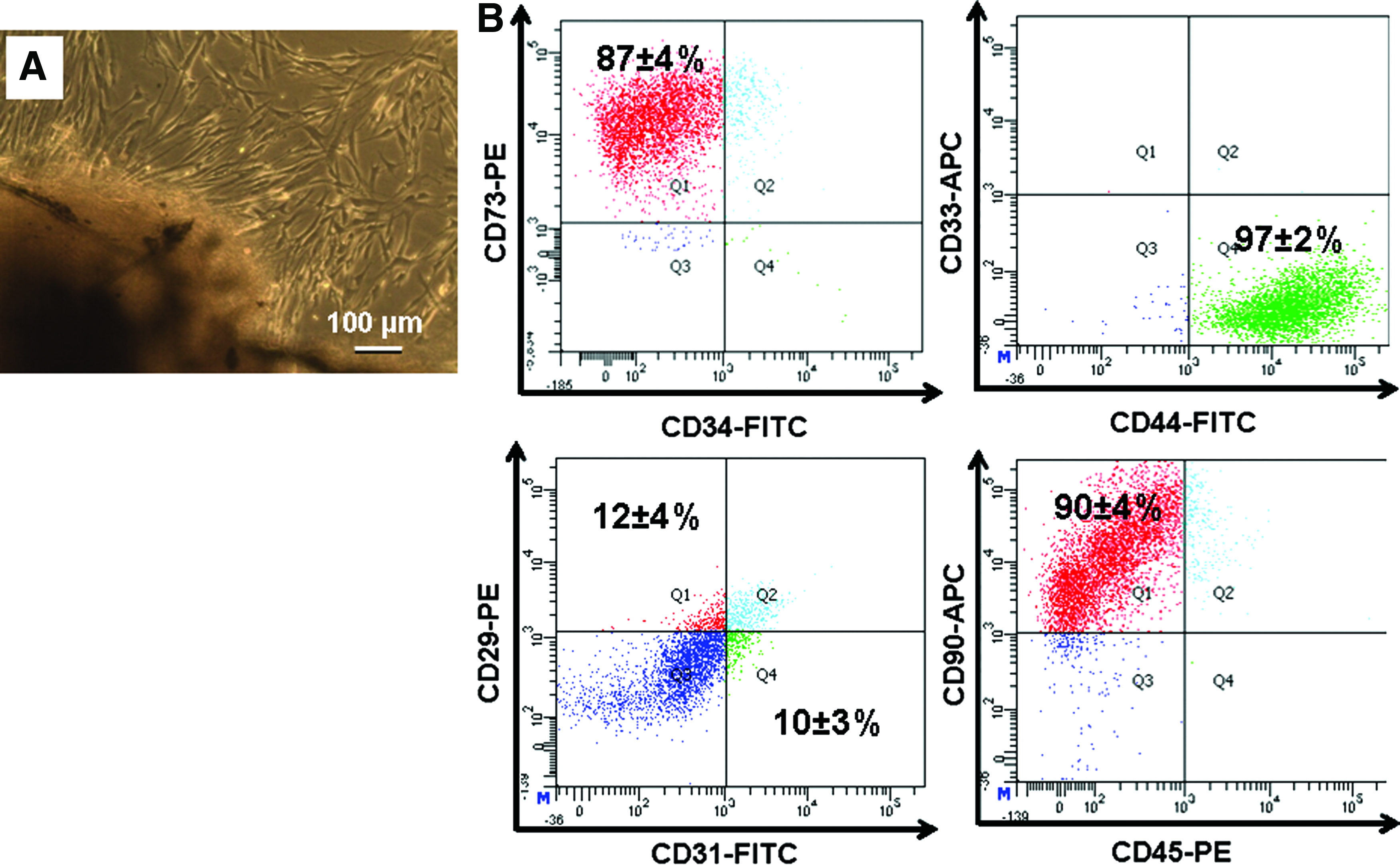
Healthy human third molars surgically removed from 18- to 24-year-old patients for orthodontic reasons were obtained with prior informed consent (approved by the Institutional Ethics Committee dated 14.11.2011). Dental follicles attached to the tooth surface were collected in sterile falcon tubes containing phosphate-buffered saline (PBS) and 100 U/L of antibiotic/antimycotic (Anti-Anti) solution (Gibco, Invitrogen). The follicles were then separated from the tooth surface, thoroughly washed with PBS containing 100 U/L of Anti-Anti solution, and minced using a sterile scalpel. Further, the minced tissue was digested in 0.2% collagenase (Gibco, Invitrogen) for 1 h at 37°C. After the desired incubation, the digested tissue was centrifuged at 1500 rpm/5 min. The supernatant was discarded; the follicle tissue explant was seeded into T-25 culture flask containing 2 mL of Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich) with 10% fetal bovine serum (FBS; Gibco, Invitrogen) and 100 U/L of Anti-Anti and incubated at 37°C, 5% CO2 in a humidified atmosphere. After 24 h, the medium was replaced with fresh 10% DMEM. Thereafter, medium was changed once in every 3 days. Upon confluency, the cells were passaged into new flasks. Optical microscopic images of the isolated DFCs were acquired (Leica Optical).
Immunophenotypic analysis of isolated DFCs
1×107 DFCs from third passage were harvested, wherein the cell pellet was suspended in 0.7 mL of PBS and incubated with CD-29 (integrin β1; PE conjugated), CD-31 (platelet-endothelial cell adhesion molecule-1; FITC conjugated), CD-34 (hematopoietic progenitor cell antigen; FITC conjugated), CD-73 (ecto-5′-nucleotidase; PE conjugated), CD-33 (sialic acid-binding immunoglobulin-like lectin 3 [SIGLEC3]; a surface marker for very early bone marrow-derived hematopoietic stem cells; APC conjugated), CD-44 (hyaluronate/lymphocyte homing-associated cell adhesion molecule-HCAM; FITC conjugated), CD-45 (protein tyrosine phosphatase, receptor type, C/PTPRC/leucocyte common antigen/cell marker of hematopoietic origin; PE conjugated), and CD-90 (Thy-1/Thy-1.1; APC conjugated) for 30 min. All the antibodies were purchased from BD Biosciences. Flow-cytometry analysis was carried out using the Beckman Coulter Cytomics FC 500 MPL system (Beckman Coulter).
Expression of PLAP-1, FGF-2, CEMP-1, and RUNX-2 proteins in DFCs without stimulation
For immunocytochemical staining, DFCs (passage 3–6) were seeded at a density of 4×104 cells per well in a 24-well plate. Following an incubation period of 24 h, the cells were fixed with 4% paraformaldehyde (PFA; Sigma-Aldrich) for 15 min and permeabilized with 0.2% Triton X-100 (Sigma-Aldrich) in PBS for 5 min. Subsequent to background inhibition in 0.1% FBS, cells were labeled with anti-human periodontal ligament associated protein-1 (PLAP-1) IgG goat polyclonal antibody (Santa Cruz Biotechnology) and incubated at 4°C overnight. Following incubation, the cells were thoroughly washed with PBS and incubated with FITC-labeled donkey anti-goat IgG antibody (Santa Cruz Biotechnology) at room temperature in dark for 1 h. The cells were then washed with PBS and analyzed using fluorescence microscope. The same protocol was followed for labeling cells with other primary antibodies such as anti-human fibroblast growth factor-2 (FGF-2) IgG goat polyclonal antibody (Santa Cruz Biotechnology), anti-human cementum protein-1 (CEMP-1) IgG goat polyclonal antibody (Santa Cruz Biotechnology), and anti-human Runt-related transcription factor 2 (RUNX-2) IgG mouse polyclonal antibody (Santa Cruz Biotechnology).
For flow cytometry analysis, DFCs (passage 3–6) at a density of 2×105 cells, suspended in PBS in 2 mL eppendorf tubes were fixed with 4% PFA for 15 min. Following PBS wash, the cells were permeabilized with 0.2% Triton X-100 in PBS for 5 min and blocked with 0.1% FBS. Anti-PLAP-1, anti-FGF-2, anti-CEMP-1, and anti-RUNX-2 antibodies were added to the respective tubes, incubated at room temperature for 1 h followed by FITC-labeled secondary antibody incubation at room temperature in dark for 45 min. The percentage of cells positive for anti-PLAP-1, anti-FGF-2, anti-CEMP-1, and anti-RUNX-2 antibodies was analyzed using Beckman Coulter Cytomics FC 500 MPL system (Beckman Coulter).
