
Abstract
Studies on hematopoiesis currently rely on animal models because in vitro culture methods do not accurately recapitulate complex bone marrow physiology. We recently described a bone marrow-on-a-chip microfluidic device that enables the culture of living hematopoietic bone marrow and mimics radiation toxicity in vitro. In the present study, we used this microdevice to demonstrate continuous blood cell production in vitro and model bone marrow responses to potential radiation countermeasure drugs. The device maintained mouse hematopoietic stem and progenitor cells in normal proportions for at least 2 weeks in culture. Increases in the number of leukocytes and red blood cells into the microfluidic circulation also could be detected over time, and addition of erythropoietin induced a significant increase in erythrocyte production. Exposure of the bone marrow chip to gamma radiation resulted in reduction of leukocyte production, and treatment of the chips with two potential therapeutics, granulocyte-colony stimulating factor or bactericidal/permeability-increasing protein (BPI), induced significant increases in the number of hematopoietic stem cells and myeloid cells in the fluidic outflow. In contrast, BPI was not found to have any effect when analyzed using static marrow cultures, even though it has been previously shown to accelerate recovery from radiation-induced toxicity in vivo. These findings demonstrate the potential value of the bone marrow-on-a-chip for modeling blood cell production, monitoring responses to hematopoiesis-modulating drugs, and testing radiation countermeasures in vitro.
Introduction
T
Various methods have been developed to culture hematopoietic cells and produce specific blood cell types in vitro.6–12 These culture systems, however, do not model the bone marrow microenvironment13,14 in sufficient complexity to allow the investigation of radiation responses and radiation countermeasures in a physiologically relevant context. To overcome this barrier, we recently developed a bone marrow-on-a-chip microfluidic device that supports intact living bone marrow as a functional hematopoietic niche in vitro, and we showed that it produces a radiation injury response that is highly similar to those observed in living marrow in vivo. 15 In this method, a tissue engineering approach is used to induce formation of new marrow-containing bone in mice, and the bone is surgically removed, placed in a microfluidic device, and continuously perfused with medium in vitro. In this study, we explore whether this microfluidic device can be used to study continuous production of blood cells and to test pharmacological countermeasures that might help radiation-injured bone marrow repair itself. To test the utility of the system, we analyzed the dynamics of blood cell production of a 13-day culture period, and we evaluated the radiation-protecting effects of two potential therapeutic proteins, granulocyte-colony stimulating factor (G-CSF)16,17 and bactericidal/permeability-increasing protein (BPI), 5 which have been reported to accelerate recovery of hematopoiesis after radiation-induced bone marrow failure in vivo.
Materials and Methods
Bone marrow engineering in vivo
Our methods for engineering bone with functional marrow in vivo, and for harvesting the engineered bone marrow (eBM) so that it can be cultured in a microfluidic bone marrow-on-a-chip device in vitro, have been previously described. 15 In brief, 3 mg of demineralized bone powder prepared from femurs harvested from CD-1 mice was mixed with 30 μL solution of type I collagen gel (3 mg/mL, Cellmatrix Type I-A; Nitta Gelatin, Inc.), 100 ng BMP2 (Alpha Diagnostic Intl., Inc.), and 100 ng BMP4 (Alpha Diagnostic Intl., Inc.) and placed in the central cylindrical cavity (1 mm high × 4 mm diameter) of a device, which was fabricated from polydimethylsiloxane (PDMS). The PDMS devices filled with the bone-inducing materials were implanted subcutaneously on the backs of 8- to 12-week-old C57BL/6 mice and harvested 8 weeks after implantation. CD-1 mice were purchased from Charles River Laboratories, and C57BL/6 mice were purchased from Jackson Laboratories. All animal studies were reviewed and approved by the Animal Care and Use Committee of Children's Hospital Boston.
