
Abstract
Sheep are one of the most frequently used large animal models in stem cell research. However, minimal invasive or noninvasive sources of mesenchymal stem cells (MSCs) in sheep are scarce. In the light of the principles of the 3Rs (reduce, refine, replace), it would therefore be desirable to identify a minimally invasive or noninvasive ovine MSC source. In humans, the chorionic villi of the placenta, which can be noninvasively harvested as part of the afterbirth, have been identified as a rich source of MSCs. Therefore, in the present study, ovine placenta cotyledons, which have similar function and structure to human chorionic villi, were tested for their potential use as a noninvasive source of ovine MSCs. Through mincing of the placental cotyledons, collagenase digestion, and Ficoll density gradient centrifugation, combined with plastic adherence selection, MSCs were successfully isolated. Their morphological, immunophenotypical, and cellular growth characteristics, as well as their proliferation, differentiation, and migration potential, were evaluated and compared to the currently best-researched MSC source, bone marrow-derived stem cells. Ovine cotyledons were shown to be a reliable, abundant source for the noninvasive, pain- and risk-free harvest of MSCs. The collection procedure does not interfere with partum or the initial bonding phase between ewes and lambs and is therefore exempt from ethical debate. Ovine placenta cotyledon-derived MSCs exhibit multipotential characteristics and can be cryopreserved for later use.
Introduction
M
Bone marrow and adipose-derived stem cells are the best researched stem cell sources of humans and animals. However, a disadvantage of bone marrow-derived MSCs (bmMSCs) is the invasive harvesting methodology.3,20–26 Adipose tissue is routinely gained through a less but still invasive surgical procedure.21,26–31
Minimal invasive or noninvasive MSC sources in sheep are scarce. Peripheral blood would be a largely pain- and risk-free source of stem cells. However, as in other species,17,32–35 the amount of stem cells contained in ovine peripheral blood is relatively low.34,35 Therefore, there is a need for an abundantly available, reliable, noninvasive MSC source for this frequently used model animal. Perinatal tissues, such as cord blood, umbilical cord tissue, amniotic membranes, or amniotic fluid, are often used as MSC sources.13,15,18,19,36–69 They are abundantly available and can be harvested in a noninvasive, ethically inoffensive manner.40,45,70,71 The cell populations derived from these sources express all typical MSC markers and produce mesodermal progenitor cells with multilineage differentiation potential.37,72 Access to perinatal MSC sources is pain free and bears no risk for the mother or the newborn.70,73–75 Above that, juvenile MSCs derived from perinatal tissues provide several advantages over adult MSCs. They are characterized by a higher proliferation rate, longer life span, better quality, and lower immunogenicity.45,70–72,76 In addition, juvenile MSCs have a broader differentiation capability toward cell types of endo- or ectodermal origin.45,63
A disadvantage of perinatal tissues especially from large animal donors may be the higher risk of bacterial or mycotic contamination due to the nonsterile birth canal and the delivery in a farm environment.44,61 This makes the use of disinfectants on tissue harvest and antibiotics in the cell culture medium mandatory. 61 Therefore, to date, ovine cord blood in sheep was mainly collected during preterm cesarean sections. 43 Also, amnion-derived and amniotic fluid-derived MSCs, which were successfully obtained in sheep, were mainly collected during aseptic surgical approaches.13,15,18,19,36,41,51,52,59,67
In humans the chorionic villi of the placenta (Fig. 1A) have been identified as rich source of MSCs.77–79 It is nowadays agreed that MSCs are of perivascular origin.80–82 They reside in the so-called stem cell niche, which is typically located around blood vessels and offers an environment for MSCs, in which they remain quiescent until internal or external signals stimulate their replication, migration, differentiation, or secretory activity to repair or replace tissue.83–86 Chorionic villi serve as junction between the maternal and fetal blood circulation. 87 Therefore, they contain a lot of blood vessels, which makes them a potentially valuable MSC source. De facto MSCs were confirmed to reside in a vascular/perivascular niche in the placenta chorionic villi. 77

Human and ovine placenta. Schematic drawing of a human placenta
The placenta of ruminants is cotyledonary, nondeciduate, and epitheliochorial in contrast to the discoid, deciduate, and hemochorial human placenta. However, the function and structure of chorionic villi largely correspond to the human counterparts. 87 In ruminants, the chorionic villi are organized in distinct structures called cotyledons (Fig. 1B–F). Cotyledons (fetal part of the placenta) and caruncles (maternal part of the placenta) (Fig. 1B, C) form specialized units known as placentomes.87,88 Following parturition, the chorionic villi are withdrawn from the maternal caruncles and the fetal membranes are expulsed (Fig. 1C).87,88
Ovine placental cotyledons obtained on full-term delivery were, to our knowledge, not yet described as MSC source. As part of the afterbirth, they are abundantly available and harvesting them is not associated with any ethical concerns because they are usually discarded as trash. MSCs from placenta cotyledons would be an easily and truly noninvasively accessible source of sheep MSCs. Therefore, the aim of the present study was to test ovine placenta cotyledons for their potential use as noninvasive source of ovine MSCs and to compare the obtained MSCs to bone marrow-derived MSCs, the currently best-researched stem cell source.