Growth factors released by DFCs
The growth factor analysis was carried out as reported earlier. 22 Briefly, DFCs (passage 3–6) were seeded at a density of 2×104 cells per well in a 24-well plate containing 10% DMEM. After 24 h of incubation, the cells were washed with PBS; replenished with 0.1% DMEM and cultured for 48 h. Following incubation, the supernatant, that is, cell-free medium was harvested and stored at −80°C until further analysis. To determine the secretion of growth factors such as vascular endothelial growth factor (VEGF, RayBio® Human VEGF ELISA Kit; RayBiotech), FGF-2 (RayBio® Human FGF-2 ELISA Kit; RayBiotech), transforming growth factor-beta 1 (TGF-β1, RayBio® Human TGF-β1 ELISA Kit; RayBiotech), and CEMP-1 (Human CEMP-1 ELISA Kit; CUSABIO), the collected supernatant or cell-free medium of DFCs was analyzed through enzyme-linked immunosorbent assay (ELISA) according to the manufacturer's protocol.
Osteogenic proliferation and differentiation
Total DNA content of DFCs cultured in osteogenic induction medium (OM) was quantified using the PicoGreen dsDNA assay (Invitrogen). Briefly, DFCs were seeded at the density of 5×104 cells per well in a 12-well plate containing OM [10% DMEM containing 10 nM dexamethasone (Dex; Sigma-Aldrich), 10 mM β-glycerophosphate (β-GP; Fluka), and 50 μg/mL ascorbic acid (AA,
RUNX-2 expression
DFCs (passage 3–6) were seeded at a density of 4×104 cells per well in a 24-well plate containing 10% DMEM. At 80% confluency, DFCs were replenished with OM. The medium was changed once in every 3 days. After 5 days of culture, cells were fixed with 4% PFA for 15 min and permeabilized with 0.2% Triton X-100 in PBS for 5 min. After background inhibition in 0.1% FBS, cells were labeled with anti-human RUNX-2 IgG mouse polyclonal antibody, and incubated at room temperature for 1 h. After the desired incubation period, the cells were thoroughly washed with PBS and incubated with FITC-labeled donkey anti-mouse IgG antibody (Santa Cruz Biotechnology) for 45 min at room temperature in dark. The cells were then washed with PBS and analyzed using flow cytometry and fluorescence microscope.
Alkaline phosphatase activity
Alkaline phosphatase (ALP) activity was measured using a colorimetric endpoint assay that utilizes p-nitrophenol phosphate (PNPP; Sigma-Aldrich) as the substrate. DFCs (passage 3–6) were seeded at a density of 4×104 cells per well in a 24-well plate containing 10% DMEM. At 80% confluency, DFCs were replenished with OM and cultured for a period of 21 days. The medium was changed once in every 3 days. After the predetermined incubation time of 7, 14, and 21 days, the cells were washed with PBS and incubated with 1% Triton X-100 for 2 h to obtain cell lysates, which were ultrasonicated for 30 min. The supernatant thus obtained was incubated with PNPP substrate in the presence of glycine buffer (to maintain an alkaline pH) for 30 min in dark at 37°C. The ALP activity corresponds to spectrophotometric endpoint, which determines the conversion of colorless PNPP substrate into colored p-nitrophenol. The absorbance was measured at 405 and 490 nm. Standards were prepared from ALP control (Himedia). The DFCs cultured in OM were compared with cells in non-osteogenic medium, that is, 10% DMEM (non-OM). All the samples were analyzed in triplicates.
Mineralization
DFCs (passage 3–6) were seeded at a density of 4×104 cells per well in a 24-well plate containing 10% DMEM. At 80% confluency, DFCs were replenished with OM and cultured for a period of 28 days. The medium was changed once in every 3 days. After the predetermined incubation time of 14, 21, and 28 days, the cells were rinsed with deionized water and stained with 2% alizarin red solution (pH=4.2) and incubated for 5–10 min at room temperature. Alizarin was aspirated and the wells were thoroughly washed with deionized water at least three to four times before observation. Calcium deposits can be visualized by their intense red color. Images were acquired using an optical microscope. The DFCs cultured in OM were compared with cells in non-OM. All the samples were analyzed in triplicates.
To quantify the calcium content, 500 μL of 1 M acetic acid (Merck Millipore) was added to 24-well plate containing cells and incubated overnight at 4°C with gentle shaking for the extraction of the mineralized content. A volume of 300 μL of calcium assay reagent (Arsenazo III; Coral Clinical Systems) was added to 20 μL of extracted solution in triplicate in 96-well plates. To generate a standard curve, serial dilutions of calcium chloride (CaCl2; Merck Millipore) were prepared (0–50 μg/mL). Absorbance was read at 650 nm using a spectrophotometric plate reader. The calcium deposition at each incubation time was expressed as μg/mL. The DFCs cultured in OM were compared with cells in non-OM. All the samples were analyzed in triplicates.
Expression of COL-1 and OPN proteins
DFCs were seeded at the density of 4×104 cells per well in a 12-well plate containing 10% DMEM. At 80% confluency, DFCs were replenished with OM. Medium was changed once in every 3 days. After 14 and 21 days of culture, the cells were fixed with 4% PFA for 15 min and permeabilized with 0.2% Triton X-100 in PBS for 5 min. After background inhibition in 0.1% FBS, cells were labeled with anti-collagen type-1 (COL-1; Antibodies Online) and anti-osteopontin (OPN; Santa Cruz Biotechnology) mouse-IgG antibodies, and incubated at 4°C overnight. Following incubation, the cells were thoroughly washed with PBS and incubated with FITC-labeled donkey anti-mouse IgG antibody (Santa Cruz Biotechnology) at room temperature in dark for 1 h. The cells were then washed and analyzed using fluorescence microscope.
Fibroblastic proliferation and differentiation
Total DNA content of DFCs cultured in fibroblast induction medium (FM) (10% DMEM containing recombinant human FGF-2 [rhFGF-2, 20 ng/mL; Gold Biotechnology] and 50 μg/mL AA) was quantified using the PicoGreen dsDNA assay (Invitrogen) similar to osteogenic proliferation as indicated before. All the samples were analyzed in triplicates. The DNA concentration was calculated based on the standard curve.
Expression of FGF-2 and PLAP-1 proteins
DFCs (passage 3–6) were seeded at a density of 4×104 cells per well in a 24-well plate containing 10% DMEM. At 80% confluency, DFCs were replenished with FM containing three different concentrations of rhFGF-2 (2, 10, and 20 ng/mL) and 50 μg/mL AA. The medium was changed once in every 2 days. Following 10 days of culture, cells were fixed with 4% PFA for 15 min and permeabilized with 0.2% Triton X-100 in PBS for 5 min. Subsequent to background inhibition in 0.1% FBS, cells were labeled with anti-FGF-2 and anti-PLAP-1 antibodies and incubated at room temperature for 1 h. Following incubation, the cells were thoroughly washed with PBS and incubated with FITC-labeled donkey anti-goat IgG antibody at room temperature in dark for 45 min. The cells were washed and analyzed using flow cytometry.
Expression of COL-1 protein
DFCs (passage 3–6) were seeded at a density of 4×104 cells per well in a 24-well plate containing 10% DMEM. At 80% confluency, DFCs were replenished with FM. The medium was changed once in every 2 days. Following 14 and 21 days of culture, the cells were fixed with 4% PFA for 15 min and permeabilized with 0.2% Triton X-100 in PBS for 5 min. Subsequent to background inhibition in 0.1% FBS, cells were labeled with anti-COL-1 antibody and incubated at 4°C overnight. Following incubation, the cells were thoroughly washed with PBS and incubated with FITC-labeled donkey anti-mouse IgG antibody at room temperature in dark for 1 h. The cells were then washed and analyzed using fluorescence microscope.
Cementogenic proliferation and differentiation
Total DNA content of DFCs cultured in cementogenic induction medium (CM; 10% DMEM containing recombinant human cementum protein-1 [rhCEMP-1; provided by H. Arzate] at a concentration of 2 μg/mL; 10 mM β-GP and 50 μg/mL AA) was quantified using the PicoGreen dsDNA assay (Invitrogen) similar to osteogenic proliferation as indicated before. All the samples were analyzed in triplicates. The DNA concentration was calculated based on the standard curve.