Microfluidic bone marrow-on-a-chip
The eBM was removed from the PDMS device and cultured in a similarly shaped central chamber within a microfluidic chip device (Fig. 1A) that was separated from an underlying microfluidic channel (200 μm high) by a porous PDMS membrane (20 μm thick with 100 μm pores). Culture medium (serum-free expansion medium [SFEM] basal medium; STEMCELL Technologies) supplemented with 10% fetal bovine serum (FBS), penicillin (100 U/mL), streptomycin (100 U/mL), and cytokines (50 ng/mL mouse stem cell factor (SCF), 100 ng/mL mouse FLt-3; STEMCELL Technologies) was perfused through the bottom to the top channels (2 μL/min) after the eBM was inserted into the central chamber, pierced multiple times with a surgical needle (32 gauge), and the overlying microchannel layers were attached. eBM was cultured on-a-chip for up to 2 weeks and outflow was collected every 3 days following 1 day of culture to analyze blood cell production. To evaluate the production of erythrocytes, 2 U/mL erythropoietin (EPO; Cell Sciences) was added in the cytokine-containing culture medium after 1 day of culture and included in the medium for 12 additional days. To image blood cells, cells in outflow were stained with modified Wright-Giemsa stain (Diff-Quik; American Scientific Products) after cytospin preparations. To harvest bone marrow cells, the eBM was removed from the PDMS devices, cut into small pieces, and digested using 1 mg/mL collagenase (Roche) for 30 min. The number of cells in outflow was counted using a hemocytometer.
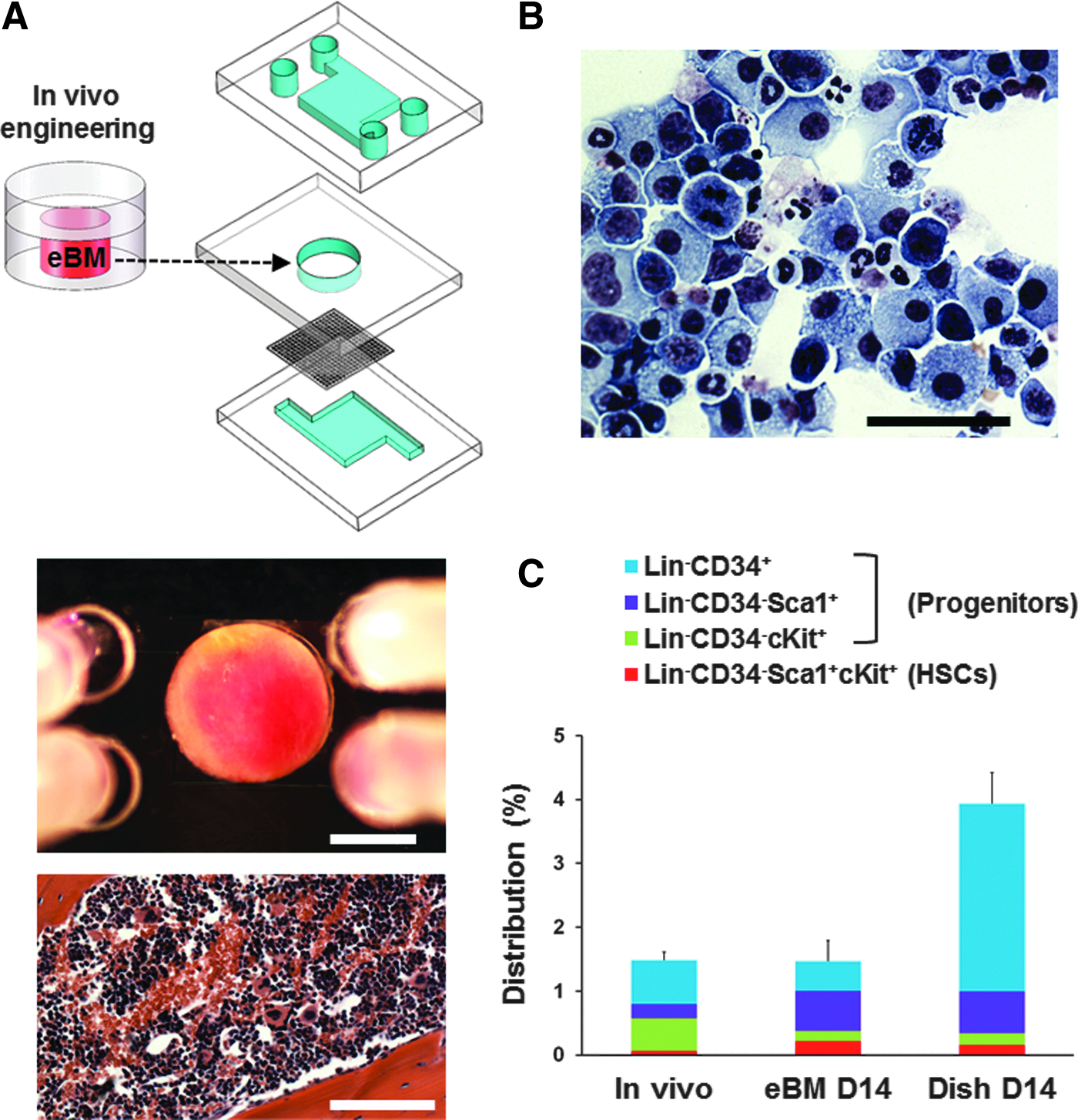
Bone marrow-on-a-chip microdevice.