Materials and Methods
Collection of the afterbirth and preparation of cotyledons
All afterbirth samples were collected during the same breading season on full-term delivery of the lambs according to the Vetmeduni Vienna Guidelines for Good Scientific Practice. The afterbirths were collected post partum from Merino ewes of the Vetmeduni Vienna teaching herd. They were left in situ until they were spontaneously passed (one to 3 h post partum). After delivery of the lambs, and after the important first bonding phase between the ewes and lambs, the ewe's tails were covered with a rectal glove and the vulva was cleaned and washed with a soap solution containing iodine. When the afterbirth was passed, it was collected directly into a sterile pot with a lid to protect it from the farm environment and potential fungal spores in the air. For sterility reasons, the part of the afterbirth, which had been hanging out of the vulva after delivery, was discarded. All further sample processing was done in a laboratory environment. The afterbirths were transferred into a 1% iodine solution for 2 min before their dissection with sterile scissors and sterile forceps. Between 8 and 12 cotyledons were harvested per afterbirth (n = 5). In case of twins or triplets, the secundines were not divided and assigned to individual lambs but the afterbirth of the entire litter was harvested in toto. The harvested cotyledons were washed twice with sterile saline (Braun, 0.9% NaCl for infusion) and stored at 4°C in phosphate-buffered saline (DPBS+ Ca, Mg; Lonza) containing 1% pen/strep (10,000 U penicillin/10 mg streptomycin/mL; Sigma Aldrich) and 1% amphotericin B (250 μg/mL; Biochrom) for a maximum of 18 h post partum.
One cotyledon of each afterbirth was immediately placed in buffered 4% formalin (ACM, Herba Chemosan Apotheker AG) for histologic analysis. It was embedded in paraffin and 4 μm slices were cut and subjected to hematoxylin (Thermo Scientific) and eosin staining (Merck). 89
Isolation and culture of cotyledon-derived MSCs
Under a laminar airflow workbench, the cotyledons (between 7 and 11 per afterbirth) were transferred into a Petri dish and all remaining tissues attached to the cotyledons were carefully removed. Then, the cotyledons were minced into small pieces and digested in a 1 mg/mL collagenase (type I; Sigma Aldrich) solution (Dulbecco's modified Eagle's medium [DMEM] low-glucose, without L-glutamine; Lonza) for 1 h under constant stirring at 37°C. The obtained viscous and bloody solution was mixed with medium to stop the collagenase activity and filtered through a cell strainer (100 μm; Greiner BioOne). A density gradient filtration was performed using Ficoll (Ficoll Paque Premium, Ficoll density 1.077 g/mL; GE Healthcare) to separate the mononuclear cell fraction. For this purpose, the filtered suspension was loaded onto the Ficoll in 50-mL falcon tubes (2:1 ratio of cotyledon solution to Ficoll), and centrifuged for 30 min at room temperature at 330 g (without brake). The buffy coat containing the mononuclear cell fraction was carefully transferred into a new 15-mL falcon tube. It was washed with phosphate-buffered saline (PBS) and centrifuged at 400 g for 5 min at room temperature twice. After resuspension in standard culture medium consisting of DMEM (low glucose, without L-glutamine; Lonza), 10% fetal calf serum (FCS, low endotoxin; Sigma Aldrich), 1% L-glutamine (L-alanyl-L-glutamine 200 mM; Biochrom), 1% pen/strep (100 × ; Sigma Aldrich), and 1% amphotericin B (250 μg/mL; Biochrom), the cells were seeded onto T75 culture dishes and cultured in a humidified atmosphere at 37°C and 5% CO2. The following day the cells were washed to remove nonadherent cells and tissue debris. Medium was further changed twice weekly. Passaging was performed on colony confluence for the first passage and later at 80–90% confluence by trypsinization (trypsin 0.05%/EDTA 0.02%; Biochrom). Cells were frozen and stored in liquid nitrogen until further use at the lowest possible passage (P1–2).
