
Abstract
Calvarial defects pose a continued clinical dilemma for reconstruction. Advancements within the fields of stem cell biology and tissue engineering have enabled researchers to develop reconstructive strategies using animal models. We review the utility of various animal models and focus on the mouse, which has aided investigators in understanding cranial development and calvarial bone healing. The murine model has also been used to study regenerative approaches to critical-sized calvarial defects, and we discuss the application of stem cells such as bone marrow-derived mesenchymal stromal cells, adipose-derived stromal cells, muscle-derived stem cells, and pluripotent stem cells to address deficient bone in this animal. Finally, we highlight strategies to manipulate stem cells using various growth factors and inhibitors and ultimately how these factors may prove crucial in future advancements within calvarial reconstruction using native skeletal stem cells.
Introduction
M
Clinical strategies for reconstruction include autologous tissue from the patient's own body (autografts), allogeneic tissue such as cadaveric bone (allografts), xenogeneic tissue from animals (xenografts), or synthetic (alloplastic) materials. Autologous sources can either be vascularized, such as free vascularized fibular grafts, or nonvascularized, such as calvarial outer table bone grafts. 10 However, autologous tissue for bony reconstruction has limitations due to donor site morbidity, bone resorption, and lack of tissue availability.11–13 Synthetic alloplastic materials can be divided into inorganic and organic, with synthetic substances ranging from titanium to hydroxyapatite and organic substances such as collagen. Although synthetic sources may possess superior mechanical properties, they are limited in their ability to integrate and grow with the host bone.14,15
Combining cellular therapies, our understanding of cytokines controlling bony healing, and advances in materials science, researchers have begun to develop regenerative strategies to address deficient bone.16–19 The use of animal models to investigate reconstruction of calvarial defects provides scientists, engineers, and clinicians with an in vivo system to investigate and augment bony healing. The calvarium is easily utilized due to minimal load bearing, as it is relatively free from exogenous forces in comparison to long bones. Outcomes of various reconstructive interventions can be assessed radiologically, with the use of microcomputed tomography, histologically, and at a cellular level with high-resolution imaging modalities. 20 In this review, we present some of the animal models used to study calvarial defects, in particular the mouse, which have improved our understanding of the key regulatory pathways that govern calvarial development, healing, and reconstruction. Investigations with these animals have facilitated creation of novel cell-based approaches to address challenging calvarial defects.
Animal Models of Calvarial Defects
Several large and small animal models have been developed over the last half century to study calvarial bone healing (Table 1). 21 In particular, the rhesus macaque and beagle have been frequently used to evaluate bone regeneration, and early development of growth factor-based regenerative strategies has emerged from these models.22,23 Similarly, the positive effects of bone morphogenetic proteins (BMPs) were described in healing calvarial defects of sheep. 24 Finally, pigs have been used to assess the synergistic effects of scaffolds and growth factors in healing calvarial defects. While these large animal models offer a potentially closer clinical model to those of humans in bony anatomy, morphology (lamellar bone structure), bone mineral density, and bone healing timeframes, they have the obvious disadvantages of requiring larger, often expensive housing, take longer to reach skeletal maturity, and are difficult to handle and costly to operate. 25
Smaller animals such as the rabbit have also been used to investigate the healing capabilities of various scaffolds, growth factors, and stem cells.26,27 The rabbit, although small, has a similar bone mineral density to humans, making studies on this animal clinically relevant, and they are an effective model for assessment of healing over scarred dura mater. 28 To mimic clinical factors affecting healing, rabbits have also been used to investigate regeneration of calvarial defects after infection with Staphylococcus aureus and after irradiation.29,30 However, defect sizes used in rabbits have been ill-defined, with reports utilizing parietal bone defects ranging from 8 to 17 mm in diameter.31–33 In addition, rabbits are known to have a superior ability to regenerate bone in comparison to other animal models such as primates and rodents, which may render in vivo studies with this animal overly optimistic in their translational potential to humans.34,35
In contrast to rabbits, murine models have been shown to have a less robust tissue regenerative ability, with bone-forming capacity more similar to humans. 36 Murine models have thus been used to study calvarial regeneration, particularly as age-related differences in calvarial regeneration have been appreciated, which parallel clinically observed differences in bone-forming capacity between infants and older patients.2,37 While both rats and mice have proved to be effective models (Table 2), studying calvarial defects in mice has allowed for use of transgenic animals, which has opened the door to determining tissue origins of the calvarium, signaling pathways involved in development and healing, and strategies to enhance bone formation in the calvarium. With these capabilities, the mouse model thus remains at the forefront for development of techniques to reconstruct calvarial defects.38,39