Static bone marrow culture
To produce conventional static marrow cultures, 5 million bone marrow cells harvested from a femur of a C57BL/6 mouse were cultured in a tissue culture plate (Falcon) using the same medium used in the microfluidic device. Half of the culture medium was replaced with fresh medium every 3 days, and ∼90% of cells remained viable after 14 days in culture.
Flow cytometry
To evaluate the distribution of the various hematopoietic cell populations, flow cytometric analysis was performed using 5-laser-Fortessa flow cytometer (Becton Dickinson). 15 Cell types were evaluated based on expression of surface antigens that are characteristic for hematopoietic stem cells (HSCs), hematopoietic progenitor cells, and multiple differentiated blood cell lineages. Cells were stained in cold phosphate-buffered saline containing 3% FBS for 60 min with antibodies directed against (i) eFluor 450 hematopoietic lineage cocktail (1:5, #88-7772-72; eBioscience), APC Sca1 (1:333, #17-5981-82, clone D7; eBioscience), APC-eFluor780 cKit (1:160, #47-1172-82, clone ACK2; eBioscience), and FITC CD34 (1:50, #11-0341-82, clone RAM34; eBioscience) to identify HSCs (Lin−Sca1+cKit+) and progenitor cells (Lin−Sca1+, Lin−cKit+, Lin−CD34+); (ii) PerCP-Cyanine 5.5 Ter119 (1:40, #45-5921-82, clone TER-119; eBioscience), Pacific Blue CD45 (1:200, #48-0451-82, clone 30-F11; eBioscience), APC-eFluor780 CD3 (1:20, #47-0032, clone 17A2; eBioscience), APC CD19 (1:160, #17-0193, clone 1D3; eBioscience), FITC Mac1 (1:100, #11-0112-82, clone M1/70; eBioscience), and FITC Gr1 (1:400, #11-5931-82, clone RB6-8C5; eBioscience) to identify erythrocytes (Ter119), leukocytes (CD45), B cells (CD19), T cells (CD3), and myeloid cells (Mac1 and Gr1). Cellular viability was evaluated using 7-AAD viability staining solution (#00-6993-50; eBioscience).
Radiation exposure
Freshly harvested eBM made in C57BL/6 mice or freshly isolated mouse femur bone marrow cells (5 × 106 cells) were exposed to 4 Gy dose of γ-irradiation (Cs-137) and then, respectively, cultured either in the microfluidic device or on standard tissue culture plates for 2 weeks. Outflow from the microfluidic device was collected every 3 days following the first day of culture for flow cytometric analysis. Five hundred units per milliliter mouse G-CSF (STEMCELL Technologies) or 1 μg/mL recombinant human BPI was added in the cytokine-containing culture medium 1 day after exposure to γ-radiation and these factors were maintained in the medium at the same concentration for nine additional days. The recombinant human BPI was generated as a Fc fusion protein, BPI-Fc, which consists of the N-terminal domain of human BPI (residues 1–193 of the mature protein) followed by the hinge, CH2 and CH3 domains of human IgG1 Fc residues E216-G446 with two point mutations (C220S and N297D), and a single terminal alanine residue. This BPI-Fc was expressed in 293F cells (Invitrogen) and purified by protein A affinity; analysis of the purified protein confirmed that it was ∼100 kDa molecular weight as expected, when analyzed by SDS-PAGE with Coomassie Fluor Orange (Invitrogen).
Statistics
Statistical differences were analyzed by Student's t-test. All statistical evaluation was conducted using a two-tailed t-test, assuming independent samples of normal distribution with equal variance. Significant differences with p < 0.05 were deemed significant; all error bars indicate standard error of the mean.
