
Abstract
Whole heart decellularization combined with patient-specific cells may prove to be an extremely valuable approach to engineer new hearts. Mild detergents are commonly used in the decellularization process, but are known to denature and solubilize key proteins and growth factors and can therefore be destructive to the extracellular matrix (ECM) during the decellularization process. In this study, the decellularization of porcine hearts was accomplished in 24 h with only 6 h of sodium dodecyl sulfate exposure and 98% DNA removal. Automatically controlling the pressure during decellularization reduced the detergent exposure time while still completely removing immunogenic cell debris. Stimulation of macrophages was greatly reduced when comparing native tissue samples to the processed ECM. Complete cell removal was confirmed by analysis of DNA content. General collagen and elastin preservation was demonstrated. Glycosaminoglycans and collagen quantification both showed no significant differences in content after decellularization. The compression elastic modulus of the ECM after decellularization was lower than native at low strains, but there was no significant difference at high strains. Polyurethane casts of the vasculature of native and decellularized hearts demonstrated that the microvasculature network was preserved after decellularization. A static blood thrombosis assay using bovine blood was also developed. Finally, the recellularization potential of the ECM samples was demonstrated by reseeding cardiac fibroblasts and endothelial cells on the myocardium and endocardium samples.
Introduction
H
Advances in technology, including total artificial heart transplantations, ventricular assist devices,3,4 and improved treatment techniques, 5 have provided some improvement in the treatment of heart disease and slowed the accelerating need for end-stage organ transplantation, yet alone are insufficient to meet the rising demand. The latest technologies, including stem cell patches, 6 three-dimensional (3D) printing of tissues and whole organs, 7 and tissue engineering using extracellular matrix (ECM)-derived biologic scaffolds, are currently under development.
The ECM consists of thousands of types of proteins, the concentration of each varying between organs, and the mechanical properties change based on the environment and age. 8 In addition to providing a mechanical scaffold for homing and 3D functioning of the cells, the ECM serves as a signaling and communication material between the cells and promotes cell proliferation and differentiation in the body. 9
Synthetic polymer scaffolds lack in advancing complexity of vasculature and essential growth and differentiation factors. 7 The use of decellularized tissue scaffolds has become an attractive solution as heart vasculature, mechanical integrity, and biocompatibility are preserved, while the magnitude of the immunogenic response after transplantation is reduced if seeded with terminally differentiated autologous induced pluripotent stem cells (iPSCs).10,11
The ECM provides an acellular structure capable of being seeded by iPSCs or progenitor cells. 12 However, organ structural complexity raises issues of oxygenation and nourishing of seeded cells from the difficulty of effective diffusion through the entire vasculature. 13 Less complicated organs, such as the trachea 14 or urinary bladder, 15 have less such problems and are already capable of recellularization. The heart provides a unique opportunity because retrograde perfusion through the aorta can facilitate vascular perfusion. 16
Ideally, decellularization is a process in which the cellular components that promote a chronic immune response, such as DNA and GAL epitopes, are removed from the ECM, while the ECM nonimmunogenic matricellular proteins and growth factors are preserved. 9 A key challenge in decellularization is to avoid chemical damage from overexposure of cell removal reagents such as sodium dodecyl sulfate (SDS). Although SDS is considered one of the most effective detergents for removing cells and immunogenic components from the ECM, 17 excess exposure creates a risk of removing glycosaminoglycans (GAGs) and growth factors, as well as denaturing ligands and proteins such as collagen, elastin, and fibronectin within the structure.18,19
Automated systems assist in standardizing the decellularization process 20 and a pressure controlled system can assure effective perfusion through the coronary vessels during the decellularization process. 21 In this study, automated pressure control was used to optimize the decellularization protocol with retrograde perfusion through the aorta in a porcine heart model. Once hearts were decellularized into cardiac ECM (cECM), they were characterized chemically, mechanically, and the in vitro thrombosis potential was investigated. The cECM was partially recellularized using endothelial cells and human cardiac fibroblasts (HCF) to demonstrate the recellularization potential.
Materials and Methods
Harvesting
Porcine hearts were harvested from slaughter-weight, 6-month-old, mature female Hampshire, Yorkshire, or Spot crossbreed swine at a local abattoir according to approved protocols for safety and animal care. After exsanguination, the chest cavity was opened using a 12″ Finochietto retractor and the heart was dissected from the lung while preserving as much of the aorta, pulmonary artery, vein, and caudal and cranial vena cava as possible.
Immediately after removing the heart, a pressurized hand action pump filled with a solution of 1× phosphate-buffered saline (PBS), heparin (10 U/mL), and 1× antibiotic solution (100 U/mL penicillin, 100 μg/mL streptomycin, 25 μg/mL amphotericin B, Anti-Anti; Gibco, Billings, MT) was used to rinse all the chambers and vessels of the heart by means of antegrade perfusion through the vena cava and pulmonary vein and retrograde perfusion through the aorta. Hearts were manually massaged while being washed to remove any static blood in the coronary vessels to prevent further thrombosis. The hearts were then transported in cold heparinized (10 U/mL) type 1 distilled water to the laboratory and frozen at −20°C.
Decellularization apparatus design and automation
A Langendorff system based on the Radnoti® porcine working heart system (Monrovia, CA) was modified to be used as the decellularization apparatus. The modified apparatus contained a 5-L reservoir for the perfusate solution, a heat exchanger, a bubble trap, and the main 5-L decellularization chamber, with the heart submerged in the perfusate solution. A peristaltic pump (Masterflex® HV-07557-00, Vernon Hills, IL) with Tygon L/S 36 tubing (Masterflex® E-3603) was used to produce a pressurized pulsatile flow.
A negative feedback control system was implemented to maintain critical parameters during the decellularization process. The system consisted of a computer running National Instruments (Austin, TX) LabVIEW software and contained hardware to collect pressure data while sending control signals to a pump. A proportional-integral-derivative control algorithm monitored the pressure signal every tenth of a second and adjusted the pump signal to maintain the desired pressure.