Development of the Calvarium
Embryologic development of the calvarial bones provides a foundation for understanding subsequent healing of the postnatal calvarium and design of tissue engineering approaches to bone injury. Early seminal work by Le Douarin using the quail-chick chimera system to decipher origins of the avian cranial bones revealed that frontal bones were fully neural crest-derived while the remainder of the calvarial vault appeared to be of mixed mesodermal and ectomesenchymal origin.40,41 Other nonmammalian models, including Xenopus laevis and zebrafish, have subsequently reiterated Le Douarin's findings and have shown that there is a high degree of developmental conservation between species.38,42 More recent delineation of embryonic origins for individual elements of the calvarium have been facilitated by studies in transgenic mice with use of animals doubly heterozygous for Wnt1-Cre and R26R alleles.38,43 Combining indelible β-galactosidase labeling of neural crest cells with DiI labeling of paraxial mesoderm, Jiang et al. have provided a more refined description of tissue derivation for the skull, with dura mater and frontal bones originating from cranial neural crest and parietal bones deriving from mesoderm. 38 Interestingly, many of the natural bone-forming growth centers in the skull, known as cranial sutures, were found to form at different interfaces between mesodermal and neural crest-derived tissues. Finally, the interparietal bone was described to arise as a composite of both neural crest and paraxial mesoderm origin.
Calvarial Healing in the Murine Model
Underscoring the importance of understanding embryologic derivation of the calvarial bones, investigations have shown frontal bone defects in mice to regenerate faster than those in the parietal bone. Interestingly, neural crest-derived bones of the calvarium have been found to express increased levels of fibroblast growth factor (FGF)-2, -9, and -18, as well as their cognate receptors (FGFR-1, -2, and -3), and increased FGF expression in the frontal bone has been described to enhance osteogenic potential of resident osteoblasts.44–46 Prior studies by Iseki et al. have demonstrated that active sites of osteogenesis in the developing skull are governed by FGFR-2 signaling, and increased FGF-2/FGFR-2 pathway activity, whether endogenous or ectopically induced, resulted in increased proliferation of calvarial bone-derived osteoblasts.47–49 Moreover, FGF-2 has been reported to induce glycogen synthase kinase-3 beta phosphorylation, with subsequent enhanced Wnt signaling. 44 These findings suggest that Wnt pathway activity in neural crest-derived tissue may be regulated by FGF-2 signaling, mediating enhanced frontal bone regenerative capacity. Based on these results, subsequent studies on Axin knockout mice have reported increased proliferation and reduced apoptosis of calvarial osteoblasts, and concomitant increased Wnt signaling among mesoderm-derived osteoblasts enhanced their bone regenerative capacity. 39 Importantly, recent work by Skogh et al. has now shown that the healing capability of frontal bone is greater than that of parietal bone in humans. 50 These findings thus confirm that developmental pathways are also keys to differential bone regeneration in humans.
Apart from specific tissue origin of individual bones, the ability for young animals as well as humans to regenerate calvarial defects has also been well documented, and studies in mice have begun to elucidate mechanisms underlying this capacity. 51 Investigations evaluating gene expression within the early regeneration of calvarial defects have revealed upregulation of multiple osteogenic growth factors, including BMP-2, -4, and -7, as well as FGF-2 and its receptor FGFR-1, in skeletally immature mice. 52 Interestingly, Cowan et al. have also observed juvenile murine calvarial-derived osteoblasts to remain in a more dedifferentiated state compared with adult osteoblasts, and FGF-2 stimulation of these young osteoblasts resulted in increased expression of collagen 1a and osteocalcin, along with enhanced formation of bone nodules. 19 Building on these findings, administration of BMP-2 and FGF-2 has been reported to address age-associated reduction in calvarial bone healing, with delivery of these growth factors in low dosages to parietal defects of old mice, resulting in improved reossification and radiographically measured bone volume. 53
Age-related reduction in bone formation has also been attributed to alterations in Wnt signaling, and investigations in mice have demonstrated reduced Wnt expression and canonical Wnt signaling within cortical long bones and vertebral bodies with increasing age.54,55 Similarly, decreased Wnt signaling has also been identified within the adult mouse parietal bone, with expression analysis demonstrating increased levels of sclerostin, a Wnt pathway inhibitor. 56 Exogenously induced Wnt signaling in bone grafts from skeletally mature mice through brief ex vivo incubation with liposomal Wnt3a was found to significantly improve calvarial defect reconstruction. Reduced Wnt signaling has also been associated with age-related fatty changes seen in osteoporosis, 57 and these findings suggest a potential regenerative-based strategy for revitalizing calvarial bone healing potential in aged patients.