Harvest, isolation, and culture of bmMSCs
Bone marrow was harvested postmortem from the iliac crest of three adult sheep, aged 2–4 years, which were euthanized for reasons unrelated to this study. The sheep were placed in sternal recumbency. After surgical skin preparation, the iliac crest was punctured using a Jamshidi biopsy needle (13G; Greiner BioOne). Bone marrow (17.5 mL) was aspirated carefully with a preheparinized syringe (2.5 mL heparin in 20 mL syringe) and taken to the laboratory for further processing. To isolate the mononuclear cell fraction, a density gradient filtration was performed using Ficoll as described above. MSCs were cultured, frozen, and stored in liquid nitrogen until further use at the lowest possible passage (P1–2) as described above.
Cell proliferation
Population doubling rates per day and generation times were evaluated for cotyledon-derived MSC (cotMSCs, n = 5) and bmMSCs (n = 3) samples. For that purpose, cells were seeded at a defined density (3000 cells/cm2). Care was taken that the proliferation assay was carried out in the same passage (P1) for cotMSCs and bmMSCs. On 80–90% confluence, cells were trypsinized and counted using an automatic cell counter (Countess II FL; Invitrogen). Population doubling rates (PD) and generation times (GT) were calculated according to the following formulas37,90:
PD: (cell count harvest/cell count seeding)/ln (2)
PD/day: PD/day in culture
GT: 1/(PD/day in culture)
For statistical analysis, a t-test for two groups with different variances was performed using IBM SPSS Statistics 24.
Migration assay
To confirm their migration potential, a scratch migration assay was performed for cotMSCs (n = 5) and bmMSCs (n = 3). On a six-well plate, 3 × 105 cells/well were seeded in triplicate and cultured until subconfluence. Care was taken that the cotMSCs and bmMSCs used were in the same passage (P3). On subconfluence, a scratch was created using a pipette tip (20–200 μL). Pictures of the scratch area were taken every 6 h for 48 h using an automatic cell imaging system (EVOS® Auto FL Imaging System; Thermo Fisher Scientific). Based on the pictures, the time until complete closure of the scratch was determined and the migration capability of cotMSC and bmMSC samples was confirmed.
MSC differentiation
To prove their trilineage differentiation potential, cotMSCs (n = 5) and bmMSCs (n = 3) were differentiated into the chondrogenic, adipogenic, and osteogenic lineage. Care was taken that the cotMSCs and bmMSCs used were in the same passage (P3). cotMSCs cultured for the same time period but in standard culture medium (described above) served as control.
Chondrogenic differentiation
For chondrogenic differentiation, 350,000 cells were suspended in 3 mL chondrogenic differentiation medium (StemPro Chondrogenesis Differentiation Kit; Gibco) and centrifuged for 5 min at 280 g, 4°C to form a pellet. The cell pellets were cultured in duplicate in 15-mL Falcon tubes and were daily shaken carefully to prevent attachment to the plastic tube. Medium was changed twice weekly. After 3 weeks of cultivation in differentiation medium, the pellets were fixed with buffered 4% formalin (ACM, Herba Chemosan Apotheker AG) and subjected to paraffin embedding and sectioning. The sections were stained with Alcian blue (Chroma, Waldeck GmbH) according to a standard staining protocol 89 to confirm the production of acidic mucosubstances.