The program also included functions for running timed experiments, automatically adjusting pressure levels, and switching fluid streams through electrically controlled solenoid valves, as well as a manual control mode (Supplementary Figs. S1–S3; Supplementary Data are available online at
A 10-L water bath with controlled temperature was used to maintain the temperature of the decellularization process at a desired temperature (23°C) by circulating the water through the jackets of the reservoir and the main chamber, as well as the heat exchanger and the bubble trap. Figure 1 shows the schematic of the apparatus design and the interface of the LabVIEW program. The operating apparatus is shown in Supplementary Videos S1 and S2.

Automated controller interface (left) and schematic view of the automated pressure controlled bioreactor for porcine heart decellularization (right). 1, solution reservoir; 2, peristaltic pump; 3, heat exchanger; 4, bubble trap; 5, decellularization chamber; 6, temperature controlled water bath; 7, control loop.
Decellularization
Decellularization of porcine hearts was performed using SDS and Triton X-100 in the decellularization apparatus. First, the hearts were thawed at 4°C overnight and removed from the 10 U/mL heparinized water. Excess fat was trimmed and the aorta was separated from the pulmonary artery and cut perpendicularly at the brachiocephalic trunk. The aorta was then cannulated with a Radnoti ½″ male tube and connected to the apparatus. The heart was submerged in the 1× PBS solution to avoid axial stress on the aorta due to the heart's weight. Care was taken while connecting the heart to attach the connector away from the aortic valve to ensure that the coronary orifices were not covered and the valve could work properly.
The decellularization process then started with perfusing 1× PBS through the coronary vessels to remove the blood and thromboemboli followed by an hour of nonrecycled perfusion of type 1 distilled water. Alternating between PBS and distilled water resulted in a hypotonic shock to the cells, facilitating decellularization.
During the 1 h of PBS perfusion, the pressure was gradually increased to 4 psi using the automated system and distilled water perfusion was performed at 5 psi. The perfusion continued with three recirculating 10 L batches of 0.5% (w/v) SDS in distilled water for 2 h, followed by 2 h of nonrecirculating distilled water, all performed at controlled pressure of 5 psi. The sixth hour of SDS exposure was followed by 2 h of nonrecirculating distilled water at 5 psi and 12 h of overnight perfusion of 10 L recirculating distilled water at 3 psi. The water was then replaced with 10 L 1% (v/v) Triton X-100 and perfusion was conducted while automatically increasing pressure to 5 psi for 2 h to remove remaining SDS and cellular debris.
Decellularization was finished with an extensive wash of the heart with distilled water at 5 psi for at least 5 h. The decellularized heart was then dissected and random samples from the left and right atria, ventricles, and septum wall were selected for characterization. Samples from the left ventricle were isolated for recellularization or the heart was preserved intact for corrosion casting.
DNA, collagen, and GAG measurements
To measure the amount of residual DNA fragments, samples were taken from left and right ventricles, left and right atria, and the septa of decellularized and native hearts. Samples were sterilized by immersion in 1× antibiotic solution in 70% ethyl alcohol for 2 h, and then the decellularized samples were submerged in a cell growth medium (DMEM/F-12 with 10% fetal bovine serum [FBS; Gibco], 100 U/mL penicillin, and 100 μg/mL streptomycin [Gibco]) for 48 h. The samples were then lyophilized and weighed. DNA was extracted from ∼5 mg of dry samples using Qiagen® DNeasy Blood and Tissue Kit (Valencia, CA).
The samples were solubilized with proteinase K overnight and after several centrifugation steps and washing with different buffers, DNA extracts were prepared for quantification. The Quant-iT PicoGreen (Invitrogen Corp., Carlsbad, CA) reagent was added to each sample. Samples were excited at 485 nm, and fluorescence intensity was measured at 520 nm using a Synergy 2® Multi-Mode Microplate Reader. Gel electrophoresis was performed with DNA extracts loaded on 1% agarose gels to characterize the residual DNA in the structure of the ECM.
To examine the collagen and sulfated glycosaminoglycan (sGAG) content of the cECM, the Sircol® Soluble Collagen Assay Kit (Biocolor Ltd., Newtownabbey, United Kingdom) and the Blyscan® sGAG Assay Kit (Biocolor, Ltd.) were used, respectively. Approximately 3 mg of dry cECM samples was digested over 3 days using an acid/pepsin solution at 4°C and the Sircol® Dye Reagent was added to each sample to form a collagen–dye complex based on the manufacturer's instructions.
After solubilizing the dye–reagent complex with an alkali reagent, the absorbance of samples at 555 nm was measured. For sGAG measurements, each sample was completely solubilized in papain extraction solution for 3 h at 65°C and the Blyscan® dye reagent was added to all samples to precipitate the sGAG–dye complex. The dissociation reagent was then added to dissolve the complex and absorbance was measured at 659 nm.
Cell culture
Murine endothelial cells (MS-1 cell line) labeled with the green fluorescent protein (GFP) were thawed and cultured in the cell growth medium (DMEM/F-12 with 10% FBS [Gibco], 100 U/mL penicillin, and 100 μg/mL streptomycin [Gibco]). HCF cells were cultured in the Cardiac Fibroblast Growth Medium (Cell Applications, Inc., San Diego, CA). MS-1 and HCF cultures were maintained at 37°C and 5% CO2 and both were used at early passages. A FLoid® Cell Imaging Station was used to visualize and measure the confluency of the cells as well as for image acquisition.
Macrophage stimulation assay
The immunogenic potential of the cECM was measured and compared to fresh heart samples by quantifying the amount of nitric oxide (NO) generated by macrophage cells in contact with specimens. Mouse leukemic monocytes and macrophages (cell line RAW 264.7, Sigma-Aldrich, St. Louis, MO) were cultured with 3 mL of cell growth medium in six-well plates until 70% confluent. Lyophilized samples of decellularized (n=6) and native (n=6) hearts were ground to a fine powder using a pestle and mortar filled with liquid nitrogen. Fourteen milligrams of each specimen was added to a well of six-well plates.