Finally, unique to critical sized calvarial defects is the complex interplay of surrounding tissues, namely the underlying dura mater and the overlying periosteum. While periosteum is well known to promote regeneration in long bones, in the calvarium, dura mater has also been shown to participate in bone healing, contributing both osteoprogenitors and key pro-osteogenic growth factors to promote calvarial regeneration.58–60 Interruption of the dura mater interface with overlying bone defect has been observed to impair calvarial healing as well as suture fusion. 61 Greenwald et al. demonstrated that immature murine dura mater expresses osteogenic factors such as transforming growth factor (TGF)-β as well as produces extracellular matrix components such as collagen 3 and alkaline phosphatase. 62 Transcriptional analyses between mouse juvenile and adult dura mater have also demonstrated upregulation of growth factors such as BMP-2 and FGF-2, along with several osteoblast differentiation markers (osteopontin, runt-related transcription factor 2, and FGFR-1).52,63,64 Interestingly, juvenile dura mater has also been found to express increased markers for osteoclast activity, including acp5, matrix metalloprotease-9, and cathepsin K. 52 Localization of osteoclasts within the dura mater of young mice has been shown histologically, and these findings suggest that bone homeostasis within the young calvarium may be dependent not only on osteoblastic activity but also on coordinated osteoclastic bone resorption. Collectively, these findings thus highlight the importance of the underlying dura mater in calvarial bone regeneration, and alteration or damage to this tissue may impair any reconstructive attempt.
Cell-Based Strategies for Calvarial Reconstruction
Development of novel clinical strategies to address calvarial defects has benefited from the establishment of critical-sized defect animal models, particularly in the mouse (Table 2). Creation of critical-sized calvarial bone defects, those that will not heal spontaneously during the life of the animal, 65 has been well described, with Aalami et al. demonstrating that 3, 4, and 5 mm size full-thickness defects within the parietal bone fail heal in adult wild-type mice. 2 However, similar-sized defects in juvenile animals were noted to heal, which falls in line with clinical observations of enhanced infant calvarial regenerative capacity. Furthermore, a critical-sized calvarial defect model has also been established in immunocompromised, athymic mice, allowing for in vivo study of implanted human osteoprogenitors cells. 66 These descriptions have provided a valuable platform for study of cell-based approaches to mouse calvarial reconstruction with translational promise.
The ability to heal bone defects in elderly patients or in those with medical comorbidities, including osteoporosis, diabetes, and cancer, highlights the fact that acellular approaches alone are usually insufficient. While improvements continue in development of scaffolding materials, a full discussion of which is beyond the scope of this review, effective regenerative approaches will likely require cellular building blocks and incorporation of growth factors (Fig. 1). Contributing to the difficulty in translating this strategy, however, includes the lack of consensus over source of osteogenic cells. Various cell types have been explored for healing mouse critical-sized calvarial defects, and most commonly these have included both adult stem cells such as bone marrow-derived mesenchymal cells (BM-MSCs), adipose-derived stromal cells (ASCs), and muscle-derived stem cells (MDSCs), as well as nonadult stem cells such as induced pluripotent (iPSCs) and embryonic stem cells (ESCs). Finally, the recent discovery of a resident mouse skeletal stem cell (mSSC) opens the door for future development of strategies to isolate and/or reactivate these progenitor cells to regenerate large defects in calvarial bone.