Expression of CEMP-1 and BSP-2 proteins
DFCs (passage 3–6) were seeded at a density of 4×104 cells per well in a 24-well plate containing 10% DMEM. At 80% confluency, DFCs were replenished with CM containing three different concentrations of rhCEMP-1 (100, 500 ng/mL, and 2 μg/mL), 10 mM β-GP, and 50 μg/mL AA. The medium was changed once in every 2 days. Following 10 days of culture, cells were fixed with 4% PFA for 15 min and permeabilized with 0.2% Triton X-100 in PBS for 5 min. Subsequent to background inhibition in 0.1% FBS, cells were labeled with anti-CEMP-1 and anti-bone sialoprotein-2 (BSP-2) antibodies and incubated at room temperature for 1 h. Following incubation, the cells were thoroughly washed with PBS and incubated with FITC-labeled donkey anti-goat IgG antibody at room temperature in dark for 45 min. The cells were washed and analyzed using flow cytometry.
Expression of COL-1 and OPN proteins
DFCs (passage 3–6) were seeded at a density of 4×104 cells per well in a 24-well plate containing 10% DMEM. At 80% confluency, DFCs were replenished with CM. The medium was changed once in every 2 days. Following 14 and 21 days of culture, the cells were fixed with 4% PFA for 15 min and permeabilized with 0.2% Triton X-100 in PBS for 5 min. Subsequent to background inhibition in 0.1% FBS, cells were labeled with anti-COL-I and anti-OPN antibodies and incubated at 4°C overnight. Following incubation, the cells were thoroughly washed with PBS and incubated with FITC-labeled donkey anti-mouse IgG antibody at room temperature in dark for 1 h. The cells were then washed and analyzed using fluorescence microscope.
Quantitative real-time polymerase chain reaction
Total RNA extraction from DFCs cultured in 10% DMEM, OM, FM, and CM, was performed using TRIzol® Reagent (Invitrogen) according to the manufacturer's instructions. Complementary DNA (cDNA) was synthesized using Superscript III reverse transcriptase (Invitrogen) from 1 μg of total RNA following the manufacturer's instructions. Quantitative real-time polymerase chain reaction was performed on Applied Biosystems 7900HT Fast Real-Time PCR System (Applied Biosystems) using SYBR Green detection reagent (Ambion). Amplification conditions were 95°C for 10 min for denaturation, 40 cycles of 95°C for 15 s, and 60°C for 1 min followed by a dissociation curve. Primer sequences were as follows: Glyceraldehyde 3-phosphate dehydrogenase (GAPDH), forward 5′-ATGGGGAAGGTGAAGGTCG-3′ and reverse 5′-GGGGTCATTGATGGCAACAATA-3′; ALP, forward 5′-ATGGGATGGGTGTCTCCACA-3′ and reverse 5′-CCCACCCCGCTATTCCAAAC-3′; osteocalcin (OCN), forward 5′-CACTCCTCGCCCTATTGGC-3′ and reverse 5′-CCCTCCTGCTTGGACACAAAG-3′; OPN, forward 5′-TAGCTTGGCTTATGGACTGAGG-3′ and reverse 5′-AGACTCACCGCTCTTCATGTG-3′; COL-1, forward 5′-ATGACTATGAGTATGGGGAAGCA-3′ and reverse 5′-TGGGTCCCTCTGTTACACTTT-3′; CEMP-1, forward 5′-ACCAAGAGTGCTTCCCCCACAC-3′ and reverse 5′-CCCATGTGGCAAACACGGGC-3′ FGFR-2, forward 5′-GGATGGGGTGCACTTGGGGC-3′ and reverse 5′-TCTCTAGGCGTGGCGCCCTT-3′; and PLAP-1, forward 5′-AGGGTGCAGCCACACCAGGA-3′ and reverse 5′-ACCCCTTCAAATGCCCCTGGC-3′ Data were analyzed using ABI SDS v2.4 software (Applied Biosystems). All samples were assayed in triplicates. The mean cycle threshold (Ct) value of each target gene was normalized against the Ct value of GAPDH. The relative expression was calculated using the following formula: 2−(normalized average Ct).
Statistical analysis
All results were obtained from triplicate samples. Data were expressed as the mean±standard deviation. One-way analysis of variance was used to compare differences in the DNA concentration and ALP activity at different time points followed by Student's two-tailed t-test. Values at *p≤0.05, **p≤0.01, and ***p≤0.001 were considered to be statistically significant.
Results
Immunophenotypic analysis of isolated DFCs
DFCs are gaining widespread interest in stem cell-based therapy owing to ease of harvesting from extracted tooth considered as medical waste, plastic adherence, self-renewal, and differentiation ability similar to umbilical cord stem cells. 15 To confirm the existence of mesenchymal stem cells in the dental follicle, immunophenotypic analysis was carried out. DFCs were expanded from human dental follicle, harvested from extracted normal human third molars. Optical microscopic images of the isolated cells exhibited spindle-shaped fibroblast-like morphology (Fig. 1A). Cell surface antigen/immunophenotypic analysis was performed using flow cytometry (Fig. 1B). Cells showed 90% positivity toward CD-73, 99% positivity toward CD-44, and 94% positivity toward CD-90. The expression of CD-29 and CD-31 was limited to 12% and 10% respectively. The cells were negative to CD-33, CD-34, and CD-45.
Expression of PLAP-1, FGF-2, CEMP-1, and RUNX-2 proteins in DFCs without stimulation
Although majority of DFCs express mesenchymal stem cell markers, reports have suggested the existence of PDLF and cementoblast progenitor population in the DFCs. To confirm, immunocytochemical analysis was carried out, which showed the localization of PLAP-1, FGF-2, and CEMP-1 proteins in the cytoplasmic region of DFCs. No expression of RUNX-2 protein was observed (Fig. 2). Flow cytometry analysis also confirmed the expression of PLAP-1 (16%±4%), FGF-2 (40%±6%), and CEMP-1 (23%±6%) proteins. However, the expression of RUNX-2 was almost negligible (4%±1%) (Fig. 3).