To ensure the production of NO by stimulated macrophages, powdered lyophilized Escherichia coli bacterial extract was added to wells as positive controls and wells without extract served as negative controls. After 24 h of incubation at 37°C, 5% CO2, all media were centrifuged at 15,000 g and the amount of NO in the supernatant was quantified as the free stable nitrite form (NO2−) using the Griess Reagent Kit (Life Technology, Grand Island, NY) according to the manufacturer's instructions. The concentration of nitrite in the media was then interpolated from a nitrite standard reference curve (0 to 100 μM).
Corrosion casting
Corrosion casting was performed on native and decellularized porcine hearts to ensure the integrity of the coronary vessels after decellularization. Two hearts were prepared for casting by surgically suturing the aortic valve closed and placing two 2.5-cm-diameter (1″) and 1.6-mm-thick (1/16″) N45 neodymium magnets on either side of the aortic valve. Magnets were positioned carefully not to block the coronary orifices. The aortas were then cannulated with a Radnoti® ½″ male tube and connected to a peristaltic pump. A solution of ice-cold 2× PBS dyed with trypan blue was slowly pumped into the heart to ensure perfusion through the coronary vasculature and to hypertonically shrink the native endothelial cells to enhance perfusion.
Ice-cold polyurethane and an epoxy casting resin containing a catalyst (Dowagiac®; Specialty Resin & Chemical LLC, Dowagiac, MI) were then mixed at a 35:65 ratio, and immediately perfused through the heart for 5 min. The tubes were then detached and the hearts were allowed to be incubated at room temperature for 1 h to ensure curing was complete. Hearts were then immersed in 3.5 L of 15% NaOH at 37°C to digest the organic tissue. The NaOH solutions were renewed every 3 days or when the solutions became cloudy due to being saturated with the digested tissues. After 10 days, the heart tissues were fully digested and the polymer casts of the internal coronary vasculature were visible. The hearts were placed in 1× PBS overnight, and then the residual tissue was manually removed.
Histology and scanning electron microscopy
Decellularized, native, and re-endothelialized cardiac samples from left and right ventricles and atria and the septum wall were dissected and fixed overnight in a solution of 4% paraformaldehyde (PFA; Sigma- Aldrich) in 1× PBS. The samples were then dehydrated using 30%, 50%, 70%, 95%, and 100% ethanol. Afterward, samples were immersed in xylene and embedded in paraffin. Using a microtome, 5 μm slices were made and placed on Fisherbrand® (Pittsburgh, PA) microscope slides.
Sectioned samples were then deparaffinized and stained according to the manufacturer's instructions with hematoxylin and eosin (H&E; Thermo Scientific, Pittsburgh, PA), Orcein (O7380; Sigma), or Picrosirius red (Direct Red 80, #365548; Sigma-Aldrich) to examine the cellular components, elastin, and general collagen in the native and decellularized samples. Images were acquired using an EVOS® FL Cell Imaging System (Life Technology, Grand Island, NY).
Scanning electron microscopy (SEM) was performed on decellularized and native heart samples for surface characterization. Specimens were prepared for high-vacuum microscopy by fixation in 2% glutaraldehyde in 1× PBS for 24 h at 4°C, dehydration in acetone, and crosslinked with OsO4. Samples were then dried in a CO2 critical drier and coated with a 3 nm layer of platinum to enhance conductivity. A Philips®/FEI XL30 ESEM™ FEG (Hillsboro, OR) Environmental Scanning Electron Microscope was used to acquire images with a field emission gun (FEG) electronic beam at high vacuum.
Mechanical properties assay
Elastic moduli of the decellularized and native heart samples were calculated by measuring the compression stress as a function of strain to evaluate the mechanical stiffness of the decellularized heart samples compared to native hearts kept in PBS at 4°C for 24 h after harvesting. Square-shaped, wet samples, ∼10×10 mm and 5 mm thick, were longitudinally dissected from the septum and left and right atrium and ventricle walls.
An Instron® 3342 Single Column Universal Testing System (Instron, Norwood, MA) was used to perform compression tests. Tissue samples at room temperature were placed flat on the lower metal plate and compressed at a constant rate of 0.07 mm/s until a compressive force of 45N was achieved. Plots of stress versus strain for the specimens were created and the elastic moduli were calculated at the linear portions of the stress/strain curves.
Cytotoxicity assay
MS-1 endothelial cells were used to assess cytotoxicity of the decellularized hearts to ensure that the cECM did not adversely affect cell viability. One-cm2 samples were dissected randomly from the endocardium of the left ventricular walls of decellularized porcine hearts and sterilized by immersion in a solution of 1× antibiotic solution in 70% ethyl alcohol overnight. Samples were then soaked twice in 1× PBS and incubated in the cell growth medium at 37°C, 5% CO2 for 48 h. Samples of cECM (n=9) in 3 mL of cell growth medium were then placed in wells of six-well plates, while wells without cECM samples served as controls.
MS-1 endothelial cells (1×105) were detached from culture flasks using 0.05% Trypsin-EDTA and seeded onto each of the cECM samples and control wells. Cells were labeled with 2 μM calcein AM and 4 μM EthD-III (Biotium® Viability/Cytotoxicity Assay, Hayward, CA) to examine the health and viability of the cells in samples with and without cECM after 1 day and 3 days of culture.
Recellularization
To recellularize cECM samples, MS-1 and HCF cells were labeled with Vybrant® DiO and DiI lipophilic tracers (Life Technologies, Grand Island, NY), respectively. Labeling was performed by incubating 70% confluent cell layers in T-75 culture flasks with 5 mL of serum-free media and 25 μL of the supplied dye labeling solution at 37°C for 20 min. Cells were then detached using Trypsin/EDTA, centrifuged, and slowly dispensed, at a concentration of 104 cells/mL, onto 8-mm-diameter round, 1-mm-thick cECM samples (n=12) placed in 48-well plates. One milliliter of medium was renewed every 2 days. Representative fluorescence images were acquired after 1 week of culture and recellularized samples were fixed in 4% PFA for histology imaging.