Osteogenic differentiation
Osteogenic differentiation was performed in monolayer using the osteogenic differentiation medium (StemPro Osteogenesis Differentiation Kit; Gibco). First, 1000 cells/cm2 were seeded on 24-well plates in standard culture medium. After 3 days, the medium was replaced by osteogenic differentiation medium. Medium was changed twice weekly. After 3 weeks, the cells formed three-dimensional structures (cell sheets or nodules), which were fixed with buffered 4% formalin, harvested, paraffin embedded, and sectioned. To demonstrate extracellular calcium deposition, as sign for osteogenic differentiation, von Kossa staining (silver nitrate, Merck; Natriumthiosulfat, Sigma Aldrich) was performed. 89
Adipogenic differentiation
Adipogenic differentiation was performed in monolayer culture. After cell seeding (1000 cells/cm2) in standard culture medium and culturing for 3 days, adipogenic differentiation was induced using adipogenic differentiation medium (StemPro Adipogenesis Differentiation Kit; Gibco). Medium was changed twice a week. After 3 weeks, the cells were fixed with buffered 4% formalin and stained for intracellular lipid vacuoles with Oil red O (Sigma-Aldrich) according to a standard staining protocol. 89
Flow cytometry analysis
Cell surface antigen expression of cotMSCs (n = 5) and bmMSCs (n = 3) was analyzed using positive and negative surface markers to define their stem cell character (Table 1). The markers have been selected according to Boos et al. 91 For flow cytometry, cells at P2 were detached from the flask using trypsin. MSCs were washed with FACS buffer containing PBS, 3% FCS, and 0.1% NaN3 (Sigma Aldrich) and pelleted by centrifugation for 5 min at 400 g. Subsequently, 1 × 105 cells per sample were incubated with a specific primary antibody at 4°C. After 30 min, unbound antibody was removed by washing with 1 mL FACS buffer. The cell pellets were resuspended in 200 μL FACS buffer and analyzed with a BD FACSCanto II (BD Biosciences). Per sample, 1 × 104 events were recorded. Unstained and isotype controls were performed. The results were analyzed by FlowJo (10.0.7) software.
Results
Collection of cotyledons
Collection of the afterbirth was easy and not associated with any harm to the ewe or the newborn lamb. After mincing and digesting the cotyledons, a viscous bloody solution was obtained. Despite the tissue harvest in a farm environment, no bacterial or mycotic contamination of the cells was noticed in culture.
MSC isolation and culture
MSCs could be reliably isolated from all cotyledon (n = 5) and bone marrow (n = 3) samples with a reproducible cell yield (Table 2). MSCs from both cell sources fulfilled the minimal criteria of MSCs defined by the International Society for Cellular Therapy (spindle-shaped morphology, adherence to plastic, and colony formation). 92 The morphology of cotMSCs at different stages of colony confluence in P0 is presented in Figure 2. It was similar to the widely and well-known morphology of the “gold standard” bmMSCs.

CotMSCs in P0 showing spindle-shaped morphology, adherence to plastic, and colony formation at different stages of colony confluence. Scale bar 100 μm. cotMSCs, cotyledon-derived mesenchymal stem cells.
bmMSCs, bone marrow-derived MSCs; cotMSCs, cotyledon-derived MSCs.
Cell proliferation
Cell proliferation was evaluated for cotMSCs and bmMSCs (Fig. 3). The mean population doubling rate per day of bmMSCs was 1.672. The individual values were 0.920, 1.789, and 2.308. The mean cell doubling rate of the cotMSCs was 0.729. The individual values were 0.294, 0.635, 0.877, 0.879, and 0.960) (Fig. 3A, B). Accordingly, the generation times for bmMSCs (mean: 0.693 days, individual values: 0.433, 0.559, and 1.086) were shorter than for cotMSCs (mean: 1.659 days, individual values: 1.041, 1.137, 1.140, 1.575, and 3.405). Due to the low sample number, the proliferation differences between cotMSCs and bmMSCs did not reach statistical significance (population doubling per day: p = 0.135; generation time: p = 0.101).

Cell proliferation. Population doubling rates per day of bmMSCs and cotMSCs
Migration assay
After seeding cotMSCs and bmMSCs for the migration assay, they reached confluence after 3 days and a scratch was performed (Fig. 4A, B). The scratch was photographed every 6 h to show cell migration into the cell-free area. Although bmMSCs showed subjectively faster migration at all time points (Fig. 4C–F), after 48 h, both cell types had closed the gap confirming their migration potential (Fig. 4G, H).