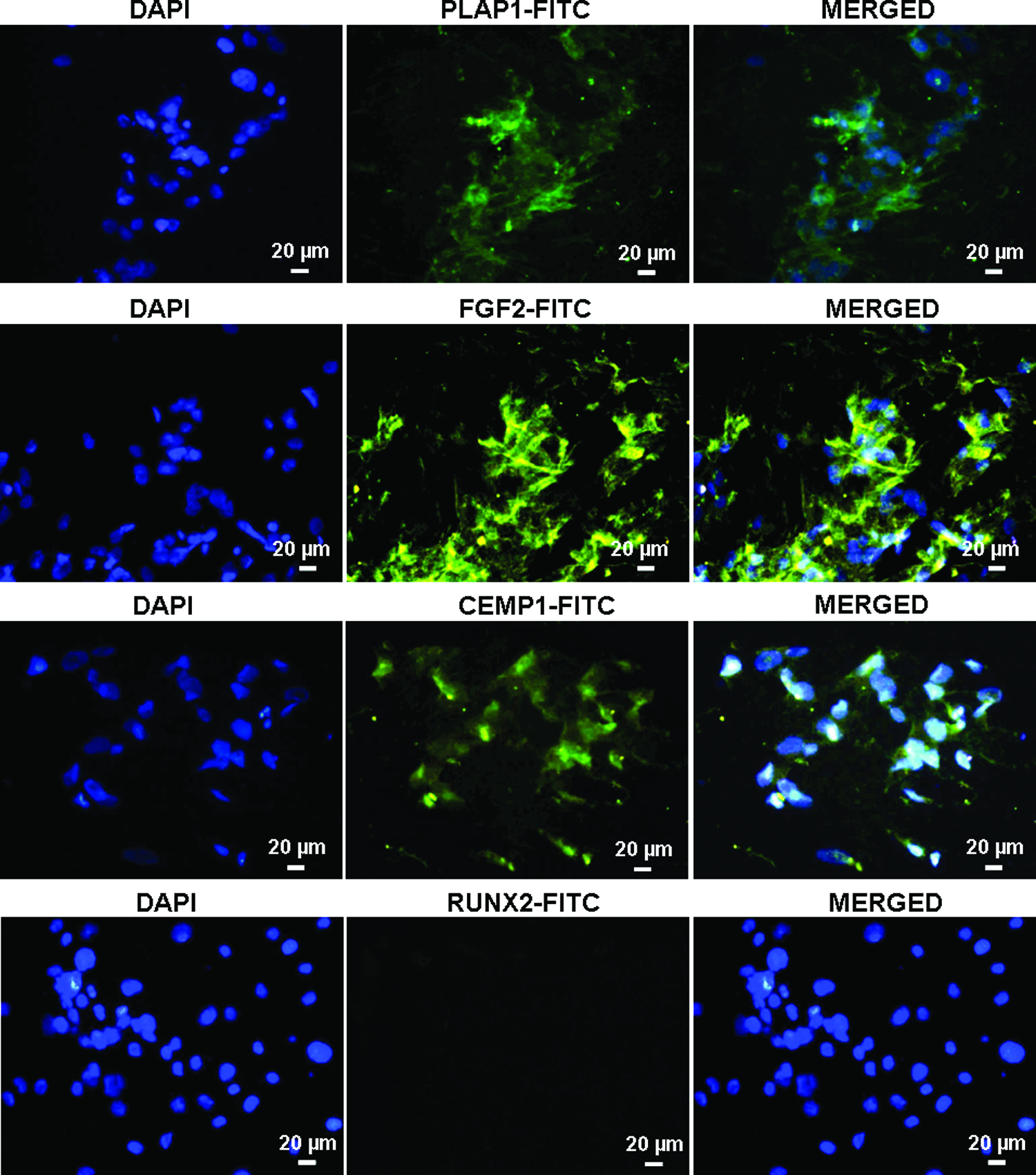
Immunocytochemical analysis of periodontal ligament-associated protein-1 (PLAP-1), fibroblast growth factor-2 (FGF-2), cementum protein-1 (CEMP-1), and Runt-related transcription factor 2 (RUNX-2) in DFCs. Nucleus was stained with DAPI. Color images available online at
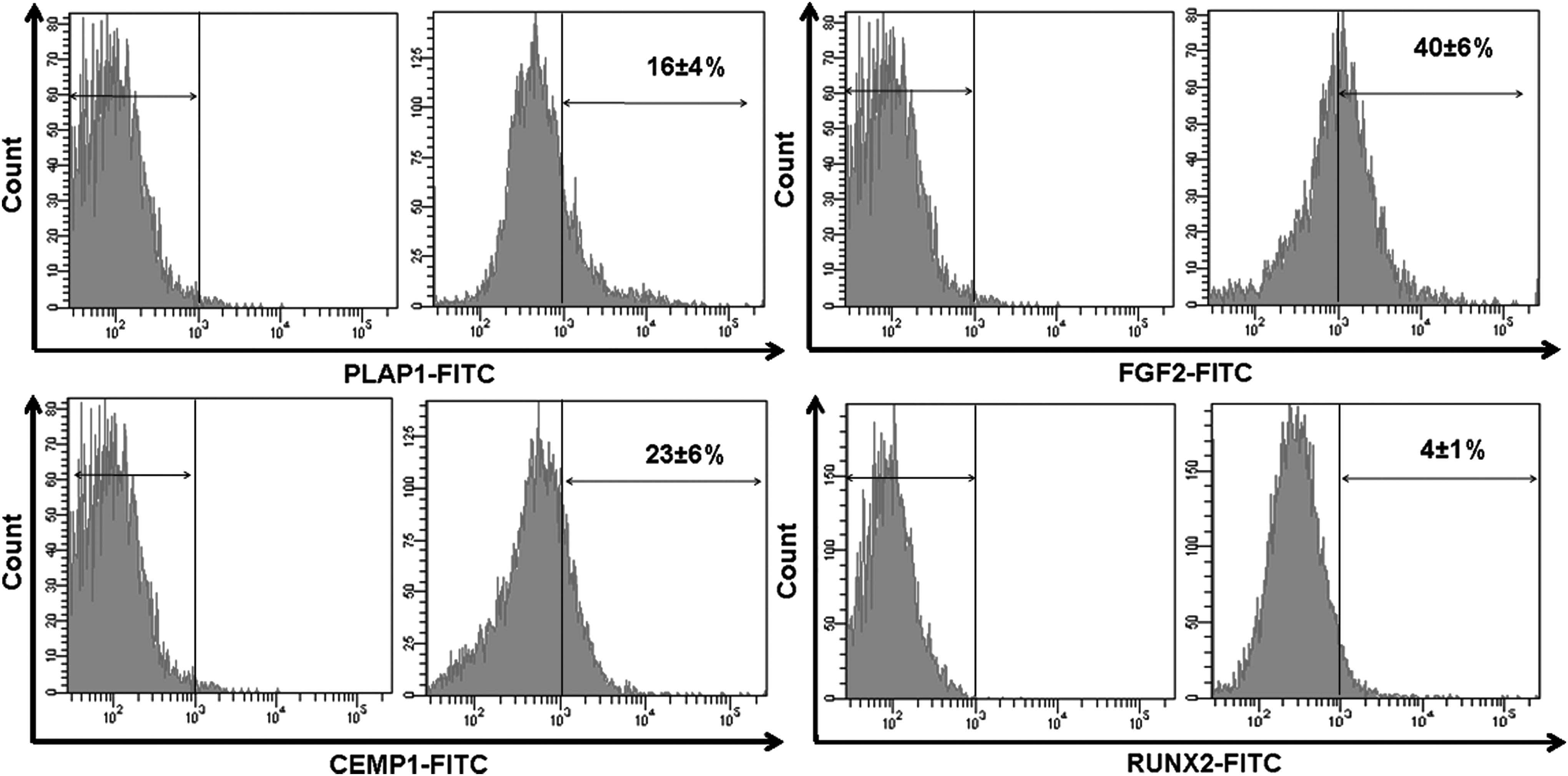
Flow cytometry analysis of PLAP-1, FGF-2, CEMP-1, and RUNX-2 in DFCs.
Growth factors released by DFCs
Reports have indicated that mesenchymal stem cells secrete growth factors and cytokines that possess paracrine effects and may assist tissue regeneration, angiogenesis, and wound healing under in vivo conditions.23,24 To confirm the presence of paracrine factors in DF cell culture supernatant, ELISA was performed. The estimated concentration after 48 h of incubation was 1600±123 pg/mL for VEGF, 320±28 pg/mL for TGF-β1, 690±15 pg/mL for FGF-2, and 300±42 pg/mL for CEMP-1 respectively.