Results
In vitro blood production in the bone marrow-on-a-chip
To explore whether we could model marrow responses to therapeutic countermeasures to radiation poisoning, we used the microfluidic bone marrow-on-a-chip to determine whether it can support sustained blood cell production over time in vitro. The chip was created by first forming bone containing marrow within a cylindrical cavity formed in a piece of PDMS that was implanted subcutaneously for 8 weeks in living mice, and then surgically removing the formed boney disk, puncturing the bone in multiple sites with a needle, and inserting it into a similar shaped cavity of a second PDMS microfluidic chip device that was perfused with the hematopoietic supporting medium (Fig. 1A). We previously demonstrated that the bone marrow-on-a-chip can maintain HSCs and hematopoietic progenitor cells in normal in vivo-like proportions for 7 days in culture. 15 In this study, we cultured the chip for 2 weeks and confirmed that the distribution of HSCs and progenitors in the bone marrow microdevice maintained themselves in similar normal proportions for the entire period of culture (Fig. 1B, C). In contrast, the relative proportions of these cells changed significantly when freshly isolated marrow was cultured for the same time in conventional static dish culture that lacks the bone marrow microenvironment (Fig. 1C).
We then collected the outflow of the microfluidic channel of the device every 3 days following the first day of culture to analyze production and release of blood cells into the microfluidic circulation. When the cells collected from the outflow were pelleted and stained with Wright-Giemsa, we detected various types of blood cells, including macrophages, neutrophils, and lymphocytes (Fig. 1B). Cells with similar morphology were observed continuously over a 13-day period of culture confirming that this bone marrow on-a-chip continues to produce differentiated blood cells and release them into microfluidic circulation. The number of cells in the outflow collected in subsequent intervals gradually increased over time and reached a maximum of about 1 million cells between day 7 and day 10 after which remained stable over time through day 13 (Fig. 2A). Thus, the bone marrow-on-a-chip supports continuous production of blood cells for at least 13 days in culture while maintaining HSCs and progenitors in normal proportions inside the microdevice. When the cells released into circulation were analyzed using flow cytometry, we detected HSCs (Lin−Sca1+cKit+), hematopoietic progenitors (Lin−CD34+), erythrocytes (Ter119+), and lymphoid cells (CD45+CD3−/+CD19−/+) in the outflow; however, most were myeloid cells (CD45+Mac1−/+Gr1−/+) (Fig. 2B–F). The number of all blood cell types increased during the 13-day culture with the exception of erythrocytes, which gradually decreased over time (Fig. 2D). This is not surprising because we used medium in this study that is designed to preferentially support survival of HSCs and hematopoietic progenitors.
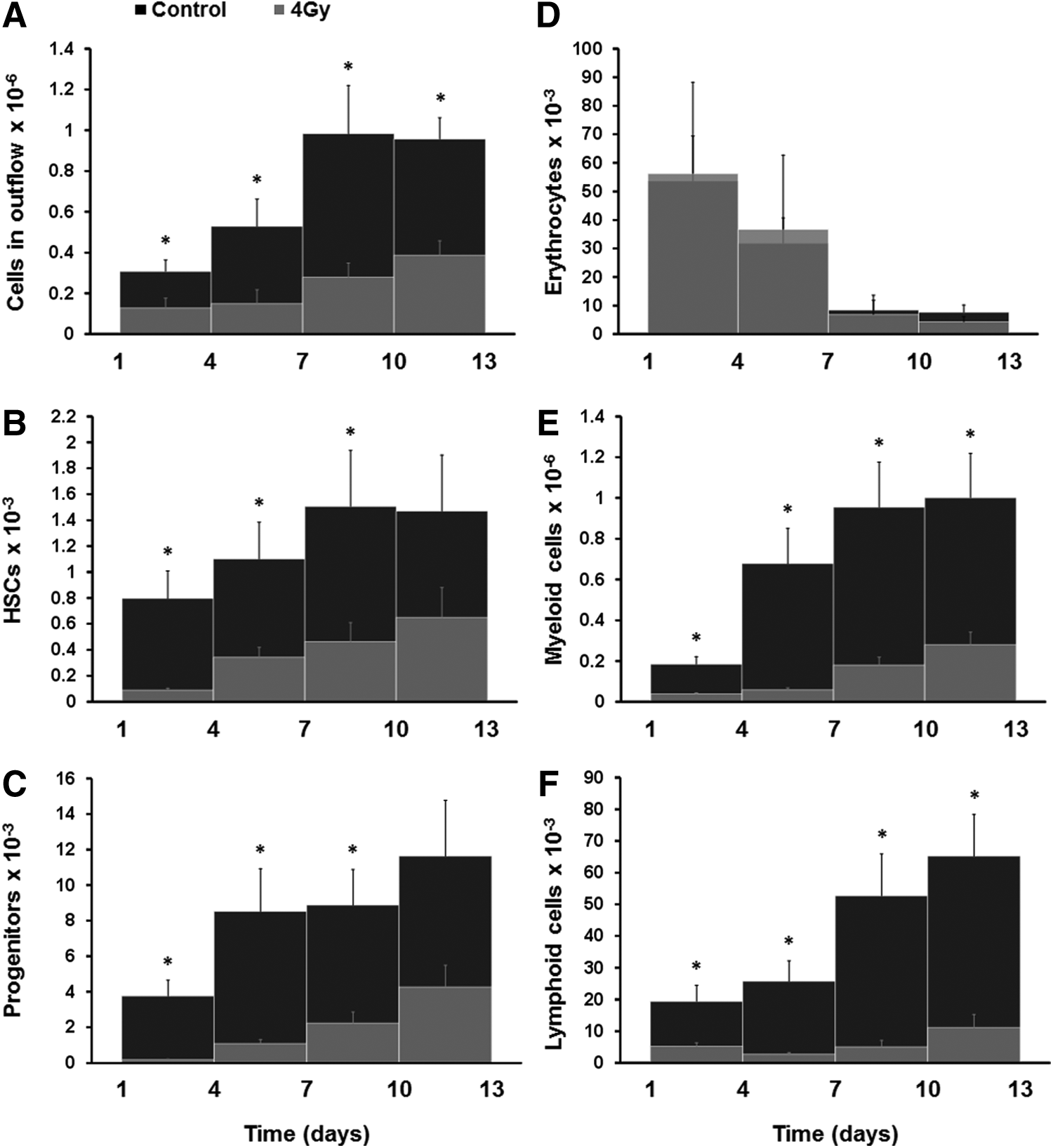
Cells in outflow of the bone marrow-on-a-chip.
We then analyzed the effect of exposing the eBM to 4 Gy of γ-radiation before placing them in culture on the microfluidic chip. The total number of cells, as well as the number of HSCs, progenitor, myeloid, and lymphoid cells in the outflow, was significantly lower than that in chips containing control unirradiated bone marrow (Fig. 2A–C, E, and F); however, the number of erythrocytes remained virtually unchanged (Fig. 2D). In addition, after radiation exposure, the HSCs and progenitor cell levels recovered more quickly than the mature blood cell types, whereas again the erythrocytes did not appear to be sensitive to radiation exposure, as suggested by previous reports.
18
In contrast, the total number of cells and proportion of different blood cell types contained within both the control and 4 Gy-exposed eBM remained constant at ∼5 million cells when measured after 2 weeks on-chip (Supplementary Fig. S1A, B; Supplementary Data are available online at
Testing of drugs that alter blood cell production
Although the culture medium was not optimal for supporting red blood cell production, we were able to induce red blood cell production in the bone marrow chip by adding EPO, which is known to stimulate erythrocyte formation. 19 When EPO was added after 1 day of culture, we detected a continuous and significant (4.4-fold at day 9) increase in the number of erythrocytes released in the outflow over the 13-day culture (Fig. 3A). When the bone marrow-on-chip was exposed to a 4 Gy dose of γ-radiation on day 0, the effect of EPO on the number of erythrocytes was significantly lower and it took more time to induce erythrocyte production (Fig. 3B). However, we still detected a significant increase in the production of erythrocytes by day 13, suggesting that radiation exposure had damaged, but not fully eradicated the erythroid progenitors. The bone marrow-on-a-chip may therefore be useful for screening of new and more effective drugs that can reinitiate red blood cell production after radiation injury.