Thrombosis assay
A static blood thrombosis assay was developed to qualitatively examine the thrombogenicity of porcine hearts in vitro. Bovine blood was collected at a local abattoir by cannulating the aorta of the heart immediately after slaughter using an 18″-long, 1″-diameter cannula and collecting the blood in 1-L flasks containing 1500 U of heparin as described by Zhang et al. 22 Bovine blood was used with decellularized bovine and porcine hearts to develop the assay.
Bovine hearts were decellularized in 7 days with immersion and aortic perfusion of 0.5% SDS, rinsing extensively with type 1 distilled water. Freshly obtained blood was then perfused through coronary arteries until the chambers of the heart were filled; the heart vessels were then clamped and the hearts were immersed in a container of blood for 4 h. Native bovine and porcine hearts were used as controls and similarly perfused. After 4 h the hearts were thoroughly rinsed with 1× PBS through all chambers and the aorta. To evaluate thrombosis, blood clots that could be rinsed out of the heart chambers and vessels were examined visually and the hearts were dissected open to assess platelet adherence to the chamber walls.
Statistical analysis
Data are shown as average±standard deviation and the sample size for all of the quantification experiments was n=5, unless stated otherwise. SAS JMP® 11 (Cary, NC) software was used for statistical analysis of the data. For examining the significance of comparisons, α=0.05 was considered as the significance level of p-values. Student's t-test was used for analyzing the data in two groups and the Tukey–Kramer test was performed on the data with multiple groups.
Because of the change in density of the heart after decellularization, the data for quantities of sGAGs and collagen in cECM samples were normalized and reported using the dry mass of the native hearts as the base. For the normalization, the overall weight loss of each of the compartments (right and left atria, ventricular walls, and septum wall) was determined by measuring the ratio of water content to the mass of lyophilized samples in both cECM and native groups and calculating their fraction. The standard deviations of the reported quantities of sGAGs and collagen in cECM samples were determined considering the standard deviations of initial non-normalized values and decellularization weight loss of dried samples.
Results
Whole heart decellularization
A total of 20 porcine hearts were used in these studies; 10 native hearts and 10 hearts were decellularized using the controlled pressure system (Fig. 1) with 6 h of ionic detergent wash (SDS) and 2 h of nonionic detergent (TX-100). Isotonic and hypotonic shocks with PBS and type 1 distilled water, respectively, before and in between steps of the decellularization process, improved the quality of the decellularization and decreased the total decellularization time and detergent exposure.
It was observed that the required flow rate to achieve the set point pressure during the SDS perfusion increased from 850 mL/min to 2250 mL/min, which is due to cell removal and expansion of the coronary vessels. Figure 2 shows a representative porcine heart before (Fig. 2A) and 24 h after decellularization (Fig. 2B). The remaining ECM was white or translucent due to the disappearance of the red coloration caused by blood and myoglobin proteins. The heart's shape was preserved and the heart valves remained intact and functional.

A representative porcine heart before
At the time of harvesting, most of the hearts were soft and flexible. At the beginning of decellularization, when they were transferred from the refrigerator to the apparatus, the hearts were rigid and inflexible, which helped the decellularization process by keeping the aortic valve closed, directing the perfusion through coronary artery orifices. After decellularization was finished, the cECM became soft and flexible again.
In the H&E staining (Fig. 2C, D), the cell cytoplasm was stained with eosin Y (pink color) and the cell nuclei stained with hematoxylin (blue color). The decellularization process removed the cells and cellular components. Sirius general collagen stain (Fig. 2E, F) stained collagen protein fibers red, cell cytoplasm yellow, and nuclei black. Comparing the representative images from these different sections of the heart before and after decellularization, the process removed cells and cellular components while the collagen matrix was kept intact. Elastin protein fibers were visualized using the Orcein stain represented by the dark brown color as shown in Figure 2G and H.
Deoxyribonucleic acid
DNA can initiate an immune response if it enters a foreign body. However, in addition to DNA, GAL epitope and other membrane proteins and epitopes have been shown to cause an acute immune response that can lead to rejection of a transplanted organ. 23 Since DNA is in the nuclei of the cells, it can be used as a marker for cell removal and decellularization efficacy. 24 Samples of cECM (n=5) from five different sections (right and left atria, ventricular walls, and septum wall) of three decellularized hearts were prepared for recellularization by sterilization and incubation in cell growth media, which contain DNAses that are known to digest residual DNA. 25
The DNA content was extracted using Qiagen™ DNeasy® kit, fluorescent-stained with Quant-iT™ PicoGreen®, and the fluorescence was measured. As shown in Figure 3A, the amounts of residual DNA varied in different portions of the heart after decellularization. Samples from the right atrium and left atrium exhibited the highest and lowest DNA removal values (with averages of 99% and 97%, respectively) from five different portions of the heart. p-Values of less than 0.0001 suggest strong statistical evidence for DNA removal from all portions of the heart after automated, pressure controlled decellularization.