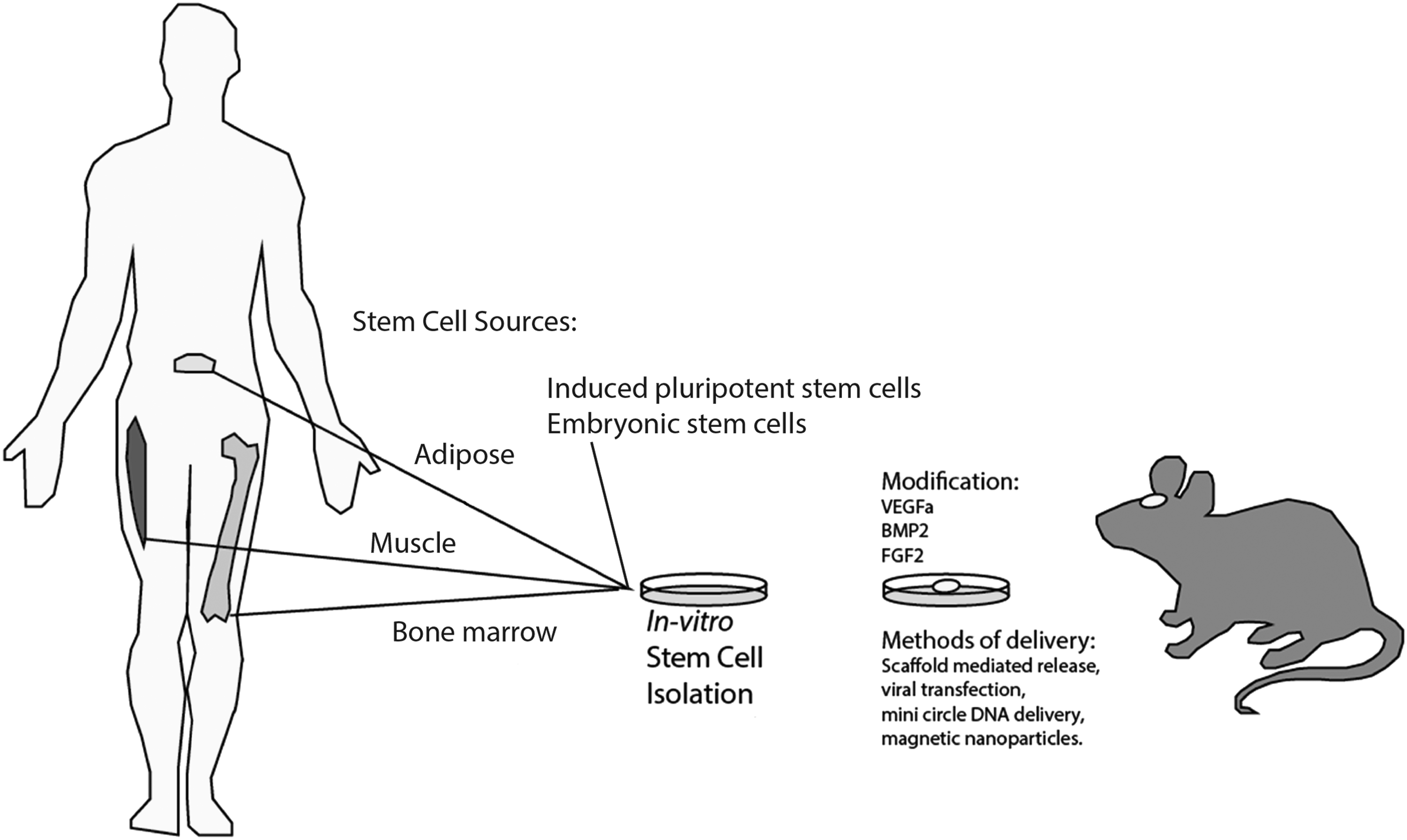
Schematic of stem cell sources and modifications to enhance calvarial healing. Stem cell sources include bone marrow-derived mesenchymal cells, adipose-derived stromal cells, and muscle-derived stem cells, as well as nonadult stem cells such as induced pluripotent (iPSCs) and embryonic stem cells (ESCs). Exogenous factors, including VEGF, BMP-2, and FGF-2 have also been studied to enhance calvarial bone regeneration. BMP, bone morphogenetic protein; FGF, fibroblast growth factor; VEGF, vascular endothelial growth factor.