Effect of EPO on blood cell production. Effect of EPO in the number of erythrocytes (Ter119+) in outflow without
Testing radiation countermeasure drugs on-chip
To provide proof-of-concept for the microfluidic device as a platform for evaluating the effects of radiation countermeasures on the hematopoietic compartment, we administered either G-CSF or BPI, which are both known to accelerate hematopoietic recovery after bone marrow failure caused by radiation exposure.5,16,17 When G-CSF was added to the microdevice 1 day after exposure to a 4 Gy dose of γ-radiation and was maintained in the medium for nine additional days, we detected significant increases in the number of HSCs (Lin−Sca1+cKit+), progenitors (Lin−CD34+), and myeloid cells (CD45+Mac1−/+Gr1−/+) 3–6 days after the addition of G-CSF (Fig. 4). Treatment with G-CSF resulted in 5.1-, 5.4-, and 4.0-fold increases in the number of HSCs, progenitors, and myeloid cells after 6 days, respectively; however, there was no significant difference in the number of lymphoid cells (CD45+CD3−/+CD19−/+). When BPI was added to the bone marrow chip, it also significantly (∼twofold) increased the number of HSCs and myeloid cells within 6 days (Fig. 5A). Thus, these radiation countermeasure drugs were both able to accelerate recovery of blood cell production after radiation injury in the bone marrow-on-a-chip in vitro, just as they have previously been reported to do in vivo.5,16,17