Migration assay. To assess migration capacity of cotMSCs
MSC differentiation
Both cotMSCs (Fig. 5, line 1–5) and adult bmMSCs (Fig. 5, line 6) showed chondrogenic differentiation potential with a subjectively higher chondrogenic potential inherent to cotMSCs (Fig. 5, row 1, line 1–5) compared to the bmMSCs (Fig. 5, row 1, line 6). After 3 weeks in culture, the cotMSCs showed subjectively more production of extracellular acidic polysaccharides and mucopolysaccharides than bmMSCs. Also, lacunae formation around the cells, which is indicative for cartilage tissue, appeared more distinct for the cotMSCs than the bmMSCs. The control samples showed no indication of chondrogenic differentiation.
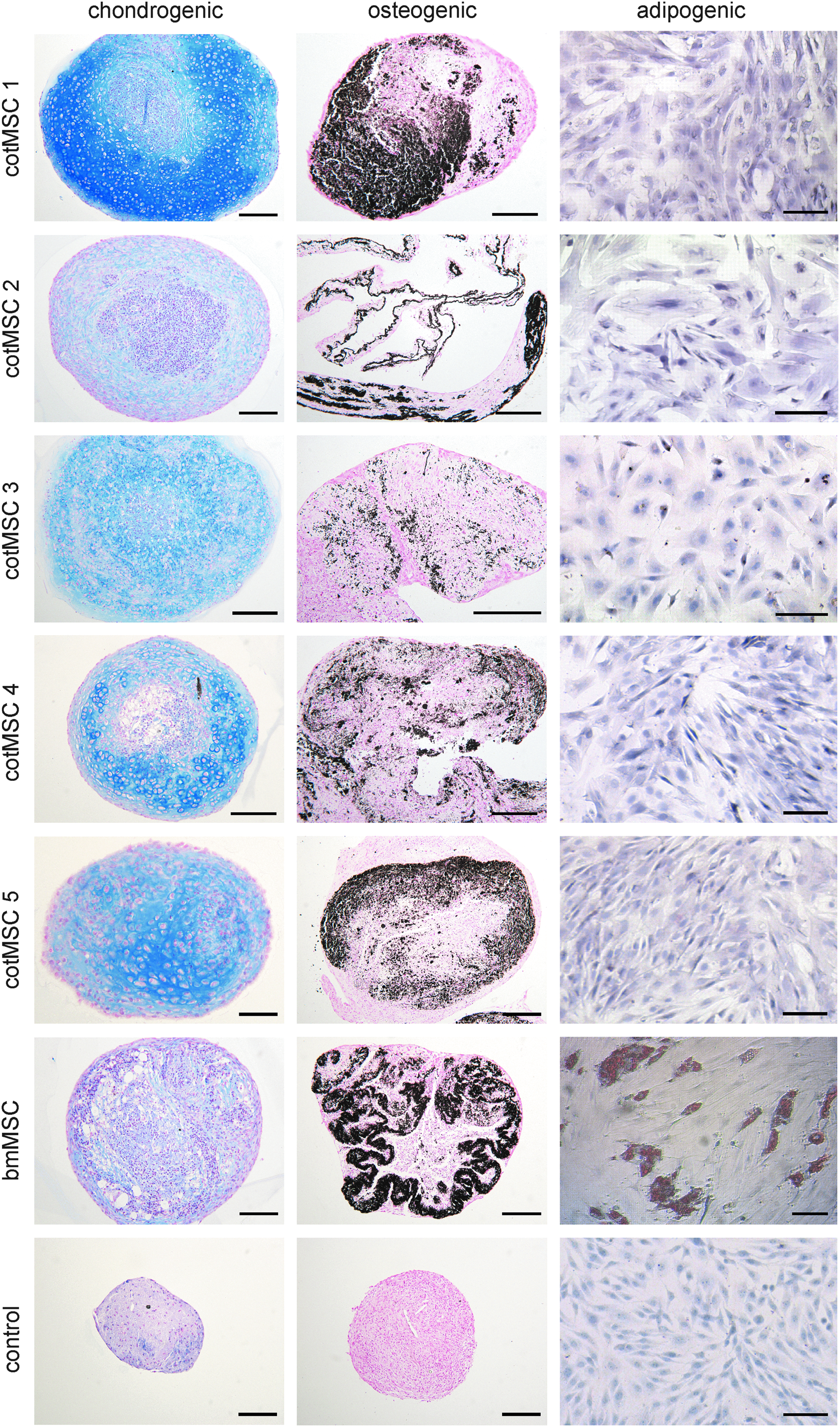
MSC differentiation. Chondrogenic (row 1), osteogenic (row 2), and adipogenic (row 3) differentiation of cotMSC (line 1–5) and bone marrow-derived MSC (line 6). Acidic mucosubstances produced during chondrogenic differentiation were demonstrated as extracellular blue substrate with Alcian blue staining. After osteogenic differentiation, von Kossa staining displayed black extracellular calcium deposition. Oil red O-positive droplets inside the cells confirmed adipogenic differentiation of bone marrow-derived MSCs (row 3, line 6), whereas cotMSCs showed no positive staining (row 3, line 1–5) indicating a lack of adipogenic differentiation potential. cotMSCs cultured for the same time period in standard culture medium served as control (line7). Scale bar = 100 μm (row 1 and 2) and 50 μm (row 3), respectively.