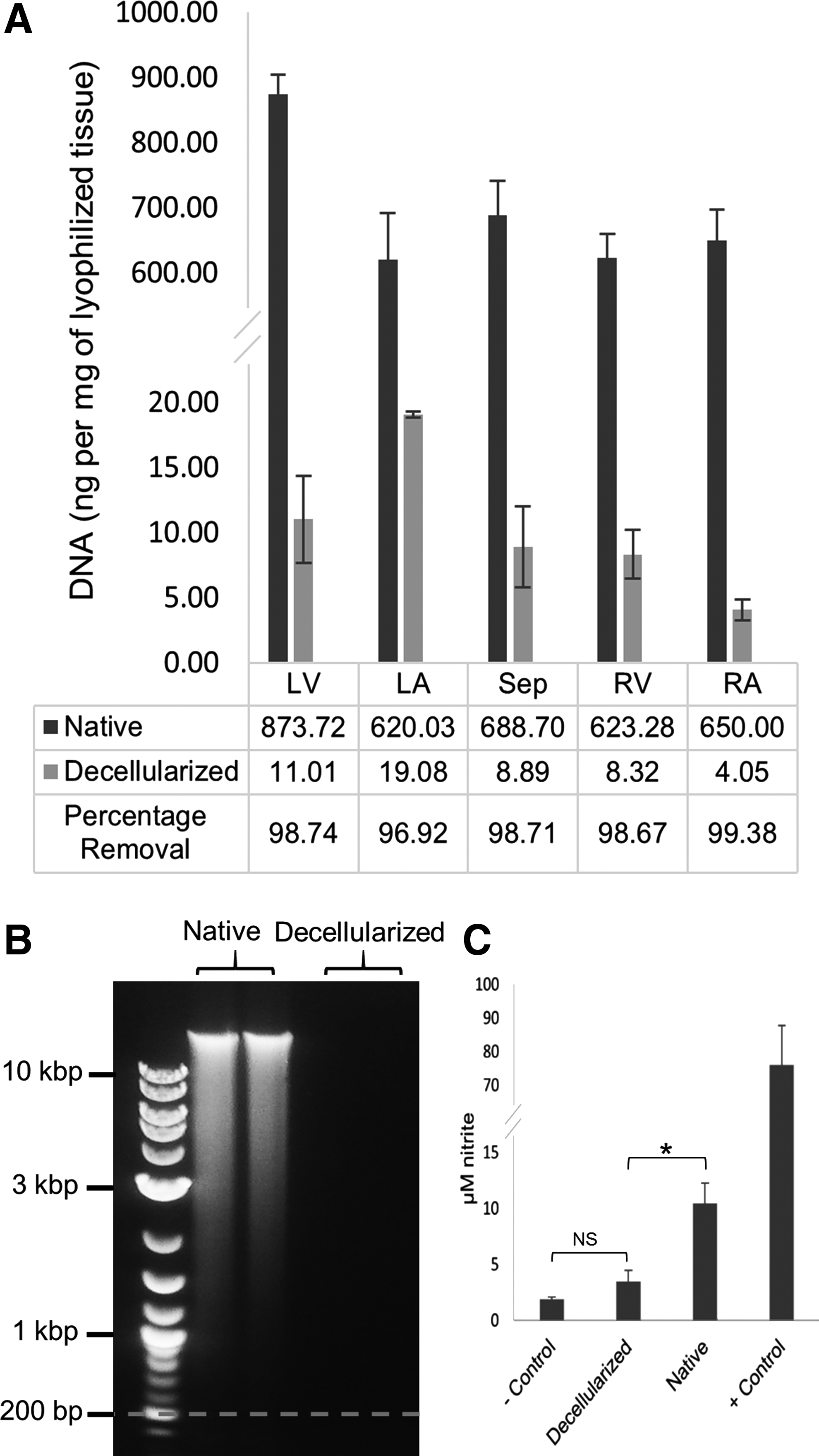
The number of DNA base pairs (bp) in left ventricle samples of native and decellularized hearts was determined by means of gel electrophoresis. It has been suggested that DNA larger than 200 bp remaining in the decellularized ECM can cause a chronic immune response. 26 As shown in Figure 3B, no DNA with more than 200 bp length in cECM samples was detected, while a wide range of DNA sizes were observed in the native heart samples.
Macrophage stimulation
Macrophages are white blood cells that can contribute to stimulating the immune system by encouraging inflammation (M1 phenotype, killer) or moderating the immune response and promoting tissue repair (M2 phenotype, healer). Killer macrophages secrete NO as they stimulate the immune system.27,28 The level of stimulation can be determined by measuring the concentration of produced NO. 29
The amount of NO produced by stimulated macrophage cells was quantified as stable nitrite in the medium using the Griess Reagent® Kit for decellularized (n=6) and native (n=6) left ventricular samples in the presence of positive and negative controls. As shown in Figure 3C, the concentration of nitrite (μM) formed in the media by macrophage cells exposed to either the negative control or stimulated with decellularized specimens did not produce statistically significant differences (p-value=0.80), while the native and decellularized groups had a 7.27 μM difference in their average of secreted nitrite with high statistical significance (p-value <0.0001). The positive controls showed the highest capacity of nitrite production by the macrophage cells (80.35 μM).
Glycosaminoglycan
To characterize cECM and examine the effects of automated, pressure controlled decellularization on the GAG content of the hearts, five random biopsies were obtained from different sections (right and left atria, ventricular walls, and septum wall) of three decellularized hearts. sGAGs were quantified using Blyscan® sGAGs Assay and the normalized results are shown in Figure 4A. Comparing the average amounts of sGAGs between the native and decellularized samples, it was observed that the amounts of sGAGs were not significantly different in various sections of the native heart samples except in the right atrium, which had the highest quantity of sGAGs; however, a statistically significant difference was observed between all groups of cECM samples except the right atrium, right ventricle, and septum.