The bone marrow has long been known as a source of mesenchymal stem cells capable of differentiating into bone, cartilage, stroma, and fat.67,68 Autologous BM-MSCs are often cited as the ideal source of stem cells for research on bone healing, 69 and apart from their osteogenic capacity, BM-MSCs can perform pericyte-like functions by secreting angiogenic and stabilization factors to enhance vessel formation. 70 Mouse BM-MSCs have been shown to heal critical-sized defects in syngeneic animals,18,71 and transgenic mice, including Col3.6Topaz fluorescent osteoblast reporter mice, have demonstrated implantation of BM-MSCs to directly contribute to bone formation in critical-sized calvarial defects. 72 The mouse critical-sized calvarial defect has also been used to investigate the regenerative capacity of human-derived BM-MSCs, and several reports have documented the ability for these cells to similarly promote in vivo bone regeneration.73–75 However, while human BM-MSCs have been documented to enhance mouse calvarial regeneration, some reports have observed poor survival of implanted cells and a preponderance of host-derived tissue within the central defect. 75 This may be due to limitations in life span of transplanted BM-MSCs, as enhanced expression of human telomerase reverse transcriptase has been reported to improved bone repair. 76 Perhaps an even greater drawback of BM-MSCs is the invasive nature of the isolation procedure and the low yield of these cells within the bone marrow aspirate, which may require ex vivo expansion for reconstruction of larger, more clinically relevant defects.77,78
Over the past decade, a more readily available source of mesenchymal cells found in adipose tissue has been thoroughly evaluated for calvarial bone regeneration, with Cowan et al. first describing the ability for mouse-derived ASCs to heal critical-sized defects in the skull. 18 Levi et al. later demonstrated the same capacity for human ASCs to regenerate calvarial bone. 79 Since then, the mouse calvarial defect model has helped to reveal several critical underlying mechanisms guiding ASC-mediated in vivo bone formation. Interestingly, while human ASCs can be utilized to address acute mouse calvarial bone deficiencies, healing of more chronic defects was substantially impaired due to reduced vascularity and endogenous BMP production from surrounding tissue. 80 Underscoring this point, the dura mater has been shown guide implanted human ASC osteogenesis through BMP-2 signaling, and injury to the dura mater at the defect base dramatically reduces calvarial healing. 81 These findings thus raise important considerations for the clinical use of ASCs in reconstructing long-standing calvarial defects.
In addition, while the osteogenic capacity of ASCs has been well documented, not all ASCs exhibit similar capabilities. Depot-specific differences have been observed in bone formation of human ASCs, 82 and more recently, granular single-cell transcriptional studies have revealed tremendous ASC heterogeneity even within the same harvest site. 83 Investigations using linear regression analysis have identified surface marker correlates with bone-forming genes, and CD105 expression has been shown to be effective at isolating a functional subpopulation of enhanced bone-forming cells. In particular, CD105low ASCs have been found to possess increased in vitro bone-forming capacity, as well as a greater ability to form bone in mouse critical-sized calvarial defects. 83 CD105, also known as Endoglin, has been described to function as a TGF-β coreceptor, possibly explaining why this factor may impact bone formation. Subsequent studies have also identified CD90 (Thy1) as capable of defining subsets of osteogenic ASCs, and Chung et al. demonstrated the ability of CD90+ cells to augment mouse critical-sized bone regeneration compared with CD90− and unsorted ASCs. 84 However, the precise role of CD90 in determining the capacity of ASC to form bone remains unclear. These findings, in addition to the fact that both CD105 and CD90 cell-surface expression have been found to significantly drift over time, have led to other studies fractionating ASCs based on more classical cell surface markers, such as BMP receptor (BMPR)-IB, which are directly implicated in osteogenic differentiation. 85 With this approach, isolated BMPR-IB+ human ASCs were also reported to improve healing of mouse critical-sized calvarial defects. 86 Ongoing studies have begun to evaluate which of these markers may prove the most effective at bone regeneration. But despite an increasingly refined understanding of ASCs, some studies have suggested a less inherent osteogenic potential for ASCs compared with BM-MSC, particularly without exogenous growth factors such as BMP. 87 Furthermore, similar to BM-MSCs, substantial contribution of implanted human ASCs to the bone regeneration in the mouse calvarial defect has been called into question, with recent reports suggesting a more paracrine role of ASCs in the activation of resident osteogenic cells through Hedgehog signaling. 88 Emerging studies have suggested upregulation for B-cell lymphoma 2 expression in ASCs to enhance their survival and contribution to bone formation in mouse calvarial defects following implantation,89,90 but more long-term in vivo studies are necessary before ASCs can truly be evaluated in the clinical setting.