Osteogenic proliferation and differentiation
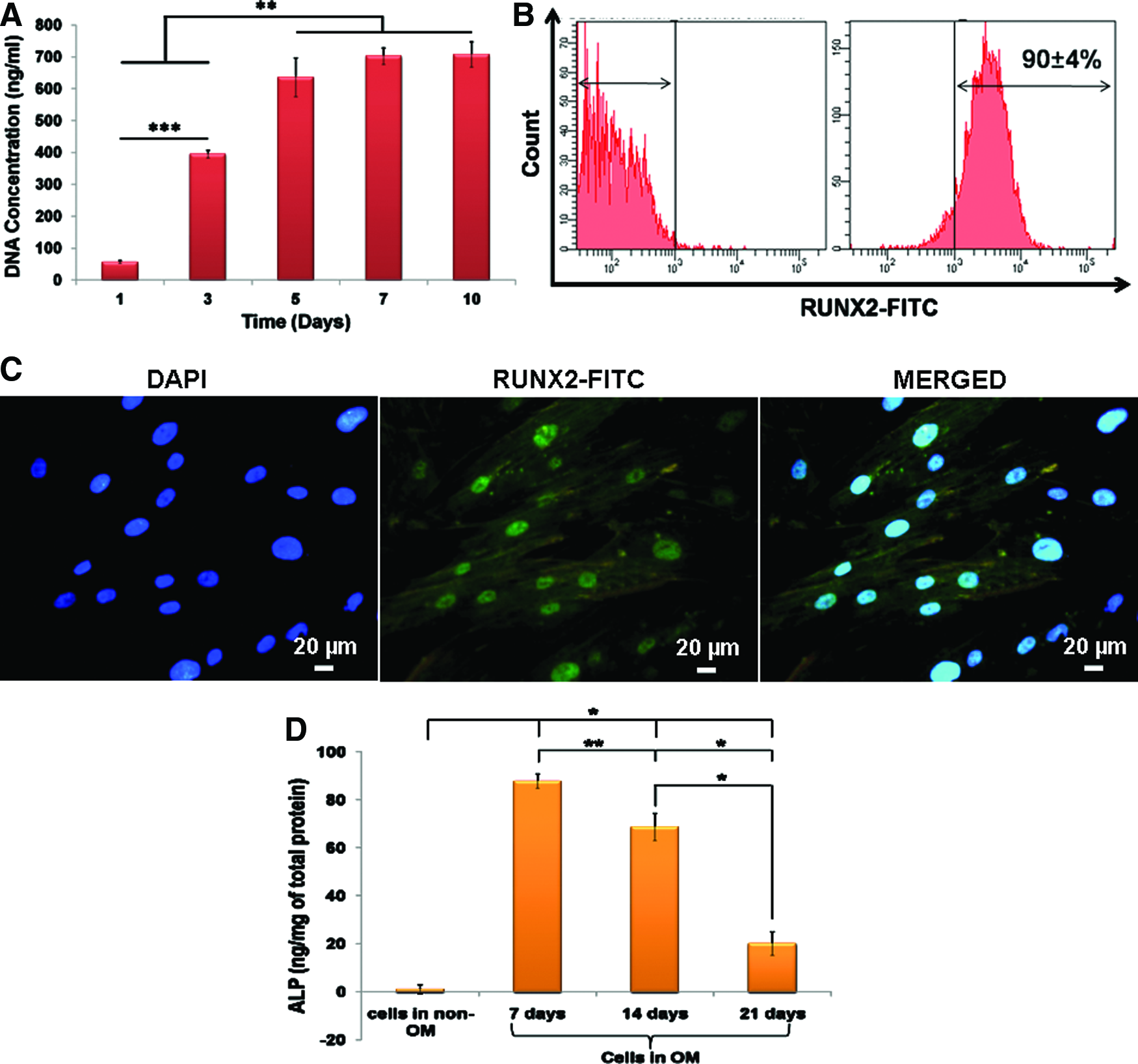
The course of osteoblast differentiation in DFCs was analyzed through the following experiments. The DNA concentration corresponds to cellular proliferation, which was determined by PicoGreen assay. DFCs cultured in OM showed a significantly higher DNA concentration at day 5 in comparison to day 1 and 3 (Fig. 4A). Thereafter, no significant increase in DNA concentration was observed.
RUNX-2 expression
To analyze the RUNX-2 expression, DFCs were cultured in OM for 5 days. Flow cytometry analysis showed 90%±4% positivity toward anti-RUNX-2 antibody (Fig. 4B). Immunocytochemical analysis also confirmed the expression wherein RUNX-2 was localized to the nuclei of DFCs (Fig. 4C).
ALP activity
The ALP activity of DFCs cultured in OM peaked at 7 days followed by a decline in its activity thereafter (Fig. 4D). In addition, the DFCs cultured in OM showed a significantly higher ALP activity in comparison with the cells cultured in non-OM.
Mineralization
The formation of mineralized calcium phosphate deposits over a period of 14, 21, and 28 days in culture, was confirmed through alizarin red staining (Fig. 5A–D). The intensity of mineralization increased with the increase in culture time. No deposits were observed on DFCs cultured in non-OM. Mineralization observed through alizarin staining was further confirmed by calcium quantification. Calcium concentration was determined by Arsenazo method wherein the calcium levels after 28 days' culture in OM was measured as 70±10 μg/mL in contrast to very minimal calcium detected in non-osteogenic culture (Fig. 5E).

Alizarin red-stained images of DFCs cultured in
Expression of COL-1 and OPN proteins
To analyze the expression of COL-1 and OCN proteins, DFCs were cultured in OM for 14 and 21 days. On immunocytochemical analysis, cytoplasmic expression of COL-1 and OPN proteins was observed at 14 days. Enhanced cytoplasmic expression of both the proteins was observed at 21 days (Fig. 6).
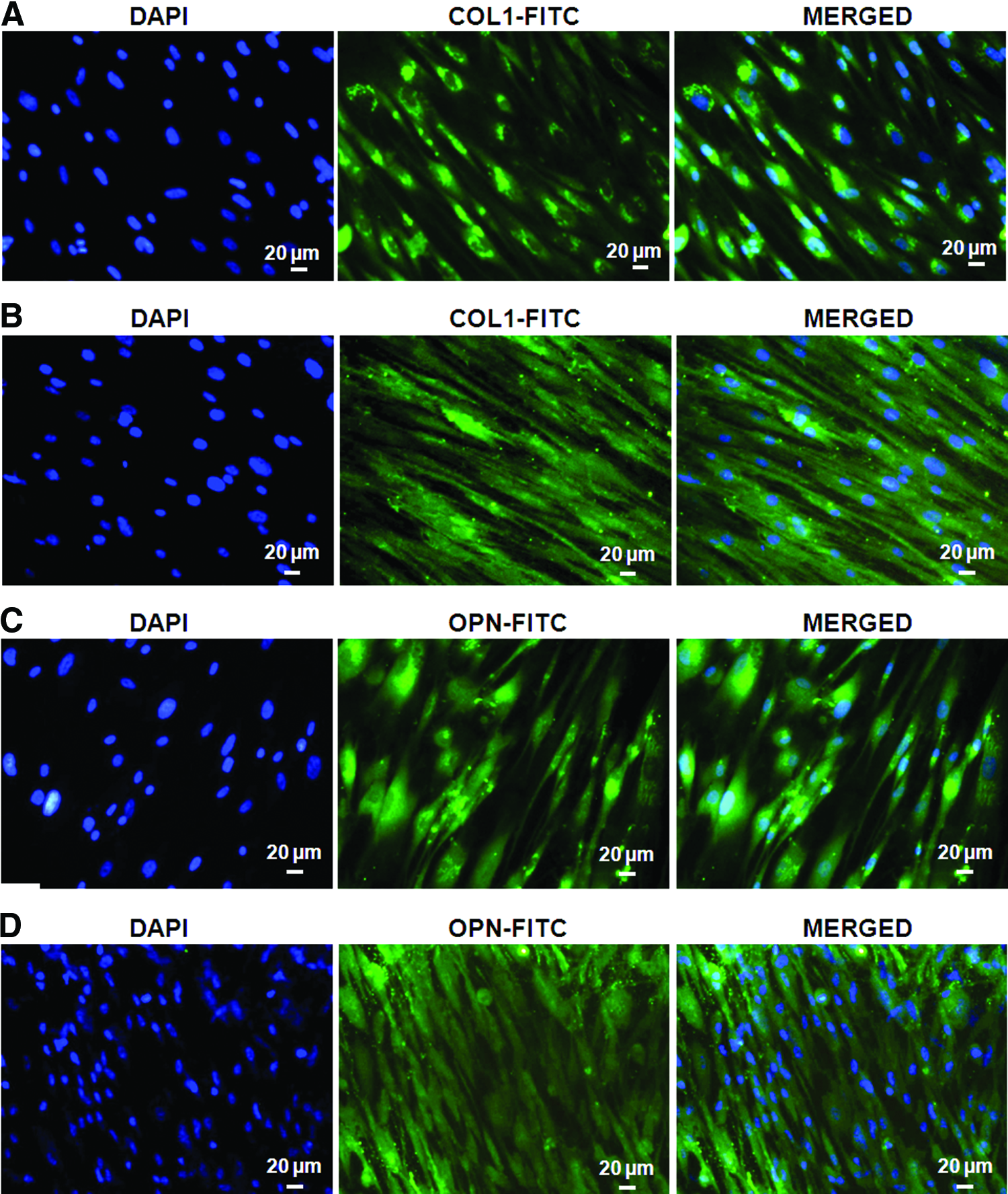
Immunocytochemical analysis of COL-1 at
Fibroblastic proliferation and differentiation
The effect of growth factor on the extent of DFC differentiation into PDLF was studied using fibroblast markers by immunocytochemistry, flow cytometry, and quantitative real-time PCR. DFCs cultured in FM showed a significantly higher DNA concentration at day 7 in comparison to day 1, 3, and 5 (Fig. 7A). Thereafter, no considerable increase in DNA concentration was observed.