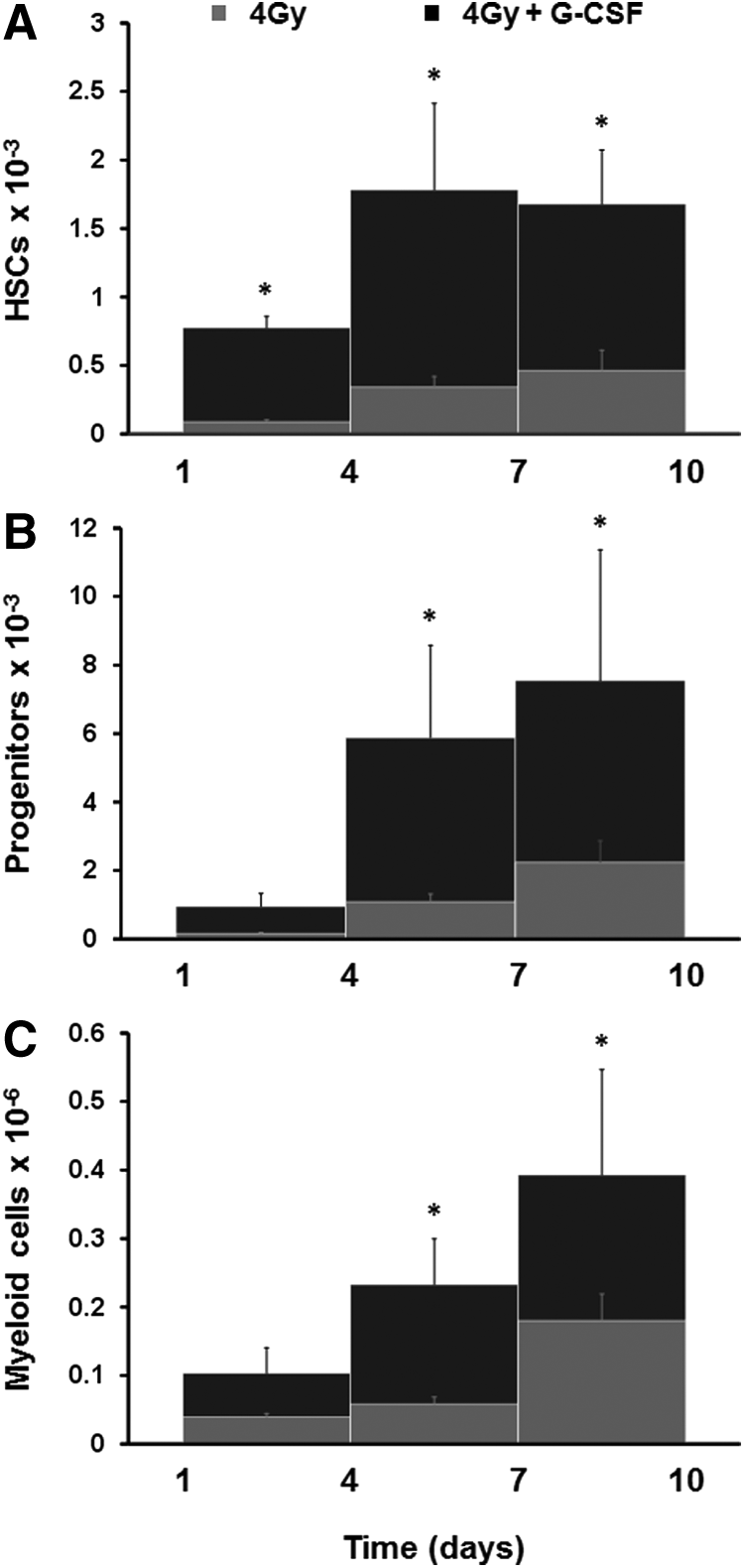
Effect of G-CSF on blood cell production after radiation exposure. Effect of G-CSF in the number of HSCs (Lin−Sca1+cKit+)
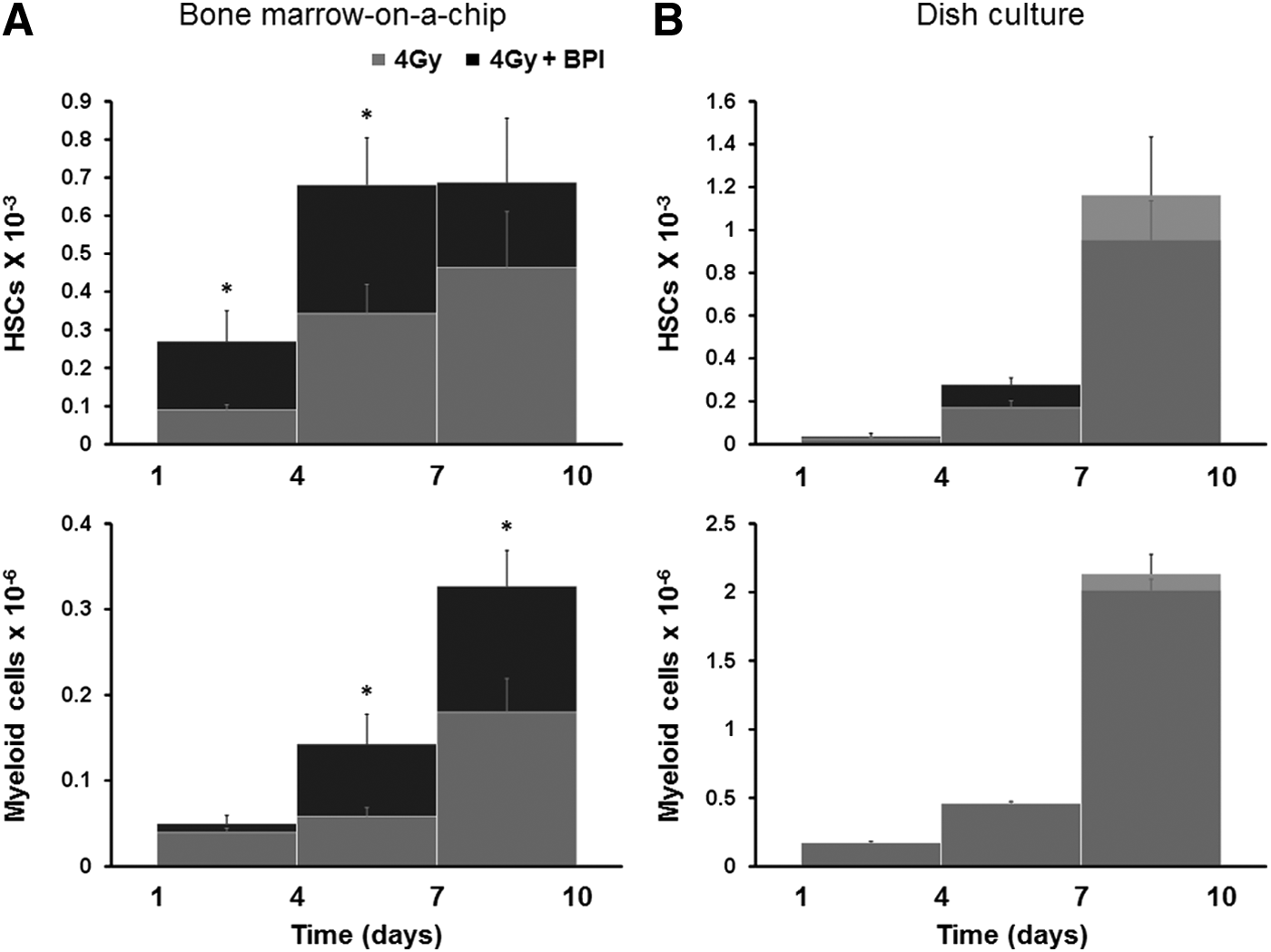
Effect of BPI on blood cell production after radiation exposure. Effect of BPI in the number of HSCs (Lin−Sca1+cKit+) and myeloid cells (CD45+Mac1−/+Gr1−/+) in outflow of the bone marrow-on-a-chip microdevice
To further investigate the novelty of this microfluidic device, we compared our results with those obtained using a conventional static culture, in which bone marrow cells freshly harvested from mouse femur are maintained on plastic dishes in the same culture medium used in the chip. Importantly, when the dish culture was exposed to 4 Gy dose of γ-radiation, it was not able to detect any effect of BPI, whereas the protective effects of this known countermeasure drug were detected in the microfluidic device (Fig. 5A vs. B). Very simply, the bone marrow-on-a-chip effectively mimics the in vivo response of living bone marrow to radiation countermeasure drugs, whereas the conventional static bone marrow cultures do not.