Similar to bone marrow and adipose tissue, muscle is another source for adult, multipotent mesenchymal cells, and the ability for ectopic bone formation within muscle, as seen with heterotopic ossification, has lead researchers to investigate these cells for calvarial bone regenerative strategies. 91 While relatively less studied than their BM-MSC and ASC counterparts, human MDSCs have been similarly isolated using cell adherence techniques and are likewise capable of expressing several key osteogenic markers under appropriate culture conditions. 92 In the mouse critical-sized calvarial defect, human MDSCs have also been shown to generate bone, particularly when implanted with exogenous BMP or transduced to overexpress this growth factor.93,94 Furthermore, recent work by Lough et al. comparing mouse BM-MSCs, ASCs, and MDSCs in healing calvarial defects found that MDSCs, unlike BM-MSCs and ASCs, were capable of regenerating vascularized corticocancellous bone with diploic space comparable to native bone. 95 These findings suggest MDSCs may possess a subpopulation of cells capable of hierarchical differentiation into lineage-restricted progenitors, including both osteogenic cells and hematopoietic supportive stroma, which are less frequent in bone marrow and adipose counterparts. However, some studies have suggested the in vivo bone-forming capacity for MDSCs to be dependent on BMP, 94 and both donor site morbidity and ability to obtain sufficient numbers of cells without ex vivo expansion limit the clinical utility of these cells. 96
In contrast to adult stem cells, pluripotent stem cells allow for enormous potential in reconstructing all tissue types in the craniofacial region.97–100 With respect to skeletal tissue, in vitro osteogenic differentiation of pluripotent cells has been thoroughly demonstrated, and recent studies by Ma et al. revealed similarities in global osteogenic gene expression profiles and osteogenic capacities for mouse ESCs and iPSCs. 101 Human iPSC exosomes have also been shown to enhance angiogenesis and osteogenesis in healing of murine critical-sized defects, 102 and upregulation of special AT-rich sequence-binding protein 2, a critical regulator of craniofacial bone development, further enhanced the ability of human iPSCs to regenerate mouse calvarial defects. 103 Importantly, use of pluripotent cells still carries the risk for teratoma formation, and the development of these tumors in various mouse models has led to significant animal disability and death. While several approaches have been studied to address this concern, including osteogenic priming through ex vivo culture, investigations have also demonstrated the ability to direct in vivo differentiation of pluripotent cells following implantation through targeted manipulation of the surrounding niche. 104 Incorporation of BMP-2 into the implanted cellular microenvironment facilitated engraftment of pluripotent human iPSCs and ESCs into mouse critical-sized calvarial defects and de novo bone formation with reduced risk for teratoma formation. However, while these cells represent powerful tools for cell-based tissue regeneration, safety concerns remain, and multiple barriers must still be overcome before this technology can be applied clinically.
Finally, recent discoveries in mouse skeletal biology have led an improved understanding in how skeletal tissues are derived, and the description of a mSSC defined by the cell surface marker profile, CD45−Ter119−Tie2−AlphaV+Thy−6C3−CD105−CD200+, has opened the door to strategies targeting activation of this cell. 16 Making this concept more compelling is the fact that several studies have failed to demonstrate significant persistence of implanted cells contributing to mature, regenerated calvarial bone in the mouse.88,104 Skeletogenesis occurs through a developmental hierarchy of lineage-restricted progenitors, similar to hematopoiesis, with formation of subpopulations from mSSCs, including the bone cartilage stromal progenitor (BCSP), which subsequently gives rise to more lineage committed progenitors. In the setting of acute skeletal injury, an activated mouse CD49f+ BCSP has been identified, characterized by increased hedgehog signaling, and with disease states such as diabetes, fracture repair can be enhanced through exogenous activation of the hedgehog pathway.17,105 Interestingly, prior studies showing ASC-mediated activation of resident calvarial osteogenic cells through Hedgehog signaling may thus be describing effects on the BCSP. Ongoing studies identifying the mSSC and BCSP in mouse calvarial bone, as well as their human analog, may therefore provide for a novel cellular target, which can be potentially stimulated to promote calvarial bone regeneration.