Osteogenic differentiation was demonstrated for both cell sources. Extracellular calcium deposition was confirmed by van Kossa staining (Fig. 5, row 2). After 3 weeks in adipogenic differentiation medium, cotMSCs showed no positive staining (Fig. 5, row 3, line 1–5) indicating a lack of adipogenic differentiation potential. In contrast, bmMSCs showed obvious and numerous intracellular lipid vacuoles that stained intensively for Oil red O (Fig. 5, row 3, line 6). cotMSCs cultured for the same time period in standard culture medium served as control (Fig. 5, line7).
Flow cytometry analysis
FACS analysis confirmed the expression of MSC markers on the protein level. In accordance to Boos et al., 91 cotMSCs and bmMSCs were positive for the surface markers CD29, CD44, and CD166 and negative for CD31 and CD45, which are typical hematopoietic cell markers. Isotype controls (IgG1 and IgG2a) were negative. FACS histograms are depicted in Figure 6.
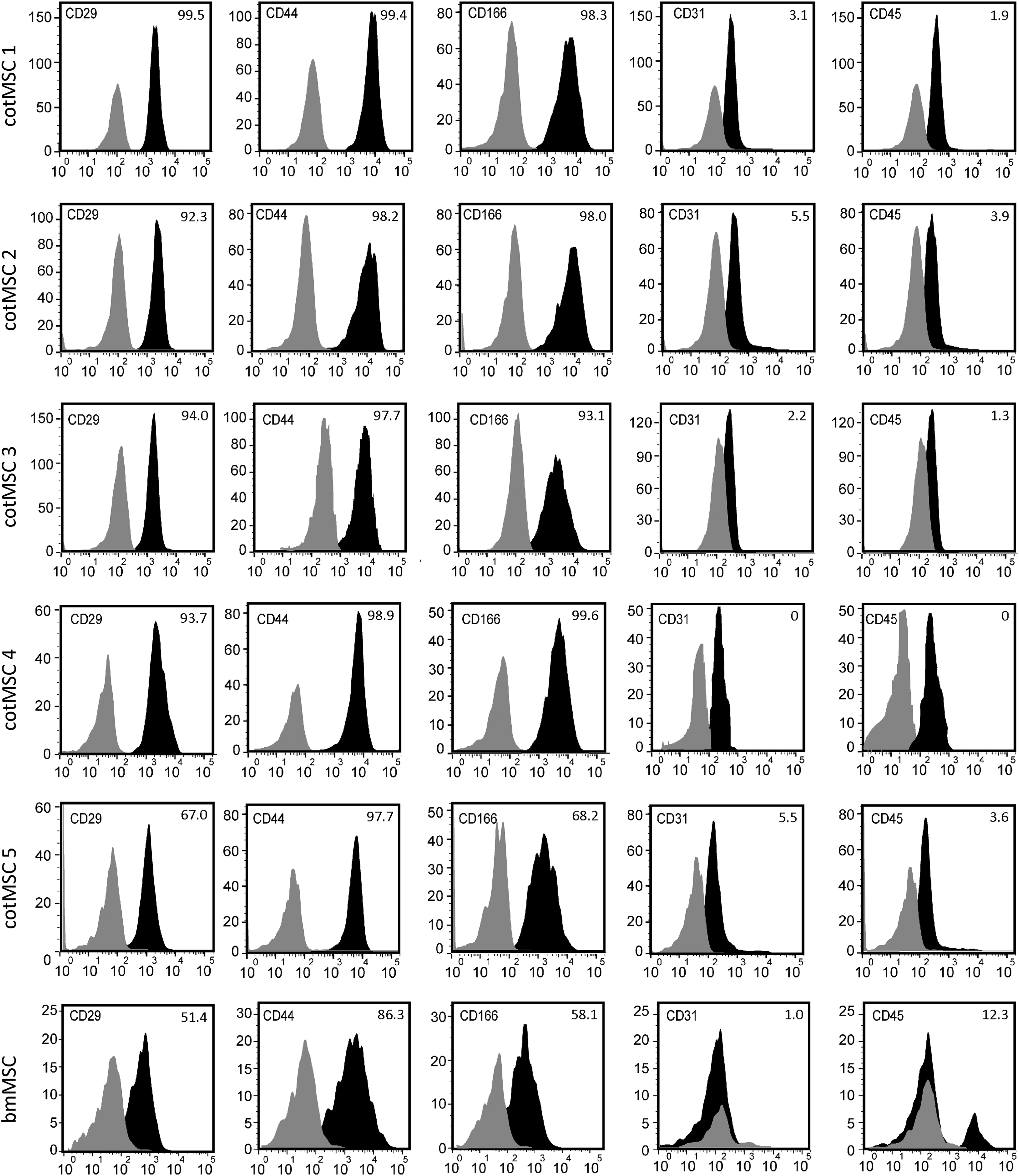
FACS analysis of cotMSCs and bmMSCs. Both MSC types were positive for the surface markers CD29, CD44, and CD166 and negative for the hematopoietic markers CD31 and CD45. Black = histograms primary antibodies, gray = histograms controls. Percentages of live, positively stained cells in relation to controls are given in the right upper corner of each histogram.
Discussion