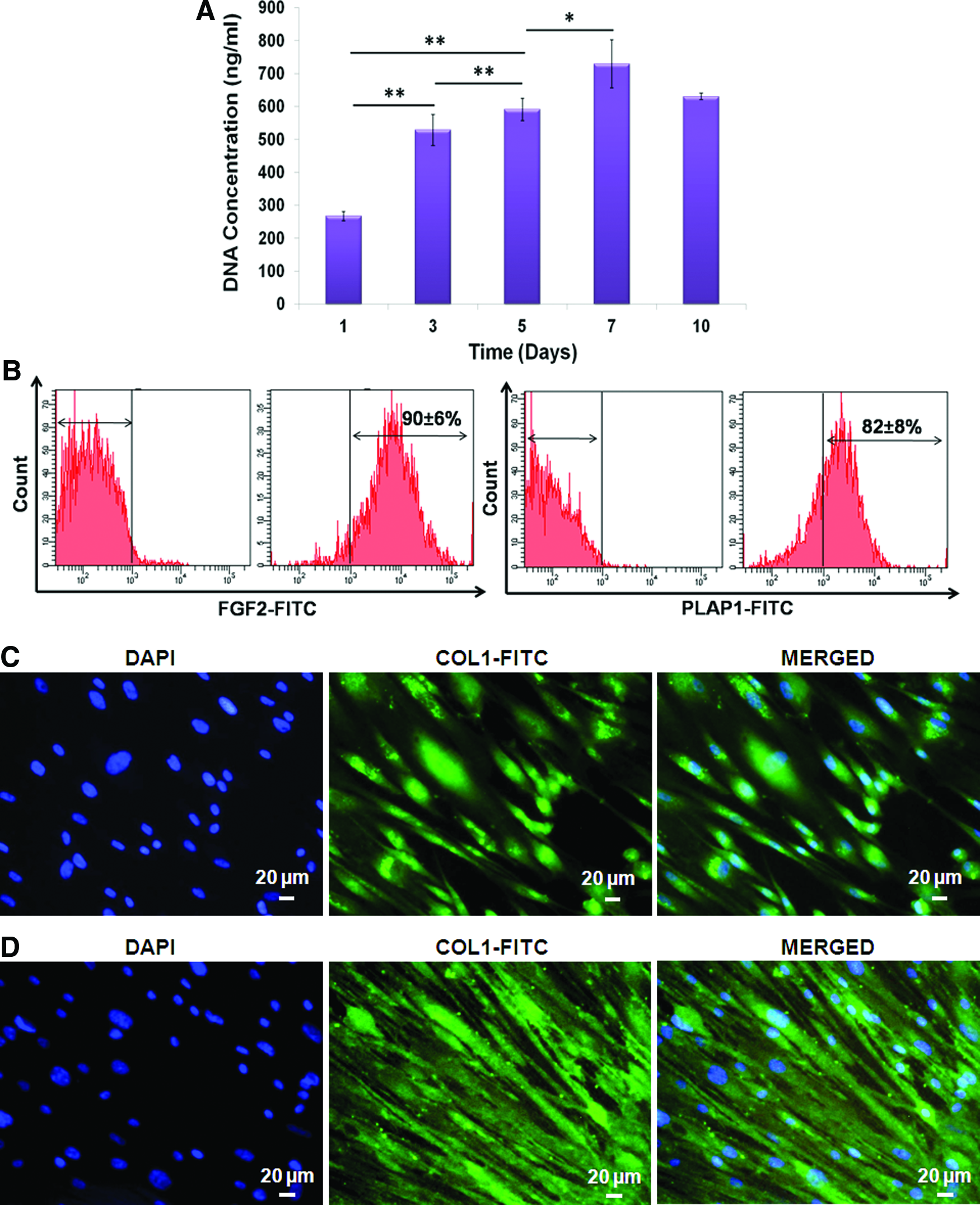
Expression of FGF-2, PLAP-1, and COL-1 proteins
DFCs were cultured in FM for 10 days and analyzed for the expression of FGF-2 and PLAP-1 proteins by flow cytometry. The FGF-2 and PLAP-1 protein expression was enhanced with increasing concentration of rhFGF-2 (Table 1). The highest expression of FGF-2 and PLAP-1 proteins was observed at 20 ng/mL concentration (Fig. 7B). In addition to FGF-2 and PLAP-1, the expression of COL-1 protein at 14 and 21 days was analyzed by immunocytochemistry. Cytoplasmic expression of COL-1 protein was observed at 14 days. The expression was further enhanced at 21 days (Fig. 7C, D).
Results are displayed as means of the protein expression in percent±standard deviation, n=3.
FGF-2, fibroblast growth factor-2; PLAP-1, periodontal ligament-associated protein-1; rhFGF-2, recombinant human FGF-2.
Cementogenic proliferation and differentiation
The effect of growth factor on the extent of DFC differentiation into cementoblast was studied using cementum-related markers by immunocytochemistry, flow cytometry, and quantitative real-time PCR. DFCs cultured in CM showed a significantly higher DNA concentration at day 7 in comparison to day 1, 3, and 5 (Fig. 8A). Thereafter, no significant increase in DNA concentration was observed.
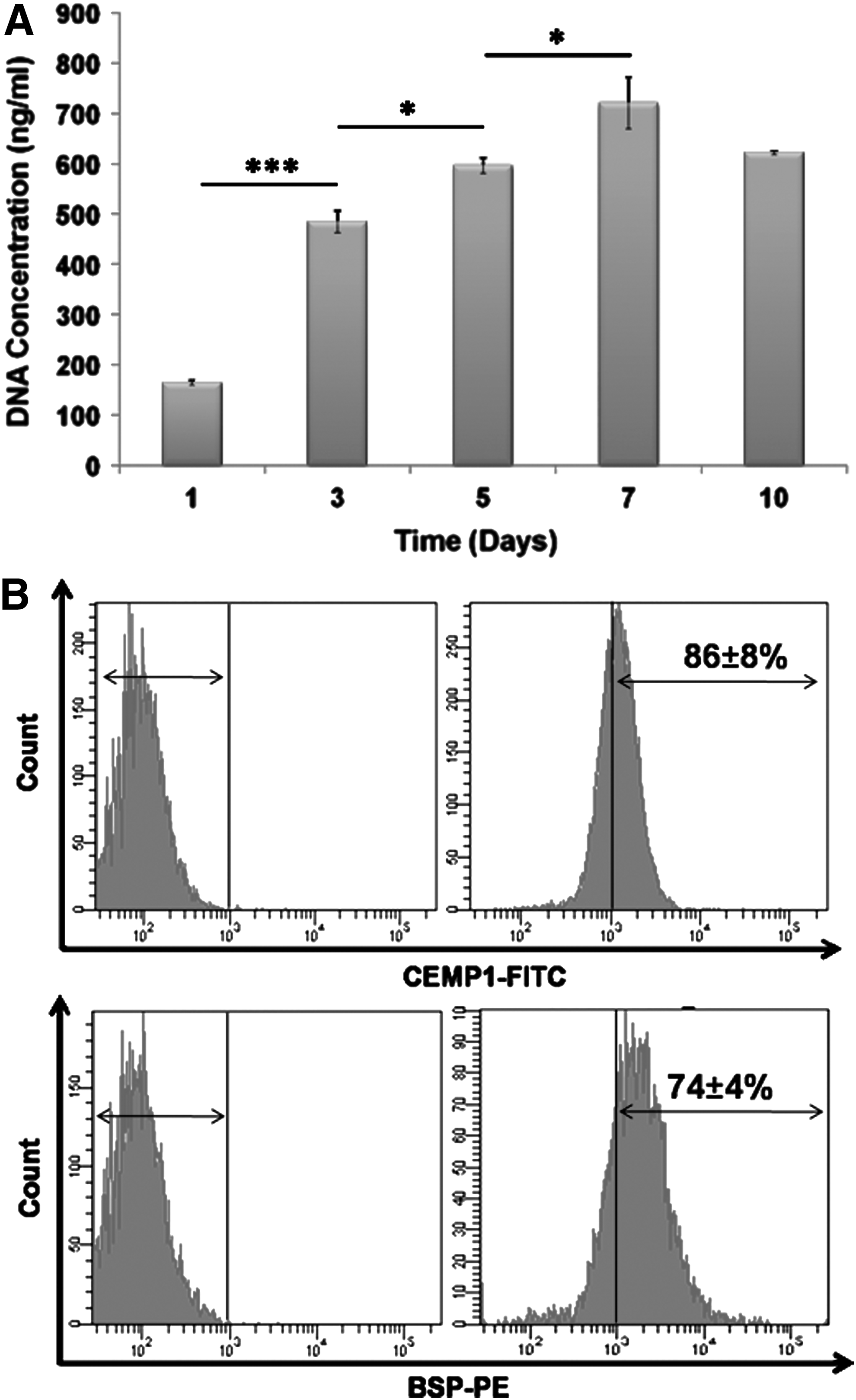
Expression of CEMP-1, BSP-2, COL-1, and OPN proteins
DFCs cultured in CM for 10 days were analyzed for the expression of CEMP-1 and BSP-2 by flow cytometry. The CEMP-1 and BSP-2 protein expression was enhanced with increasing concentration of rhCEMP-1 (Table 2). The highest expression of CEMP-1 and BSP-2 proteins was observed at 2 μg/ml concentration (Fig. 8B). Subsequently, COL-1 and OPN expression at 14 and 21 days was analyzed by immunocytochemistry. COL-1 and OPN protein expression was similar to osteogenic differentiation wherein cytoplasmic expression of both the proteins was observed at 14 days. The cytoplasmic expression of COL-1 and OPN was well enhanced at 21 days (Fig. 9).