Manipulation of Stem Cells
While investigations continue on identification of cellular building blocks for bone regenerative strategies, the mouse critical-sized calvarial defect has also provided a functional platform to evaluate addition of exogenous factors and other strategies to manipulate implanted or resident cells for the purposes of enhancing osteogenesis (Fig. 1). As the microenvironmental niche can regulate progenitor cells through diffusible factors, augmentation of this function can potentially accelerate osseous healing. While the breadth of these factors precludes an in-depth discussion of each within the scope of this review, the normal physiology of calvarial development and healing point to many promising pathways can be exploited for regeneration of critical-sized defects.
The osteogenic potential of BMPs has been thoroughly investigated in various settings, and data from gene expression analysis of early mouse calvarial regenerate as well as juvenile dura mater emphasize the importance of this growth factor for promoting bone healing in the skull. BMPs are already clinically used to enhance bone formation in FDA approved settings such as spinal fusion and tibial nonunions. Furthermore, up to 85% of BMP usage in patients is off-label for bone reconstructive purposes in both the craniofacial skeleton and extremities. 106 However, side effects, including inflammation, infections, compartment syndrome, and progressive myopathy, have been observed as well as heterotopic ossification, particularly with rhBMP-2. 107
Delivery systems with noncontrolled, nontargeted delivery of supraphysiological doses have been attributed to some of these reports. Recent strategies have therefore evolved for sustained, lower dose release of BMP-2 through incorporation of this growth factor into various vehicles, including heparin-conjugated fibrin and collagen-hydroxyapatite.108,109 With this approach, increased mineralization of murine calvarial defects through stimulation of local osteogenic progenitors has been reported, and concomitant reduction of heterotopic bone formation, adjacent bone resorption, and aberrant adipose tissue induction has been observed. Controlled delivery of BMP-2 has also been combined with implantation of BM-MSCs, ASCs, and MDSCs, as well as pluripotent cells, and in each case, enhanced regeneration of mouse critical-sized defects was appreciated.93,95,104,110 In addition, with pluripotent cells, incorporation of BMP-2 into hydroxyapatite-coated scaffolds also significantly reduced untoward outcomes, including teratoma formation, as previously described. 104
Along similar lines, other reports have incorporated small molecule BMP synergistic agonists into delivery vehicles and have shown improved calvarial bone formation. Phenamil, a derivative of the diuretic amiloride, induces the expression and stabilization of BMP signaling intermediate SMADs, and incorporation of this small molecule with ASCs improves osteogenic differentiation and critical-sized mouse calvarial defect regeneration. 111 Small molecule inhibition of TGF-β via SB431542 has also been shown to activate BMP signaling in calvarial osteoblasts and enhance bone formation in the mouse calvarium. 112 These findings, along with those from studies on sustained, controlled release delivery vehicles, support strategies aimed at reducing overall BMP dose without compromising calvarial healing efficacy.
As an alternative to direct incorporation of BMP-2 into delivery scaffolds, manipulation of implanted cells using viral and nonviral methods to enhance BMP-2 production has also been shown to accelerate healing in the mouse calvarium. Adenoviral, lentiviral, and baculoviral transduction of BM-MSCs, ASCs, and MDSCs have all been reported to be effective at inducing BMP-2 transgene expression and implanted cell-mediated calvarial bone regeneration.113–116 Furthermore, shRNA-mediated knockdown of noggin, a BMP antagonist, in lentivirally transduced ASCs enhanced endogenous BMP signaling and increased osteogenic capabilities with the mouse calvarial defect. 117 Other nonviral transfection strategies using minicircle DNA have also been reported and can potentially reduce risks of viral-mediated insertional mutagenesis. 118 However, reduced transfection efficiency and intermittent transgene expression have been observed. Addressing this limitation, a novel approach for longer release of BMP2 was recently reported through integration of minicircle DNA into polylactic-co-glycolic acid scaffolds. This approach resulted in sustained release of BMP-2 over 60 days by transfected osteoblasts and improved healing of mouse critical-sized calvarial defects. 119 The sustained release of this growth factor without a burst phase makes this approach particularly attractive for strategies aimed at guiding bone differentiation of implanted progenitor cells for regenerative purposes.