Sheep are a frequently used animal model in regenerative medicine. However, truly noninvasive sources of MSCs in sheep are scarce. Fetal/perinatal tissues are typical noninvasive sources of MSCs in many species.13,15,18,19,36–69 However, in sheep, most studies using perinatal tissues such as cord blood, amniotic membranes, or amniotic fluid obtained from these tissues preterm using surgical harvesting procedures.13,18,19,36,41,43,59 In the current study, ovine placental cotyledons obtained on full-term delivery were tested for their suitability as noninvasive MSC source.
Semisterile harvesting of the ovine afterbirth was easy to perform. Although perinatal tissues bear a high risk of bacterial or mycotic contamination44,61 due to the nonsterile birth canal and the delivery in a farm environment, which make the use of antibiotic in culture mandatory, 61 no problems with either bacterial or mycotic contamination were encountered. In this study, 1% iodine solution was used as disinfectant to clean the obtained afterbirth from potential bacterial or mycotic contamination immediately following harvest. In addition, the cotyledons were kept in a PBS solution containing antibiotics until further processing. The obtained MSCs performed well in culture and during differentiation and showed no negative effects of the initial cleaning step or the antibiotics. Whether this cell source would also be feasible for harvest and culture without the addition of disinfectants and antibiotics requires further investigation.