Discussion
Organ-on-a-chip microfluidic devices produce organ-level functions not possible with conventional in vitro culture methods by recapitulating tissue architecture, tissue–tissue interfaces, and physicochemical microenvironments.20,21 This technology has great potential to facilitate drug discovery and development and to replace animal models for efficacy and toxicity testing. An artificial bone marrow that recapitulates bone marrow structure and function in vitro could be a useful platform to test new therapeutics, as well as study hematopoiesis and hematologic diseases. We recently developed a method for fabricating a bone marrow-on-a-chip microdevice, which permits the culture of intact living bone marrow containing a functional hematopoietic niche in vitro by combining tissue engineering approaches with microfluidic techniques. 15 Using bone engineering, it is possible to control the shape, size, and thickness of the bone surrounding the marrow, which facilitates culture and maintenance of marrow cell viability compared to attempting to culture native marrow surrounded by thick cortical bone in vitro.
In the present study, we extended this work by showing that this bone marrow-on-a-chip can produce blood cells continuously and release them into the microfluidic circulation, while maintaining HSCs and hematopoietic progenitor cells in normal in vivo-like proportions inside the microdevice for at least 2 weeks in culture. Because it takes about 1 week to differentiate into mature blood cells in vivo, it is necessary to monitor blood production for more than 1 week to evaluate effects of drugs that modulate hematopoiesis.16,22 Thus, any culture system that is used for long-term analysis of blood cell formation must maintain the viability and function of HSCs, as well as their ability to differentiate and produce mature blood cells in vitro. 23 This bone marrow-on-a-chip was able to do both, and when we administered EPO, we could demonstrate induction of red blood cell formation by this clinically approved therapeutic. This finding confirms that this microfluidic device can be used effectively to study red blood cell production in vitro.
The bone marrow microdevice produces an organ-level response to radiation toxicity that is similar to observed in vivo. 15 Importantly, in this study, we found that while addition of either G-CSF or BPI induced proliferation of HSCs and myeloid cell production in the bone marrow chip, we could not detect any protective effect of BPI in static marrow cultures, even though BPI has been previously shown to accelerate recovery from radiation-induced toxicity in vivo. 5 These results suggest that the presence of a hematopoietic microenvironment (as found in the bone marrow chip) is crucial for modeling radiation toxicity and testing effects of potential countermeasure drugs in vitro.
In fact, it is precisely because conventional culture methods do not accurately recapitulate the bone marrow microenvironment, which plays such an important role in hematopoiesis, as well as radiation responses,2,24,25 that drug development studies currently rely on animal models for efficacy and safety testing. Because our bone marrow-on-a-chip device contains an intact marrow microenvironment and it is able to recapitulate physiological responses to radiation countermeasure drugs, it could offer a novel replacement to animal testing in this area. The use of microfluidics also supports maintenance of cellular viability and function, in addition to enabling analysis of physiological responses under flow. It also might be possible to generate a human bone marrow-on-a-chip by engineering bone marrow in immunocompromised mice and replacing their marrow with human hematopoietic cells in the future. Another possibility is that marrow cells inside the eBM might be inactivated by radiation exposure and then the vacated bone marrow niche could be repopulated with human hematopoietic cells. The bone marrow-on-a-chip also offers the ability to integrate other cell types, such as cancer cells, into this microenvironment to model hematologic or metastatic diseases. Therefore, this microphysiological system has the potential to serve as in vitro human replacement for animal models.
Footnotes
Acknowledgments
The authors thank Eva C. Guinan for helpful input relating to the use of BPI. This work was supported by the Wyss Institute for Biologically Inspired Engineering at Harvard University, the Defense Advanced Research Projects Agency under Cooperative Agreement Number W911NF-12-2-0036, and the U.S. FDA HHSF223201310079C.
Disclosure Statement
D.E.I. holds equity in Emulate, Inc. and chairs its Scientific Advisory Board.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.