Countless studies on the mouse calvarium have also highlighted the importance of FGF-2 in regulating Wnt signaling, enhancing the osteogenic capacity of frontal bone, and contributing to the regenerative potential of juvenile mice. Several investigations have therefore incorporated exogenous FGF-2 into strategies addressing critical-sized mouse calvarial defects.120–122 In vitro and in vivo studies have shown that treatment of osseous defects with FGF-2 alone increases proliferation of osteoblasts and enhances mineralization of bone in the craniofacial skeleton. 123 FGF-2 has also been used in combination with BMP-2 to improve osseous healing in the calvarium of old mice, allowing for a reduction in BMP-2 to be used.53,124 These findings reflect the earlier work of Rice et al. who demonstrated integration of FGF and BMP signaling pathways in calvarial bone development. 49
Finally, similar to recent studies with BMP-2, novel approaches for controlled delivery of FGF-2 have also been created to enhance murine calvarial healing. Kwan et al. described the development of a functional fusion protein, with FGF-2 conjugated to a protein-destabilizing domain (DD-FGF-2), which can be regulated by the synthetic ligand Shield-1. 125 Implantation of genetically modified MC3T3-E1 cells expressing this fusion protein into critical-sized mouse calvarial defects allowed for chemical regulation of FGF-2 function, via intraperitoneal Shield-1 administration, to promote bone regeneration. This study thus provides a novel approach to temporally control effects of growth factors for targeted cellular therapies.
While multiple factors have been shown to promote bone regeneration in the mouse calvarium, treatment of defects can still remain challenging due to poor vascularity, as previously noted in acute and chronic mouse calvarial defect studies.80,126 Importantly, investigations in vascular endothelial growth factor (VEGF) deficient mice have established a significant role for this growth factor in intramembranous bone formation. 126 Murine calvarial osteoblasts have also been shown to release VEGFA, which in turn leads to increased endothelial cell migration and enhanced osteoblast differentiation. 127 With the ability to both promote angiogenesis and osteogenesis, studies have applied VEGFA to human derived ASCs and have found increased osteogenic potential in vitro, as well as improved in vivo ability to heal calvarial defects in mice. 128 Comparative studies have also suggested VEGFA to have similar capacity to BMP-2 in promoting bone formation in the mouse calvarium, and each of these factors alone proved more capable of stimulating osteogenesis than FGF-2. 120
Furthermore, a synergistic effect has been reported between VEGF and BMP signaling in promoting regeneration in the mouse calvarial defect. Investigations have suggested that VEGF may play an important role in enhancing bone regeneration by MDSCs expressing BMP, and soluble Flt1 has been shown to inhibit bone formation. 129 Interestingly, the fate of the recently identified skeletal stem cells (SSCs) has been reported to rely on the interplay of VEGF and BMP2, showing both factors working synergistically toward de novo osteogenesis. 16 However, the combinatorial effect of these two growth factors may be ratio dependent, as higher amounts of VEGF have been noted to have detrimental effects on BMP-induced calvarial bone healing. 129 Further studies refining the precise interplay between VEGF and BMP are therefore necessary before effective translational strategies applying VEGF for bone regenerative purposes can be developed.
Conclusion and Future Perspectives
We have briefly outlined the advancements made in our understanding of cranial development and calvarial bone healing, and with the mouse critical-sized calvarial defect model, various promising stem cell sources with or without exogenous growth factors have been investigated in the development of potential strategies for reconstructing calvarial defects. The murine model has proved invaluable and has led to several new promising therapeutic options, one of which, the use of ASCs for calvarial reconstruction, has already been described with some success in small clinical reports.130,131 However, discovery of the SSC in the mouse may perhaps be the most exciting finding, and future studies identifying the human homolog may allow for development of improved translational strategies to stimulate resident osteoprogenitors for regeneration of large calvarial defects.
Footnotes
Acknowledgments
M.T.L., MD, MBA was supported by National Institutes of Health Grants R01 DE021683, R56 DE025597, and U24 DE026914, the Oak Foundation, and the Hagey Laboratory for Pediatric Regenerative Medicine. D.C.W., MD was supported by National Institutes of Health Grant K08 DE024269, the Hagey Laboratory for Pediatric Regenerative Medicine, the California Institute for Regenerative Medicine, and Stanford University Child Health Research Institute.
Disclosure Statement
No competing financial interests exist.