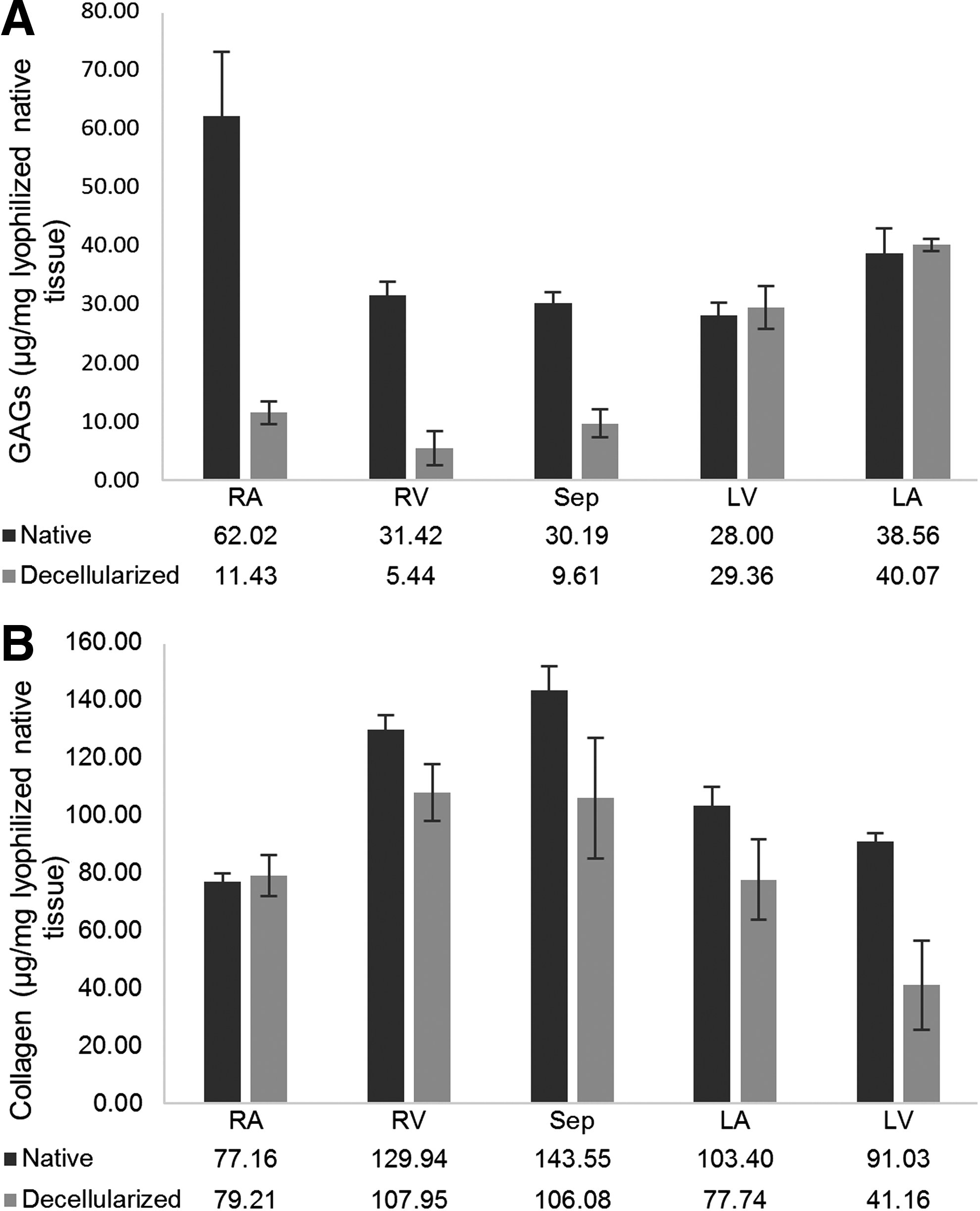
The highest amount of sGAGs in cECM was in samples from the left atrium, with an average of 40 μg/mg (normalized to weight of lyophilized native tissue) and the lowest was found in the right ventricle samples. The ratio of GAGs in cECM to native samples was 0.17, 0.18, and 0.32 for the right atrium, right ventricle, and septum, respectively, while there was no statistically significant difference between cECM and native samples in the left atrium and ventricle.
Collagen
The amount of soluble collagen was measured in decellularized (n=5) and native (n=5) sample biopsies from different sections of three decellularized hearts using the Sircol® Soluble Collagen Assay Kit. The normalized amounts of soluble collagen in different sections of the native and decellularized hearts (Fig. 4B) showed a decrease in the amount of collagen in the right ventricle, septum, left atrium, and left ventricle after decellularization. No statistical significant (p>0.05) difference was observed in the right atrium between the decellularized and native hearts. Although not statistically significant in all groups, collagen preservation tended to be higher in the right portion of the heart than the left heart.
Corrosion casting and SEM imaging
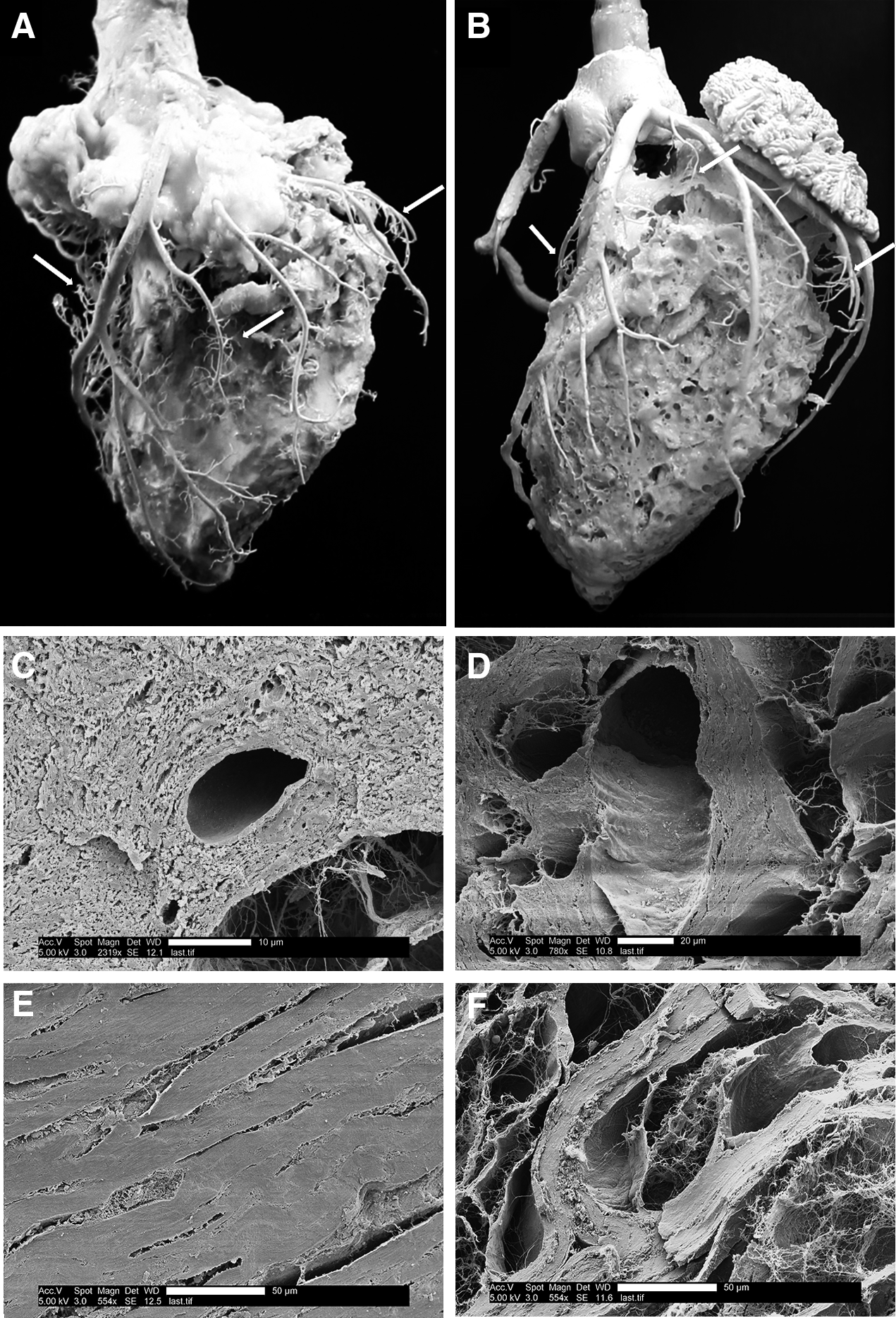
Casts of the vascular network of native and automated pressure controlled decellularized porcine hearts are shown in Figure 5A and B, respectively. The casts were created by retrograde perfusion of polyurethane resin and catalyst through the coronary blood vessels, curing the polymer, and then digesting the heart tissue with NaOH. The resulting cast produced a 3D model of the vascular network that could be used for visual examination of the coronary blood vessels. Care was used when handling the casting during and after processing since the hardened resin was brittle and the structures contained microvessels. As shown in Figure 5B, the vasculature of the cECM was preserved during the decellularization process.