During processing of the cotyledons, residual blood, which was contained in the blood vessels of the chorionic villi, resulted in a blood-like appearance of the solution obtained after mincing and digesting the villi. This finding supports the theory that the isolated MSCs do not solely originate from the perivascular niche in the placental chorionic villi (Fig. 1E) as reported for humans 77 but also from the residual fetal blood contained in the blood vessels of the chorionic villi (Fig. 1F). The fetal part of the ovine placenta (the cotyledon) and the maternal part disconnect without injury to the maternal tissue. Due to this nonbloody separation, the blood contained in the cotyledon is exclusively of fetal origin. Therefore, it stands to reason that the isolated MSCs derive from the perivascular niche in the chorionic villi and the residual fetal blood. For a more detailed characterization of the exact origin of cotMSCs, further studies will be necessary. With the isolation technique described in this article, fibroblasts are likely contained in the early passage cell population in addition to the desired MSCs. However, it was previously shown that fibroblast contamination did not adversely influence MSCs but even led to better differentiation potential than shown for fibroblast-depleted MSC populations. 93
In the current study, we demonstrated that cotMSCs have good osteogenic and chondrogenic but very limited to no adipogenic differentiation potential. It is well known that MSCs have different degrees of phenotypic plasticity depending on the tissue source.37,72,94,95 Despite similar morphology and immunophenotypic signature during expansion and growth, the differentiation potential toward the adipogenic lineage can be greatly influenced by the tissue of origin and the developmental stage (adult or fetal). 96 Adipogenic differentiation potential relies on the activation of the peroxisome proliferator–activated receptor γ (PPAR-γ) pathway, which is reduced or delayed in fetal MSCs, resulting in strong impairment of adipogenesis. 96 Since the cotMSCs used in the current study were obtained from perinatal tissue of fetal origin, the reduced PPAR-γ pathway may be a reason for the lack of adipogenic differentiation observed in these MSCs. This finding is also in accordance to other studies, which reported a low adipogenic differentiation potential for equine cord blood-derived MSCs.63,94,95,97,98 Age-related differences in adipogenic differentiation potential were also reported for muscle stem cells. It was shown that muscle stem cells have a greater propensity to adipogenic differentiation with advanced age. 99
Despite the reduced adipogenic differentiation, the cotMSCs met all minimum criteria required for the characterization of MSCs. 92 They were spindle shaped, showed formation of colonies in culture, adherence to plastic, and expression or lack of expression of specific surface antigens detected by FACS (positive for CD29, CD33, and CD166, negative for CD31 and CD45). The MSC yield was reliable as MSCs were obtained from all five afterbirths included into the study. CotMSCs and bmMSCs showed good migration potential confirmed by scratch assays. Although cell proliferation and migration of cotMSCs seemed overall slower than for adult bmMSCs, which is in contrast to findings of other authors who reported a higher proliferation rate of juvenile compared to adult MSCs.70,71
However, after further fine-tuning of the harvesting, isolation, and culture conditions, some of the shortcomings of the cotMSCs in comparison to bmMSCs may be overcome. For instance, the cotMSC yield may be raised considerably as the average ovine afterbirth comprises about 100 cotyledons87,88 and in the current study only 7–11 per donor were processed. Due to the noninvasive harvesting modalities, sheep cotyledons may lend themselves as a valuable source of MSCs to consider in the light of the principles of the 3Rs (reduce, refine, replace) for studies performed on these frequently used model animals.
In summary, in this study, MSCs have been successfully harvested from ovine placenta cotyledons. They exhibit morphological, immunophenotypical, and multipotential characteristics similar to those observed in bone marrow-derived ovine or human MSCs.3,92 Ovine cotyledons were shown to be a reliable, abundant source for the truly noninvasive, pain- and risk-free harvest of MSCs. They can be harvested easily and without a bigger risk of bacterial or mycotic contamination than any other perinatal tissue obtained on full-term delivery. The collection procedure does not interfere with partum or the initial bonding phase between ewe and lamb and is therefore exempt from ethical debate. The isolated MSCs can be cryopreserved for later use in in vitro studies or for autologous as well as allogeneic in vivo applications. Further characterization of sheep cotMSCs may be beneficial for translational studies using MSCs for particular therapeutic applications. For example, gene expression analysis of the master regulators of differentiation (Sox9, Runx2, PPARγ) and of differentiation markers during chondro-, osteo-, and adipogenesis would be interesting, to investigate if a different basal expression of the transcription factors can explain the differences in the differentiation potential.
Footnotes
Acknowledgments
We acknowledge Manuela Miksic, Brigitte Machac, Eva Haltmayer, Anna Bauer, Astrid Kohl, Michaela Pinisch, Stephanie Schmidt, Öczan Korkmaz, and the Management and Staff of the Vetmeduni Vienna Teaching and Research farm Kremesberg for their technical support.
Disclosure Statement
No competing financial interests exist.