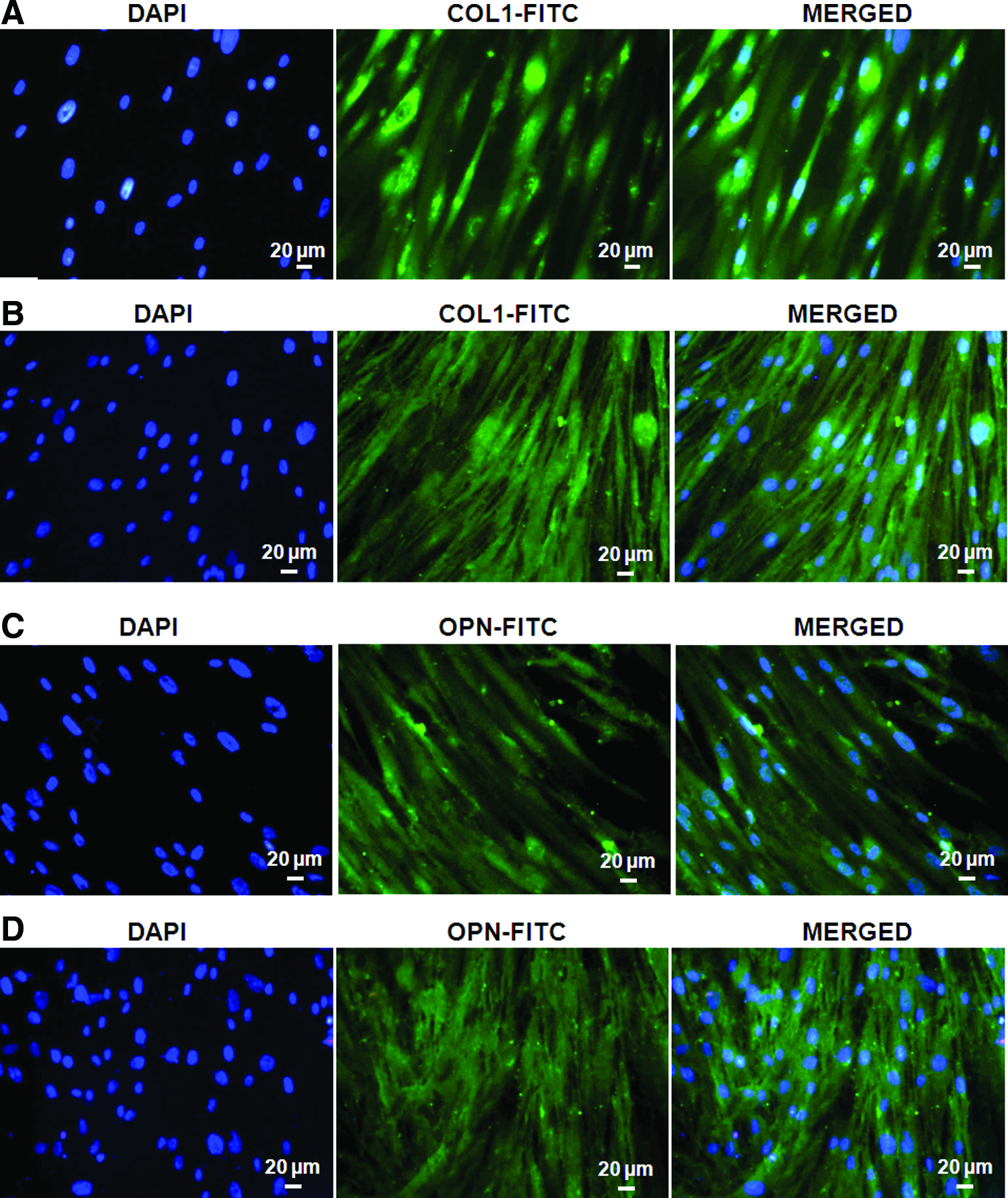
Immunocytochemical analysis of collagen type-1 (COL-1) at
Results are displayed as means of the protein expression in percent±standard deviation, n=3.
BSP-2, bone sialoprotein-2; CEMP-1, cementum protein-1; rhCEMP-1, recombinant human cementum protein-1.
Gene expression by quantitative real-time PCR
To validate the results on protein expression attained through immunocytochemical and flow cytometry analysis, quantitative real-time PCR was performed. The expression of ALP, OPN, OCN, and COL-1 genes in differentiating osteoblasts at 21 days was analyzed. Also, the expression of PLAP-1, FGFR-2, and COL-1 genes in differentiating fibroblasts and CEMP-1, ALP, OCN, OPN, and COL-1 genes in differentiating cementoblasts was analyzed. Results indicated that the expression of osteoblast, fibroblast, and cementoblast-related genes was significantly enhanced in DFCs cultured in OM, FM, and CM, compared to DFCs cultured in 10% DMEM (Fig. 10).

Quantitative real-time polymerase chain reaction analysis of
Discussion
The three major periodontal tissues namely cementum, periodontal ligament, and alveolar bone arise from the sac-like connective tissue of the developing tooth germ; the dental follicle. 25 Dental follicle is one among the major dental tissues that is reported to harbor mesenchymal stem cells. These DFCs possess the inherent and induced ability to differentiate into periodontal progenitor cell types namely cementoblast, osteoblast, and fibroblast, which led to the formation of cementum, alveolar bone and periodontal ligament-like tissues in vivo. 26 The existence of predifferentiated or heterogeneous periodontal progenitor population in the DFCs also supported the regeneration of periodontal tissues.19,27 Thus, to gain a detailed understanding of DFC differentiation into periodontal progenitors, first and foremost dental follicles were separated from extracted third molars and subjected to enzymatic digestion to isolate DFCs. Immunophenotypic characterization showed high expression of mesenchymal stem cell markers such as CD-73, CD-44, and CD-90. The expression of hematopoietic stem cell markers such as CD-33, CD-34, CD-45, and endothelial cell marker CD-31 was relatively low or absent. Thus, cell surface antigen analysis suggested that DFCs could also serve as a potential mesenchymal stem cell source similar to periodontal ligament and bone marrow-derived stem cells.
Immunolocalization of proteins, namely FGF-2 and CEMP-1, in periodontal ligament cells has already been reported.28,29 To determine the existence of a predifferentiated or progenitor population, the DFCs were analyzed for the expression of tissue-specific proteins namely PLAP-1 and FGF-2 for periodontal ligament, CEMP-1 for cementum, and RUNX-2 for alveolar bone. RUNX-2 expression was almost negligible. In addition, lack of ALP activity and mineralized deposits correspond to the absence of osteoblast progenitors. On the contrary, PLAP-1, FGF-2, and CEMP-1 expression was around 20–25%. Although the percentage of expression was limited, this expression could be indicative of the presence of fibroblastic and cementoblastic progenitor population in the dental follicle.