Polyurethane vasculature cast of native
SEM imaging was used for microstructural analysis of the cECM and comparison with the native tissue. As shown in Figure 5C–F, the decellularization process increased the porosity of the tissue by removing the cellular components but preserved the protein fibril network. The structure of the cECM was observed to be similar to the native heart samples in both left atrium (Fig. 5C, D) and left ventricle (Fig. 5E, F) and the coronary blood vessels remained intact.
Mechanical properties
Young's elastic modulus was determined by plotting compression stress as a function of strain in decellularized and native heart samples and finding the slope of the graph at linear portions at both low and high strains. The results are shown in Figure 6A and B. While the results were not statistically significant in the right and left atria, applying relatively small amounts of pressure tended to result in more deformation in the cECM samples than native hearts (Fig. 6A) in all groups. This may be because during decellularization the ECM remained the same relative size as the cellular debris was removed, and therefore, it became more hollow and easier to deform. Consequently, the cECM samples tended to have a smaller Young's elastic modulus than native hearts.

Elastic moduli of decellularized and native samples from different sections of the heart at low
As shown in Figure 6C–G, native samples reached the ultimate resistance force of 45 N earlier at lower strain than decellularized cECM samples. Although the elastic moduli demonstrated a more accelerated increase in the native samples, cECM samples showed higher elastic moduli (Fig. 6B) under high pressure, which may have been due to the incompressibility of water.
Cytotoxicity
Culture of MS-1 cells on tissue culture plastic and in the presence of cECM followed by a Live/Dead assay showed no indication of the cECM causing cytotoxicity. Cells seeded with and without cECM reached full confluency in a similar time frame (3 days) and the quantities of healthy and deceased cells were similar in both control and cECM groups (Fig. 7A–D). These results confirmed that MS-1 endothelial cells were able to proliferate in the presence of cECM with comparable results to traditional cell culture conditions.

Recellularization
Samples from the left ventricular myocardium of decellularized hearts were reseeded with DiI-labeled HCF. HCF were visualized with fluorescence excitation at 549 nm and emission at 565 nm to acquire fluorescence images from the samples. As shown in Figure 7E, cells proliferated and attached to the cECM.
Samples from the inner left ventricular wall of decellularized hearts were recellularized with GFP-positive MS-1 endothelial cells labeled with the lipophilic tracer DiO. Fluorescence images from the reseeded samples were obtained with 484 nm excitation and 501 nm emission wavelengths. MS-1 cells were observed to adhere to cECM and proliferate, covering the surface of the decellularized tissue (Fig. 7G). Histological images with H&E staining showed adherence and integrity in the reseeded cECM samples (Fig. 7F, H).
Thrombosis
In vitro static blood thrombosis assays were performed using 1.5 U/mL heparinized fresh bovine blood on native and decellularized porcine and bovine hearts to assure that the cross-species properties of blood and cECM did not interfere with the experimental results. After 4 h of static blood exposure, the hearts were perfused with a 1× PBS solution. Blood clots emerged from the coronary sinus and venous sections of the decellularized hearts, but not from the native hearts.
All of the hearts were then dissected and the adherence of platelets to the coronary vessels and inner walls of the heart was visually analyzed as described by Sukavaneshvar et al. 30 Although no clot formation was observed in any parts of the native hearts (Fig. 8A, C, E, G), the coronary sinuses and arteries of the decellularized hearts were clogged with blood clots. Platelets adhered to the inner walls of the right and left portions of the decellularized porcine and bovine hearts as shown in Figure 8B, D, F, and H. As shown in Figure 8F and H, significant thrombosis occurred inside the left and right portions of the decellularized bovine hearts and the nonadherent clots had to be removed for visual observation of the platelet adherence to the decellularized tissues.

Thrombosis assay results on porcine
Discussion
Decellularization of whole porcine hearts was accomplished with 6 h of SDS and 2 h of Triton X-100 washes. When compared to previous studies, this process included less detergent exposure while preserving the chemical and mechanical integrity of the ECM.16,17,31 It has previously been shown that detergents affect the basement membranes of biologic scaffolds, and SDS has the most destructive effect among various ionic and nonionic detergents. 18
However, to remove cellular material and avoid inflammatory response at the time of transplantation, the use of SDS seems to be an essential step to strip the ECM of any cellular components.31,32 Therefore, it was decided to use SDS in an optimized manner to preserve the vital components of the ECM while removing immunogenic cellular materials, resulting in more fully decellularized hearts without the potential for an immunogenic response due to residual cell debris.
The DNA content can be used to determine the efficiency of the decellularization process in terms of removing cells and immunogenic factors. 24 It is important to note that the reported amounts of DNA in native tissues vary greatly in the literature, as shown in Table 1, which requires reporting the percentage of DNA removal as well as residual DNA content after processing. In this study, more than 98% of DNA was removed using an automated pressure controlled process.
HUV, human umbilical vein.
Collagen is responsible for mechanical properties and acts as a signaling molecule for cell differentiation and proliferation. 39 Among the various components of the ECM, collagen is the most abundant, with more than 20 types. In a previous study, Naugle et al. showed that type VI collagen induced myofibroblast differentiation and type I and III collagen improved proliferation of cardiac fibroblasts. 40 By both collagen quantification (Fig. 4C, D) and general collagen staining (Fig. 2E, F), our decellularization results demonstrated that collagen was highly preserved.
GAGs are another fundamental component of the ECM that needs to be preserved. GAGs bind growth factors and keep water within the structure of the ECM. 39 It has also been demonstrated that GAGs are susceptible to damage by detergents 41 and dissociating enzymes. 42 Less SDS exposure from our decellularization method increased the preservation of GAGs within the structure of the ECM compared to a previous study. 34
Mechanical properties of the heart, as a continuously beating organ, are crucial for proper function. Maintaining the mechanical strength and stiffness of the cECM is one of the end goals of whole heart decellularization. However, the stiffness of the ECM can also dictate differentiation of stem cells.43,44 Pek et al. showed ECMs with elastic moduli ranging from 0.1 to 34 kPa changed the differentiation of mesenchymal stem cells from a neural phenotype to an osteogenic phenotype. 45
Previous decellularization reports of rat17,46 and porcine47–49 hearts demonstrated an increase in the stiffness (i.e., elastic modulus) of decellularized scaffolds compared to their native controls from tensile tests that stretched the scaffolds in various directions. This increase in the stiffness was believed to be due to the denser and more compact matrix that is a result of decellularization. 48 In our study, compression tests were performed and it was shown that at low compression strains, native tissues demonstrated higher stiffness; however, under high compression strains, elastic moduli of the cECM samples were higher or equal to the native hearts, which is similar to previous reports.
It is known that decellularized porcine heart valve matrices induce human platelet activation and cause thrombosis and inflammatory problems. 50 In our work, whole decellularized porcine and bovine hearts were shown to induce platelet activation and adhesion, while native hearts were found to be highly resistant to thrombus formation (Fig. 8). This is likely due to the presence of the endothelial cell layer on the inner surface of the endocardium.
As previously described by Robertson et al., 31 re-endothelialization of the decellularized rat heart with rat neonatal cardiomyocytes diminished the thrombogenicity of the scaffolds in vitro and in vivo, as well as increased the beating function of recellularized hearts. Therefore, recellularization with endothelial cells attached to the blood contacting surfaces of the ECM is crucial for whole organ engineering, and further studies need to be performed to reduce the thrombosis and inflammatory stimulation of decellularized organs.
The use of decellularized organs has been proposed by several groups for whole organ engineering. For example, in 2008, Ott et al. succeeded in creating functional beating rat hearts by decellularization of native hearts with 12-h retrograde perfusion of 1% SDS and recellularization with neonatal cardiomyocytes and endothelial cells. 17 A report on recellularization of decellularized mouse hearts with human iPSCs by Lu et al. 37 included a modified retrograde perfusion decellularization method in less than 2 h using enzymatic solutions, 1% SDS, 3% TX-100, followed by 4% sodium deoxycholate (SDC). The hearts were recellularized with multipotential cardiovascular progenitors derived from Y1-iPSCs and after 20 days of culture and differentiation, beating whole mouse hearts were achieved.
An optimized automated method was reported by Akhyari et al.
38
and Aubin et al.
51
for decellularization of rat hearts using 1% SDS (w/v), 1% SDC (w/v), 0.05% NaN3 in deionized water, 1% saponin, 0.05% NaN3 solutions, and implementing a computer pressure control system through negative feedback and engineo GmbH software (
The first report of porcine heart decellularization was in 2010 by Wainwright et al., 34 in which porcine hearts were decellularized using 2× PBS, 0.02% Trypsin in EDTA and NaN3, 3% TX-100, and 4% SDC in 8 h resulted in 92.2% DNA removal. It was reported that the amounts of elastin and GAGs per 1 mg of lyophilized sample were not different than those measured in the native hearts. However, because of the change in the density of samples through decellularization, elastin and GAGs were lost at a ratio similar to the overall loss in dry weight.
Remlinger et at. 35 in 2012 used the same decellularization method while gradually increasing the flow rate and implementing a maximum of 2200 mL/min flow rate. In our experiments, the pressure increase was also automated and controlled by the computer to a maximum flow rate of 2250 mL/min to maintain the set point pressure.
Ultimately, the use of stem cells or iPSCs would be desirable for recellularization. Therefore, retaining fundamental molecules to signal cell proliferation and differentiation will be required to mimic the function of healthy hearts. A gentle decellularization method is a promising process to obtain intact ECM ready for repopulation with stem cells. Further studies need to be performed to demonstrate the differentiating capacity of these cECM prototypes for human stem cells.
Conclusion
Porcine hearts were successfully decellularized using an automated pressure controlled apparatus. The resulting cECM contained low amounts of DNA and limited potential for causing an immune response while preserving the GAGs and collagen proteins. The cECM samples exhibited a high capacity for recellularization. The next steps include recellularization with human cells, culturing the 3D organs in bioreactors, and testing the hearts for appropriate beating function and nonthrombogenicity.
Footnotes
Acknowledgments
The authors acknowledge the contributions of Makena Ford, Andrew Priest, Camille Brantly, Holly Howarth, and Brielle Woolsey for their laboratory work. Ryan J. Morris, Jason R. Gassman, and Tyson R. Jergensen assisted with histology. Kevin Cole assisted with automation of the apparatus and Dr. Sivaprasad Sukavaneshvar assisted with the static blood assay. BYU is gratefully acknowledged for providing funding.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.