The ability of mesenchymal stem cells to secrete paracrine growth factors under serum-starved conditions is reported.23,24 In this study ELISA was performed to detect the secretion of paracrine factors from DFCs. The release of proteins such as VEGF, TGF-β1, FGF-2, and CEMP-1 as determined by ELISA confirmed that DFCs could also serve as a source for endogenous proteins. These cell-secreted factors/proteins would be beneficial in regulating wound healing, cellular proliferation, angiogenesis, osteoblast differentiation and osteogenesis, fibroblast proliferation and differentiation, and cementogenesis.23,24
Following the isolation and characterization of cells, it is essential to generate the three periodontal cell types namely osteoblast, fibroblast, and cementoblast, which would instigate the formation of alveolar bone, periodontal ligament, and cementum in vivo. To analyze osteoblast proliferation and differentiation, the DFCs were cultured in OM containing Dex, β-GP, and AA. During the course of osteoblast differentiation, a mesenchymal stem cell spans through different stages such as cellular proliferation, extracellular matrix (ECM) formation, matrix maturation, and mineralization; before transforming into an osteoblast. The influence of induction medium on DFC proliferation was analyzed by PicoGreen DNA quantification assay. An increase in DNA concentration corresponds to an increase in cell number. From the results, an evident increase in DNA concentration was observed upto 5 days. Thereafter, no significant increase was observed, which could be indicative of the termination of cellular proliferation and onset of cell differentiation induced by AA.30,31 Further to confirm, osteoblast differentiation was analyzed using specific markers such as RUNX-2, ALP, COL-1, and OPN along with alizarin staining and calcium quantification. Around 94% RUNX-2 expression confirmed the onset of osteoblast differentiation. RUNX-2 is a key transcription factor, which induces the differentiation of mesenchymal stem cells into immature or preosteoblasts, directing the formation of immature bone thereby inhibiting adipocyte and chondrocyte differentiation. 32 ALP is a biochemical marker of bone formation, synthesized and secreted by active osteoblasts. Its expression is prominently observed during the matrix maturation stage of osteoblast differentiation. 33 Under in vitro conditions, β-GP is regarded as the substrate for ALP, which is hydrolyzed to generate inorganic phosphate and thus encourages matrix mineralization in the course of osteoblastic differentiation. 34 The ALP activity of DFCs cultured in osteogenic medium peaked at seventh day followed by a decline thereafter. This decline in ALP activity might correspond to the onset of matrix mineralization. Alizarin staining revealed a time-dependent significant enhancement in calcium and phosphate deposition. Calcium deposition on the ECM was significantly higher in osteogenic culture in comparison to DFCs cultured in nonosteogenic medium, which thereby confirmed the onset and progression of matrix mineralization. In addition, the expression of COL-1 and OPN proteins, evaluated by immunocytochemical analysis, increased with an increase in culture period. COL-1, OCN, and OPN are the essential components of the organic matrix of bone. AA in the OM, plays a predominant role in the stabilization of cell-synthesized collagen matrix. 35 Among the various collagen types, type-1 is the major and abundant type of protein that forms the ECM of bone. It confers strength and structural integrity to the bone matrix and acts as a template for calcium phosphate mineral deposition. 36 Besides collagen, various noncollagenous proteins such as OPN, OCN, and BSP, are also synthesized and secreted. OPN is one such noncollagenous protein component of bone matrix, which is actively expressed during the proliferation and matrix mineralization stages, with its greatest level of expression during mineralization. 37 In our study, the results from immunocytochemical analysis also confirmed the enhanced expression of OPN protein with an increase in culture period, which could be indicative of matrix mineralization. In addition, the enhanced expression of osteoblast-related genes such as ALP, OPN, OCN, and COL-1 confirm the differentiation of DFCs into osteoblasts.
Periodontal ligament, a component of the periodontium, located between the cementum and alveolar bone is composed of collagen fibers, noncollagenous proteins, and a heterogeneous cell population. Fibroblasts are the principal cells of the PDL, which also possess the ability to differentiate into osteoblasts and cementoblasts, and can form the periodontal tissues namely cementum and alveolar bone in vivo. 38 Among the diverse range of growth factors, the potent role of FGF-2 in periodontal regeneration has been well established.39,40 In addition to its role in promoting angiogenesis, FGF-2 efficiently induces the proliferation and differentiation of PDL cells.41,42 From the immunostaining and flow cytometry results, the existence and distribution of FGF-2 in the DFCs was confirmed. Based on previous reports, 28 the FGF-2 expressing DFCs were regarded as PDLF (which were negative for CD-31 expression). Hence, DFCs were stimulated with rhFGF-2 and AA and analyzed for proliferation and differentiation. From the proliferation results, an evident increase in DNA concentration was observed up to day 7. No further increase in DNA concentration might correspond to the onset of cell differentiation induced by AA. 31 PLAP-1, a PDL specific marker, is believed to inhibit the differentiation of PDLF into hard tissue forming cells namely osteoblasts and cementoblasts and hence localized to the PDL region.43,44 The enhanced expression of PLAP-1 and FGF-2 proteins with respect to increasing concentration of rhFGF-2 might correspond to PDLF proliferation and differentiation. Collagen, the chief component of periodontal ligament is secreted by fibroblasts. Interestingly, FGF-2 is reported to downregulate the expression of COL-1, which may perhaps decrease the synthesis or secretion of collagen matrix.41,45 Hence, to overcome the effects of FGF-2 on collagen synthesis, DFCs were stimulated with AA in the presence of rhFGF-2 to facilitate collagen secretion and its stabilization. Immunocytochemical analysis confirmed the expression of COL-1 protein, which was well enhanced with the increase in culture time. In addition, the enhanced expression of PLAP-1, FGFR-2, and COL-1 genes also indicates DFCs differentiation into PDLF.
CEMP-1, a cementum-specific protein is believed to play a key role in cementoblastic differentiation of DFCs and PDL cells.46,47 As indicated earlier, immunostaining and flow cytometry analysis of DFCs confirmed the expression of CEMP-1. Thus, to facilitate cementoblast differentiation, DFCs were cultured in CM and analyzed for the expression of cementum makers. COL-1 is the major component of the organic matrix of cementum, similar to bone. It confers structural and functional support to the cell secreted calcium-phosphate mineral crystals. 48 BSP and OPN, the noncollagenous proteins of the organic matrix, are confined to the acellular extrinsic fiber and acellular afibrillar cementum. They are believed to assist cellular adhesion, migration, cementoblastic differentiation, and matrix mineralization similar to osteoblasts.48,49 They play a predominant role in the initiation and progression of osteoblast/cementoblast mineralization.49,50 BSP acts as a crystal nucleator, whereas OPN inhibits the precipitation of redundant calcium-phosphate crystals and thereby ensures precise crystal formation. 50 Based on these reports, the expression of ECM proteins namely COL-I, BSP, and OPN was analyzed. The enhanced gene and protein expression with respect to rhCEMP-1 concentration and culture time correspond to cementoblastic proliferation and differentiation.
Conclusions
To determine the effect of growth factor concentration on the differentiation of DFCs, first and foremost DFCs were isolated from dental follicle tissue and characterized. These DFCs of mesenchymal origin also expressed periodontal ligament and cementum markers such as PLAP-1, FGF-2, and CEMP-1, which confirmed the existence of progenitor cell population in the dental follicle. The DFCs cultured in induction medium showed good proliferation ability followed by differentiation. Growth factor concentration-dependent fibroblastic and cementoblastic differentiation of DFCs was observed. These findings would be beneficial for cell-based tissue engineering approaches wherein predifferentiated osteoblasts, fibroblasts, and cementoblasts would enhance the rate of periodontal regeneration.
Footnotes
Acknowledgments
The authors are grateful to Nanomission, Department of Science and Technology (DST), Government of India, which supported this work under the “Thematic Unit of Excellence.” The authors are also grateful to SERB Division, Department of Science and Technology (DST), Government of India for providing research funding support (File No. SR/S1/OC-19/2012). The author, S. Sowmya acknowledges the Council of Scientific and Industrial Research (CSIR) for the financial support through Senior Research Fellowship [SRF-Award No. 9/963(0022)2K12-EMR-I]. The authors are thankful to Dr. P. Mathew (Department of Oral and Maxillofacial surgery, Amrita School of Dentistry, Amrita Institute of Medical Sciences and Research Centre) for providing human dental follicle tissues. The author S. Sowmya is grateful to Dr. A. Sampath Narayanan, Department of Pathology, School of Medicine, University of Washington, Seattle, for his immense help and support in providing rhCEMP-1 for the cementogenic differentiation studies.
Disclosure Statement
No competing financial interests exist.